
The Tongue in Three Species of Lemurs: Flower and Nectar Feeding Adaptations

 , ,
, ,  , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
1.1. Nectar Feeding
1.2. The Tongue in Prosimian Primates
1.3. Gross Anatomy of Tongue
1.4. The Tongue’s Papillae
1.5. Aim of the Work
2. Materials and Methods
2.1. Specimen Acquisition and Preparation
2.2. Macroscopically Observation
2.3. Fixation and Processing for Histological General Stains (Hematoxylin–Eosin and Masson Trichrome)
2.4. Single Immunohistochemistry
2.5. Ultrastructural Study Using Scanning Electron Microscopy (SEM)
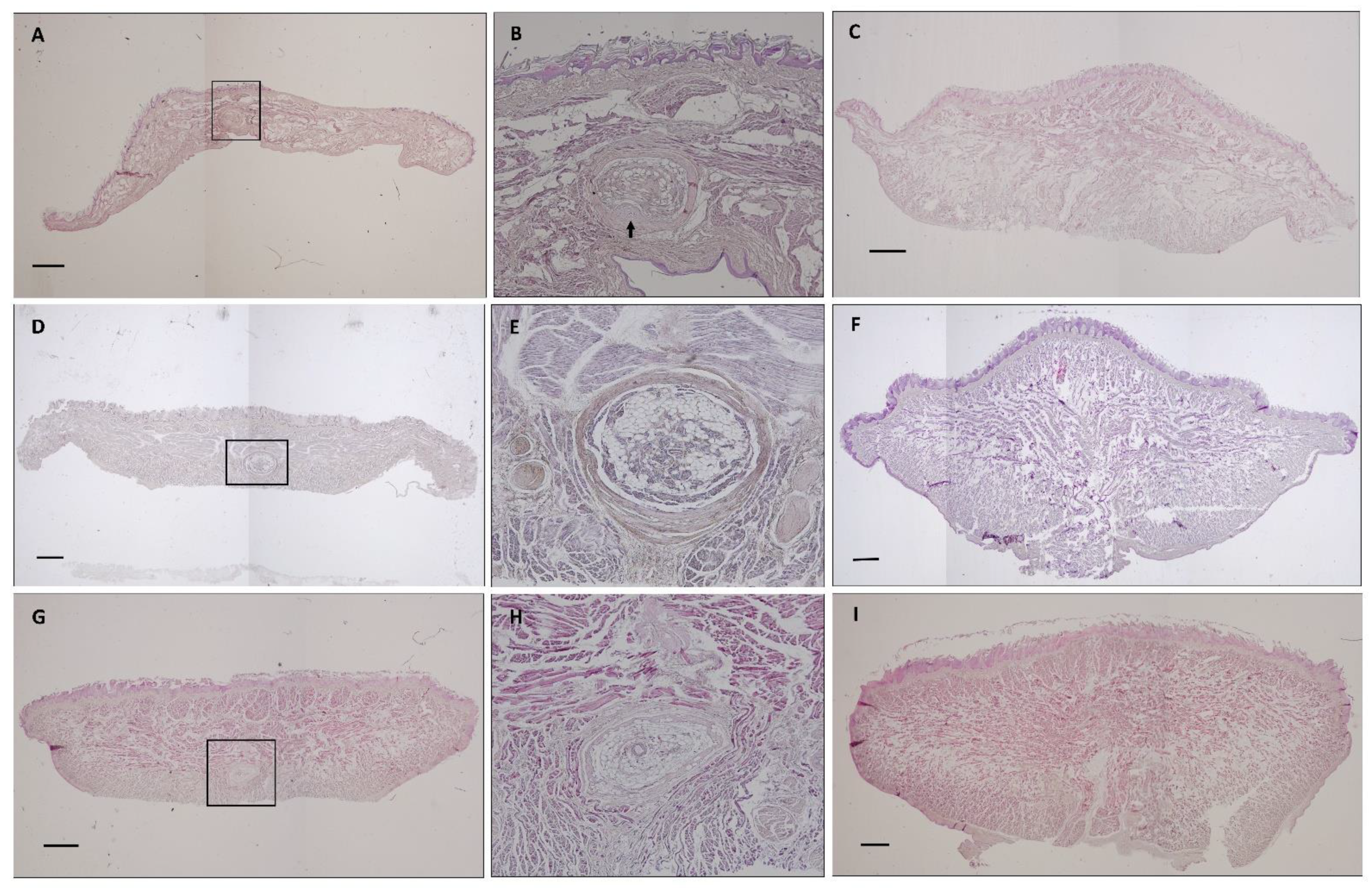
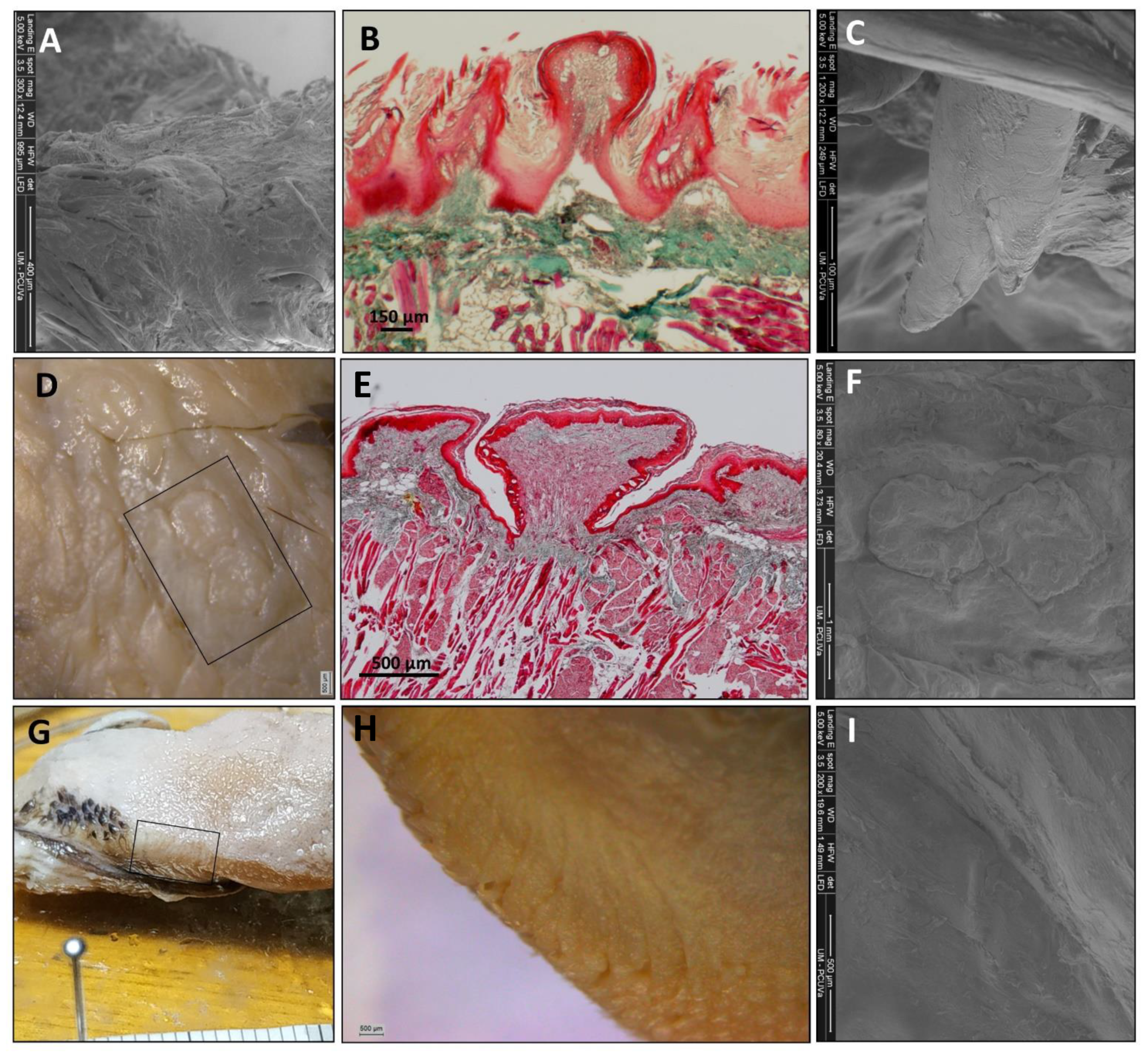
3. Results
3.1. The Tongue
3.1.1. General Observation
3.1.2. The Tongue’s Tip
3.1.3. Papillae Location and Morphology
3.2. The Sublingua
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jordano, P. Coevolution in multispecific interactions among free-living species. Evol. Educ. Outreach 2010, 3, 40–46. [Google Scholar] [CrossRef] [Green Version]
- Thompson, J.N. Four central points about coevolution. Evol. Educ. Outreach 2010, 3, 7–13. [Google Scholar] [CrossRef] [Green Version]
- Dew, J.L.; Wright, P. Frugivory and seed dispersal by four species of primates in Madagascar’s eastern rain forest. Biotropica 1998, 30, 425–437. [Google Scholar] [CrossRef]
- Chapman, C.A.; Onderdonk, D.A. Forests without primates: Primate/plant codependency. Am. J. Primatol. 1998, 45, 127–141. [Google Scholar] [CrossRef]
- Sussman, R.W.; Raven, P.H. Pollination by lemurs and marsupials: An archaic coevolutionary system. Science 1978, 200, 731–736. [Google Scholar] [CrossRef] [PubMed]
- Garber, P.A. Foraging decisions during nectar feeding by tamarin monkeys (Saguinus mystax and Saguinus fuscicollis Callitrichidae, Primates) in Amazonian Peru. Biotropica 1988, 20, 100–106. [Google Scholar] [CrossRef]
- Terborgh, J.W.; Janson, C. The socioecology of primate groups. Ann. Rev. Ecol. Syst. 1986, 17, 111–136. [Google Scholar] [CrossRef]
- Muchlinski, M.N.; Perry, J.M. Anatomical Correlates to Nectar Feeding among the Strepsirrhines of Madagascar: Implications for Interpreting the Fossil Record. Anat. Res. Int. 2011, 2011, 378431. [Google Scholar] [CrossRef] [Green Version]
- Overdorff, D.; Johnson, S.E. Eulemur, true lemurs. In The Natural History of Madagascar; Goodman, S.M., Benstead, J.P., Eds.; University of Chicago Press: Chicago, IL, USA, 2003; pp. 1320–1324. [Google Scholar]
- Janson, C.; Terborgh, J.; Emmons, L. Non-flying mammals as pollinating agents in the Amazonian forest. Reprod. Bot. 1981, 14, 1–6. [Google Scholar] [CrossRef]
- Heinrich, B.; Raven, P.H. Energetics and pollination ecology. Science 1972, 176, 597–602. [Google Scholar] [CrossRef]
- Kress, W.J. Coevolution of plants and animals: Pollination of flowers by primates in Madagascar. Curr. Sci. 1993, 65, 253–257. [Google Scholar]
- Kress, W.J.; Schatz, G.E.; Andrianifahanana, M.; Morland, H.S. Pollination of Ravenala madagascariensis (Strelitziaceae) by lemurs in Madagascar: Evidence for an archaic coevolutionary system? Am. J. Bot. 1994, 81, 542–551. [Google Scholar] [CrossRef]
- Overdorff, D.J. Differential patterns in flower feeding by Eulemur fulvus rufus and Eulemur rubriventer in Madagascar. Am. J. Primatol. 1992, 28, 191–203. [Google Scholar] [CrossRef] [PubMed]
- Terborgh, J. Community aspects of frugivory in tropical forests. In Frugivores and Seed Dispersal; Estrada, A., Fleming, T., Eds.; W. Junk Publishing: Dordrecht, The Netherlands, 1986; pp. 372–384. [Google Scholar]
- Wright, P.C.; Martin, L. Predation, pollination and torpor in two nocturnal prosimians: Cheirogaleus major and Microcebus rufus in the rain forest of Madagascar. In Creatures of the Dark: The Nocturnal Prosimians; Alterman, L., Doyle, G., Izard, M.K., Eds.; Plenum: New York, NY, USA, 1995; pp. 45–60. [Google Scholar]
- Sussman, R. Nectar-feeding by prosimians and its evolutionary and ecological implications. In Recent Advances in Promatology; Chivers, D., Joysey, K., Eds.; Academic Press: New York, NY, USA, 1978; pp. 119–126. [Google Scholar]
- Rasmussen, D.; Sussman, R. Parallelisms among primates and possums. In Primate Origins: Adaptations and Evolution; University of Chicago Press: Chicago, IL, USA, 2007; pp. 775–803. [Google Scholar]
- Rasmussen, M. Ecological influences on activity cycle in two cathemeral primates, the mongoose lemur. In Biological Anthropology and Anatomy; Duke University: Durham, NC, USA, 1999. [Google Scholar]
- Tecot, S. Seasonality and Predictability: The Hormonal and Behavioral Responses of the Red-Bellied Lemur, Eulemur Rubriventer, in Southeastern Madagascar. Ph.D. Thesis, University of Texas, Austin, TX, USA, 2008. [Google Scholar]
- Birkinshaw, C.; Colquhoun, I.C. Pollination of Ravenala madagascariensis and Parkia madagascariensis by Eulemur macaco in Madagascar. Folia Primatol. 1998, 69, 252–259. [Google Scholar] [CrossRef]
- Colquhoun, I. A Predictive Socioecology Study of the Black Lemur (Eulemur Macaco Macaco) in Northwestern Madagascar. Ph.D. Thesis, Washington University, St. Louis, MO, USA, 1997; p. 274. [Google Scholar]
- Sussman, R.W. Primate origins and the evolution of angiosperms. Am. J. Primatol. 1991, 23, 209–223. [Google Scholar] [CrossRef] [PubMed]
- Sussman, R. How primates invented the rainforest and vice versa. In Creatures of the Dark: The Nocturnal Prosimians; Alterman, L., Doyle, G., Izard, M., Eds.; Plenum Press: New York, NY, USA, 1995. [Google Scholar]
- Howell, D.; Hodgkin, N. Feeding adaptations in the hairs and tongues of nectar feeding bats. J. Morphol. 1976, 148, 329–336. [Google Scholar] [CrossRef] [PubMed]
- Tschapka, M.; Gonzalez-Terrazas, T.P.; Knörnschild, M. Nectar uptake in bats using a pumping-tongue mechanism. Sci. Adv. 2015, 1, e1500525. [Google Scholar] [CrossRef] [Green Version]
- Casali, D.M.; Martins-Santos, E.; Santos, A.L.Q.; Miranda, F.R.; Mahecha, G.A.B.; Perini, F.A. Morphology of the tongue of Vermilingua (Xenarthra: Pilosa) and evolutionary considerations. J. Morphol. 2017, 278, 1380–1399. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, S.I.; Yoshimura, K.; Shindo, J.; Kageyami, I. Comparative morphology of the primate tongue. Ann. Anat. 2019, 223, 19–31. [Google Scholar] [CrossRef]
- Jones, F.W. The Sublingua and the Plica Fimbriata. J. Anat. 1918, 52, 345–353. [Google Scholar]
- Pocock, R.I. On the external characters of the lemurs and of Tarsius. Proc. Zool. Soc. Lond. 1918, 19–53. [Google Scholar] [CrossRef]
- Moore, K.L. Clinically Oriented Anatomy, 7th ed.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 1999; ISBN 9781451119459. [Google Scholar]
- Sonntag, C.F. The comparative anatomy of the tongues of the mammalian: I General description of the tongue. Proc. Zool. Soc. Lond. 1920, IX, 115–129. [Google Scholar]
- Sonntag, C.F. The comparative anatomy of the tongues of the mammalia: V Family 5. Lemuroidea and Tarsioideae. Proc. Zool. Soc. Lond. 1921, L, 741–754. [Google Scholar]
- Buck, L.M. Smell and taste: The chemical senses. In Principals of Neural Science; Kandel, E.R., Schwartz, J.H., Jessell, T.M., Eds.; McGraw-Hill: New York, NY, USA, 2000; pp. 625–647. [Google Scholar]
- Alport, L.J. Lingual Fungiform Papillae and the Evolution of the Primate Gustatory System. Ph.D. Thesis, University of Texas, Austin, TX, USA, 2009. [Google Scholar]
- Barone, R. Anatomie compareé des mamifères domestiques: Tome troisième, Splanchnologie, fœtus et ses annexes. Bull. Acad. Vet. Fr. 1976, 129, 483–484. (In French) [Google Scholar]
- Purves, D.; Fitzpatric, D.; Katz, L.C.; LaMantia, A.S.; McNamara, J.O. (Eds.) Neuroscience; Sinaur Associates, Inc.: Sunderland, MA, USA, 1997. [Google Scholar]
- Machida, H.; Perkins, E.; Giacometti, L. The anatomical and histochemical properties of the tongue of primates. Folia Primatol. 1967, 5, 264–279. [Google Scholar] [CrossRef]
- Iwasaki, S. Evolution of the structure and function of the vertebrate tongue. J. Anat. 2002, 201, 1–13. [Google Scholar] [CrossRef]
- Hofer, H.O. Microscopic anatomy of the apical part of the tongue of Lemur fulvus (Primates, Lemuriformes). Gegenbaurs Morphol. Jahrb. 1981, 127, 343–363. (In German) [Google Scholar]
- Takemurao, A.; Ikeo, H.; Toda, I.; Tamadao, Y.; Mouri, T.; Shigehara, N.; Suwai, S.E.M. Studies of the Lingual Papillae of the Ring-tailed Lemur (Lemur catta)—The Filiform and the Fungiform Papillae. Primate Res. 2002, 18, 35–42. (In Japanesse) [Google Scholar] [CrossRef]
- Fleagle, J.G. Primate Adaptation and Evolution, 3rd ed.; Academic Press: New York, NY, USA, 2013. [Google Scholar]
- Harper, C.J.; Swartz, S.M.; Brainerd, E.L. Specialized bat tongue is a hemodynamic nectar mop. Proc. Natl. Acad. Sci. USA 2013, 110, 8852–8857. [Google Scholar] [CrossRef] [Green Version]
- Nuessle, T.M.; Garneau, N.L.; Sloan, M.M.; Santorico, S.A. Denver Papillae Protocol for Objective Analysis of Fungiform Papillae. J. Vis. Exp. 2015, 100, e52860. [Google Scholar] [CrossRef] [Green Version]
- Yoshimura, K.; Ono, K.; Shindo, J.; Iwasaki, S.-I.; Kageyama, I. Comparative morphology of the lingual papillae and their connective tissue cores in the tongue of the Abyssinian black-and-white colobus (Colobus guereza). Anat. Sci. Int. 2019, 94, 225–237. [Google Scholar] [CrossRef]
- Pastor, J.F.; Barbosa, M.; de Paz, F.J.; San José, I.; Levanti, M.; Potau, J.M.; Vega, J.A.; Cabo, R. Ultrastructure of Lingual Papillae in Common Chimpanzee (Pan troglodytes) Foetus, Newborn and Adult Specimens. Anat. Histol. Embryol. 2017, 46, 431–438. [Google Scholar] [CrossRef]
- Suárez-Quintanilla, J.; Martín-Cruces, J.; Martín-Biedma, B.; Cobo, J.L.; Mérida-García, A.J.; de Vicente, J.C.; Junquera, L.M.; Mérida-Velasco, J.R.; Vega, J.A. The juxta-oral organ of Chievitz (organum yuxtaorale) updated: Embryology, anatomy, function and pathology. Ann. Anat. 2020, 232, 151582. [Google Scholar] [CrossRef] [PubMed]
- D’Andrea, V.; Malinovsky, L.; Biancari, F.; Spyrou, M.; Dibra, A.; Malinovska, V.; Hees, H.; Gallenzi, P.; Cristalli, M.P.; Di Matteo, F.M.; et al. The Chievitz juxtaparotid organ. G. Chir. 1999, 20, 213–217. [Google Scholar] [PubMed]
- Mérida-Velasco, J.R.; Rodríguez-Vázquez, J.F.; de la Cuadra-Blanco, C.; Salmerón, J.I.; Sánchez-Montesinos, I.; Mérida-Velasco, J.A. Morphogenesis of the juxtaoral organ in humans. J. Anat. 2005, 206, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Geoffroy Saint-Hilaire, E.; Cuvier, F. Histoire naturelle des mammifères: Avec des figures originales, coloriées, dessinées d’aprèsdes animaux vivans. Hist. Nat. Mammif. 1829, 218. [Google Scholar] [CrossRef] [Green Version]
- Anakel-Simons, F. Primate Anatomy: An Introduction, 3rd ed.; Academic Press: New York, NY, USA, 2007. [Google Scholar]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pastor, J.F.; Muchlinski, M.N.; Potau, J.M.; Casado, A.; García-Mesa, Y.; Vega, J.A.; Cabo, R. The Tongue in Three Species of Lemurs: Flower and Nectar Feeding Adaptations. Animals 2021, 11, 2811. https://doi.org/10.3390/ani11102811
Pastor JF, Muchlinski MN, Potau JM, Casado A, García-Mesa Y, Vega JA, Cabo R. The Tongue in Three Species of Lemurs: Flower and Nectar Feeding Adaptations. Animals. 2021; 11(10):2811. https://doi.org/10.3390/ani11102811
Chicago/Turabian StylePastor, Juan Francisco, Magdalena Natalia Muchlinski, Josep Maria Potau, Aroa Casado, Yolanda García-Mesa, Jose Antonio Vega, and Roberto Cabo. 2021. "The Tongue in Three Species of Lemurs: Flower and Nectar Feeding Adaptations" Animals 11, no. 10: 2811. https://doi.org/10.3390/ani11102811