1. Introduction
Artificial insemination (AI) is used widely in agriculture to optimize and spread commercially valuable genetic traits, including in goats. Effective cryopreservation of semen samples is critical for efficient AI. However, cryopreservation can reduce goat sperm quality, motility, and viability, resulting in lower fertility rates [
1]. The reduced semen quality results in part from cellular oxidative stress, which causes the peroxidation of unsaturated fatty acids in the biomembrane. In healthy mammalian sperm cells, total antioxidant capacity (TAC) and reactive oxygen species (ROS) production remain in balance [
2], but an increase in ROS production, a decrease in TAC, or both during cryopreservation can result in reduced motility and cell death [
3]. Excessive ROS production during cryopreservation and after thawing not only degrade cellular membranes but may also damage DNA [
4], further enhancing dysfunction [
5]. To reduce oxidative stress, exogenous antioxidants are frequently added to cryopreservative solutions to sustain semen quality prior to AI [
6]. Several studies have confirmed ROS scavenging capacity of various medicinal plant extracts to improve sperm motility and increase fertility rates [
7,
8,
9,
10].
The genus
Turraea L., a genus of the Meliaceae (mahogany) family of tropical flowering trees and shrubs, includes 70 species distributed throughout Africa and Asia [
11,
12].
Turraea fischeri is widely used in east Africa to treat stomachache and infertility [
13]. Previous studies have also documented antioxidant and hepatoprotective properties [
14]. Furthermore, biochemical analyses of
T. fischeri extracts have identified numerous potentially bioactive secondary metabolites such as limonoids as well as various sterols and flavonoids with antioxidant and anti-inflammatory activities [
15].
To date, there are no studies available on the effect of the leaf extract from T. fischeri in semen extenders on semen cryopreservation. In the current study, the chemical profile of a leaf methanol extract from T. fischeri was characterized using HPLC-MS/MS, and its antioxidant activity was measured by in vitro spectroscopic assays. The effects of T. fischeri leaf extract on goat sperm viability, motility, and morphology, and antioxidant capacity were then examined after thawing by vital staining under light microscopy, flow cyto-metry following Annexin/propidium iodide staining, various spectroscopic assays, and transmission electron microscopy.
2. Materials and Methods
2.1. Plant Material, Extraction, LC-MS and Antioxidant Acctivities
Leaves of
Turraea fischeri were collected from the Lupaga Site in Shinyanga, Tanzania, and stored at the Institute of Pharmacy and Molecular Biotechnology, Heidelberg University, under accession number P7336 [
16].
The dried and ground leaves (100 g) were extracted three times in 100% methanol (3 × 500 mL) at ambient temperature and the three extracts combined. Pooled extracts were then filtered and dried under a vacuum at 40 °C. The obtained residue was lyophilized, yielding a fine dried powder (15 g). LC-MS analyses and in vitro antioxidant activities were performed according to Sobeh et al. [
14]. Detailed methods are included in the
Supplementary File.
2.2. Animals
Semen samples were collected from 5 mature, fertile Baladi bucks (50–60 kg LBW; 2–4 years of age) at the Animal Production Research Station, El-Karada, Kafrelsheikh, Animal Production Research Institute (APRI), Agricultural Research Center, Ministry of Agriculture, Egypt, using an artificial vagina. Sample collection was conducted in cooperation with the Physiology and Biotechnology Laboratory, Animal Production Department, Faculty of Agriculture, Mansoura University, Egypt, according to animal welfare guidelines of Mansoura University.
2.3. Animal Management
All 5 bucks were raised under the same environmental conditions. Feeding requirements were calculated according to the recommendations of Animal Production Research Institute, Ministry of Agriculture, Egypt. Each buck was fed 1.0 kg/day concentrate feed mixture containing 14% crude protein and 70% total digestible nutrients, plus 1.25 kg/day berseem hay from August to November or 5 kg/day Egyptian fresh berseem clover (Trifolium alexandrinum) from December to February. Animals had free access to trace mineralized salt and drinking water at all times.
2.4. Collection of Semen
Semen was collected from each buck by artificial vagina once weekly before feeding at 7–8 a.m. for five consecutive weeks. In total, 25 samples were obtained. Samples were transferred immediately into a water bath at 37 °C and only those with mass motility ≥ 70% and sperm concentration of at least 2.5 × 109/mL were retained for experiments. Samples were then pooled and divided into 4 aliquots for each treatment group.
2.5. Preparation of Extender
The Tris-citric-soybean lecithin extender contained 3.025 g/dL Tris (Sigma Chemical Co., St. Louis, MO, USA), 1.66 g/dL citric acid monohydrate (Sigma, Darmstadt, Germany), 1.25 g/dL glucose (Sigma Aldrich, St. Louis, MO, USA), 5% glycerol (Honeywell, Regen, Germany), 1% soybean lecithin (L-a-phosphatidylcholine, LAB: product number MC041), 100 IU/mL penicillin, and 100 µg/mL streptomycin. The components were mixed in a water bath at 37 °C and adjusted to 300 mOsm/kg in H2O (pH 6.8) before addition of extract.
2.6. Cryopreservation
Pooled semen was diluted 1:10 in extender (v/v) containing the indicated extract concentration (0, 125, 250, and 375 µg/mL) and adjusted to a final sperm concentration of 2.5 × 108/mL. The mixture was then gradually cooled from 37 °C to 5 °C over 4 h (equilibration period) and transferred to 0.25-mL French straws (IMV Technologies, L’Aigle, France) for cryopreservation. Straws were first exposed to liquid nitrogen vapor for 10 min and then immersed in liquid nitrogen at −196 °C.
2.7. Thawing
After one month, the straws were thawed at 37 °C for 30 s in a water bath, and the various assessments conducted immediately.
2.8. Semen Evaluation
2.8.1. Progressive Motility
The proportion of sperm cells showing progressive motility was examined under a phase-contrast microscope (DM 500, Leica, Switzerland) with heated stage set to 37 °C. Briefly, aliquots of diluted sperm (10 µL) were placed on pre-warmed glass slides and sealed with coverslips. A total of 200 spermatozoa/slide from 3 randomly chosen fields were counted by the same investigator and the mean proportion (%) recorded.
2.8.2. Viability
Semen samples were double-stained with a mixture of 5% eosin (vital stain) and 10% nigrosin (background stain) to estimate the live: dead ratio [
17]. The live: dead ratio was calculated by counting the unstained head area among 300 sperm at high magnification (400×) using a light microscope.
2.8.3. Gross Structural Abnormalities
Abnormalities in gross structure were assessed in 300 sperm cells during viability measurements using a light microscope. The following criteria were considered: (i) tail defects (abnormal tails), (ii) abnormal heads, and (iii) cytoplasmic droplets [
18].
2.8.4. Plasma Membrane Integrity
The hypo-osmotic swelling test (HOS-t) was used to assess plasma membrane (PM) integrity according to a previously described protocol [
19]. Briefly, semen (50 µL) was incubated at 37 °C for 30 min in a hypo-osmotic solution (500 µL at 75 mOsm/kg) containing fructose (6.75 g/L) and sodium citrate (3.67 g/L) in H
2O. A sample of the mixture was placed on a slide and covered with a coverslip. The number of spermatozoa with coiled or swollen tails (indicative of intact membranes under hypoosmotic conditions) among 300 sperm per slide was counted in each sample at 400× under phase-contrast microscopy.
2.8.5. Antioxidant Capacity and Enzyme Activities
The following biochemical parameters were measured in post-thawed extender: total antioxidant capacity (TAC, linearity up to 2 mM/L) [
20], hydrogen peroxide concentration (H
2O
2, linearity up to 1.5 mM/L) [
21], lactic dehydrogenase (LDH, linearity up to 1700 units/L) activity [
22], aspartate transaminase (AST, linearity up to 150 units/mL) activity, and alanine transaminase (ALT, linearity up to 120 units/mL) activity [
23]. All measurements were performed using a spectrophotometer (Spectro UV-VIS Auto, UV-2602, Labomed, Los Angeles, CA, USA) and commercial kits (Biodiagnostic, Giza, Egypt) according to the manufacturer’s instructions.
2.8.6. Apoptosis and Necrosis
Semen samples were stained with Annexin-V (AV, calcium-dependent probe) for tracking phosphatidylserine (PS) externalization in the membrane and propidium iodide (PI) as an indicator of genomic DNA exposure using a commercial PS Detection Kit (IQP, Groningen, The Netherlands) according to the manufacturer’s instructions. Briefly, semen samples were thawed and washed twice by centrifugation (300×
g for 10 min at 4 °C) with phosphate-buffered saline. After the second centrifugation, the supernatant was removed, and the sperm pellet resuspended in binding buffer at 1 × 10
6 sperm cells/mL. Then, 100 μL of semen sample was transferred to culture tubes (5 mL) containing 5 μL AV (fluorescein isothiocyanate, FITC label, BD Biosciences, San Jose, CA, USA) and 5 μL PI (BD Biosciences). The mixed suspension was then incubated in the dark at room temperature (25 °C) for 15 min, followed by the addition of 400 µL binding buffer to each tube. Staining patterns were then evaluated by flow cytometry using an Accuri C6 Cytometer and Accuri C6 software (BD Biosciences) [
24]. Cells negative for both AV and PI staining (A−/PI−) were classified as viable, those positive for AV and negative for PI (A+/PI−) as early apoptotic, those positive for both AV and PI (A+/PI+) as apoptotic, and those negative for AV and positive for PI (A−/PI+) as necrotic.
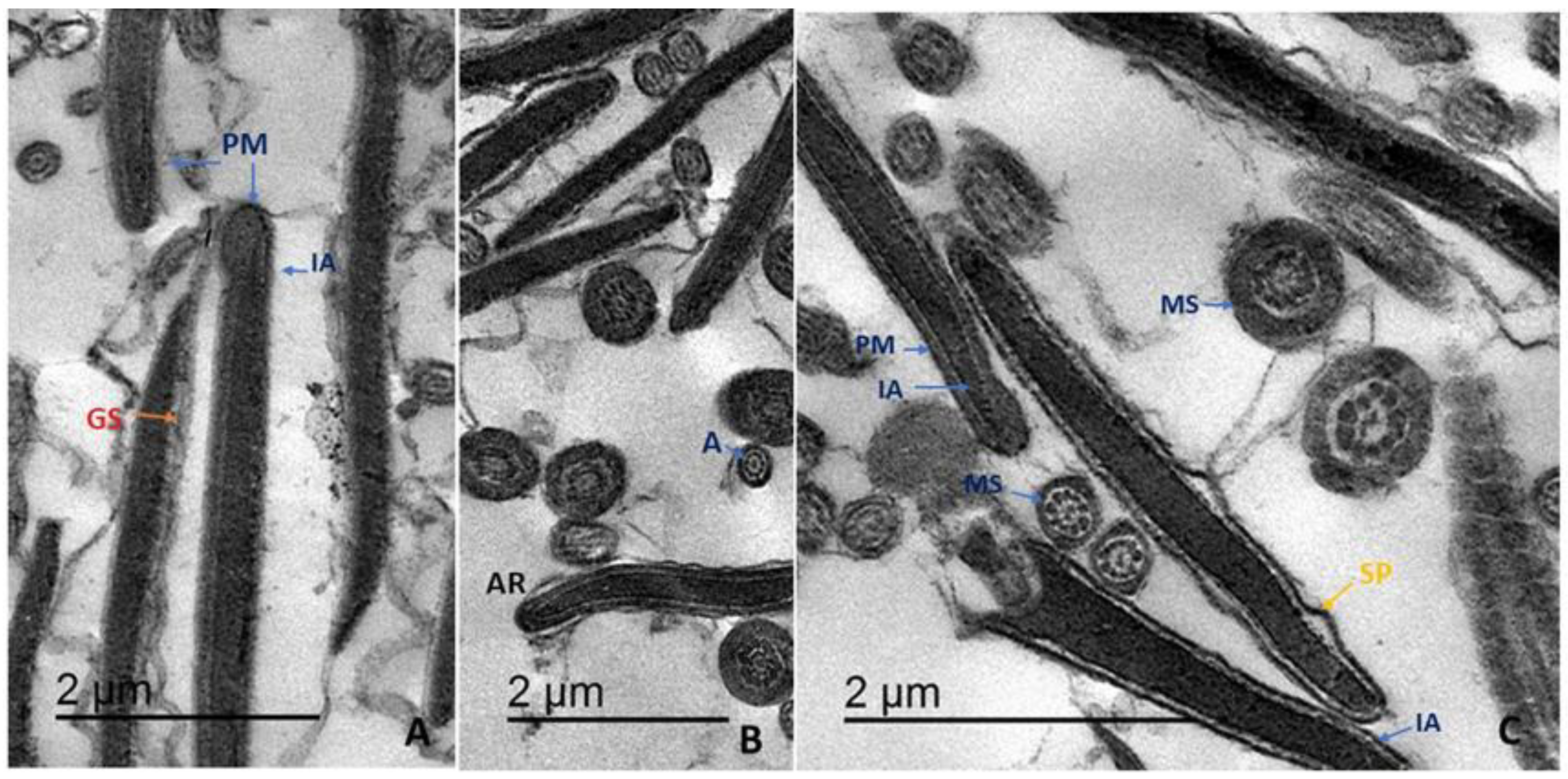
2.8.7. Ultrastructure
Sperm ultrastructure was examined by transmission electron microscopy (TEM) as described [
25] with some modifications. Briefly, 500 μL of each semen sample was centrifuged and resuspended in cold (4 °C) fixative solution (2.5% glutaraldehyde in phosphate buffer) for 2 h. Samples were then washed and post-fixed in osmium tetroxide (1%) for 90 min at room temperature, dehydrated, cleared in gradient ethanol and propylene oxide, and embedded in Epon 812 (Fluka Chemie, AG, Buchs, Switzerland). Ultrathin sections (60–70 nm) were prepared using glass knives and observed using a JEOL-JEM 2100 TEM at 80 kV. Changes in PM and acrosome ultrastructure were examined from 300 sperm per sample.
2.9. Statistical Analysis
Arcsine transformation was performed before statistical analyses because that helps in dealing with percentage values for semen characteristics including progressive motility, viability, membrane integrity, acrosome integrity, structural abnormality, sperm viability by Annexin-V, plasma membrane integrity, acrosomal ultrastructure. Treatment group means were compared by one-way analysis of variance (ANOVA) [
26] and Duncan’s multiple range tests [
27]. A
p < 0.05 was considered statistically significant for all tests.
4. Discussion
Supplementation of goat semen extender with 375 µg/mL TFLE improved the functional and ultrastructural characteristics of cryopreserved sperm by maintaining antioxidant capacity, thereby preventing membrane injury and reducing apoptosis.
Semen cryopreserved in semen extender alone (control group) demonstrated the lowest proportion of viable sperm compared to samples preserved with extender containing varying concentrations of the leaf extract. Freezing and thawing can damage cellular membranes, reducing sperm number and quality as shown by flow cytometry and TEM. Our results are in accord with previous studies demonstrating the harmful effects of thawing on sperm structure and function. Oxidative stress during cryopreservation reduces the reproductive potential of semen by impairing sperm motility, reducing mitochondrial activity, damaging DNA, and activating apoptotic pathways [
8,
9,
29]. Thus, supplementation of semen extender with antioxidants prior to cryopreservation is recommended to facilitate efficient goat breeding [
30]. Here, we demonstrate improved cryopreservation using a natural plant extract containing multiple bioactive agents with known beneficial effects against cellular stress.
Freezing and thawing induced apoptosis as evidenced by Annexin staining, which reveals the translocation of phosphatidylserine from the inner to the outer PM layer. Further, some sperm cells became necrotic during cryopreservation as evidenced by PI staining [
31]. Both apoptosis and necrosis are associated with loss of PM integrity, which is necessary to maintain sperm function within the female reproductive tract [
32]. During cryopreservation, rearrangement of membrane lipids alters fluidity and increases susceptibility to disruption, which then induces further cellular damage and ultimately death [
33]. These pathological effects are manifested by changes in sperm morphology during the freezing-thawing process [
34]. High concentrations of polyunsaturated fatty acids (PUSFAs) such as arachidonic and docosahexaenoic acids in the PM increase the vulnerability to ROS-induced peroxidative damage and membrane dysfunction [
35]. Further, oxidative injury may be spread throughout the spermatozoa population by a subset of cells overproducing ROS [
36], leading to generally reduced mitochondrial metabolic activity, motility, and viability [
37]. Maintenance of cell membrane integrity and mitochondrial function under oxidative stress are thus essential for successful fertilization using cryopreserved semen [
33].
Following cryopreservation, damage to the PM and acrosomal cap was predominantly observed in the head region, in accordance with previous observations of human sperm [
28]. Membrane swelling is most probably caused by changes in the extracellular osmotic pressure during freezing and thawing, causing cells to accumulate or lose water. The sperm PM is known to mediate the exchange of sodium, potassium [
38,
39], and calcium [
40], and these ion fluxes regulate motility and mitochondrial function as well as osmotic balance. An intact PM is also necessary for fusion with the outer acrosomal membrane and induction of the acrosome reaction [
41]. Acrosomal integrity is also essential for fertilization as this organelle contains hydrolytic enzymes such as hyaluronidase, acrosin, and esterases required for lysis of the zona pellucida and penetration of the oocyte corona radiata [
42]. Freezing and thawing significantly increased the number of sperm cells with atypical acrosomal structure, which has previously been attributed to degeneration and apoptosis [
43].
The addition of TFLE to sperm extender dose-dependently increased the TAC of goat semen and reduced the concentration of H
2O
2, a major ROS generator. On the other hand, TFLE had little effect on the activities of LDH, AST, and ALT. The dose-dependent increase in TAC was strongly associated with the progressive rise in sperm cell viability and the decreases in apoptosis, necrosis, and structural abnormalities. Consistent with these findings, Salimi, et al. [
44] reported positive correlations between TAC and both sperm motility and normal sperm morphology, while Pahune, et al. [
45] observed positive correlations between TAC and multiple seminogram parameters including sperm concentration, sperm motility, and normal sperm morphology. Collectively, these findings suggest that an imbalance between TAC and ROS production is a major contributor to impaired sperm function following cryopreservation [
46].
Mitochondrial enzymatic activities in human spermatozoa are strongly correlated with motility [
47]. Aspartate transaminase (AST) and ALT are essential for metabolic processes that provide energy for sperm survival, motility, and fertility [
48], and so are good indicators of sperm membrane stability and semen quality [
49]. An increase in spermatozoa damage within the liquid storage medium results in an elevated concentration of transaminase enzymes [
50]. Indeed, AST and ALT activities were slightly higher in control samples than samples containing 375 µg/mL TFLE, although the difference did not reach statistical significance.
TFLE contains secondary metabolites such as malic acid, quercetin, and kaempferol that may contribute to these improved functional and structural characteristics. Indeed, malic acid decreases the accumulation of ROS and enhances the glutathione cycle by regulating various endogenous antioxidant pathways [
51,
52], while the flavonoids can directly scavenge ROS, thereby resisting oxidative damage during cryopreservation [
53]. Finally, kaempferol is a flavonoid compound with potent activity against inflammation caused by oxidative stress [
54].
The effect of TFLE on semen cryopreservation was stronger compared with other plant extracts such as
Albizia harveyi leaf extract in bull [
8],
Entada abyssinica bark extract in ram [
9], and nanoformulations of mint, thyme, and curcumin in goat [
10]. All these extracts enhanced semen preservability and sperm characteristics after freezing and thawing.
TFLE is rich in phenolic and flavonoid compounds, which have antioxidant properties. This is evident in the antioxidant activity of the extract via DPPH, FRAP, and TPC assays or by reducing the concentration of hydrogen peroxide in the semen extender after thawing. This can be explained by the ability of polyphenolic compounds to scavenge reactive oxygen species such as superoxide anion radicals and hydroxyl radicals, thus interrupting free radical chain reaction [
55]. Therefore, we expect the same effect in preserving the semen in other species.


 ,
, 




{kind=link}
{kind=link}