
Telomere Length and Regulatory Genes as Novel Stress Biomarkers and Their Diversities in Broiler Chickens (Gallus gallus domesticus) Subjected to Corticosterone Feeding

 , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Management, Housing and Experimental Design
2.2. Diets and Corticosterone Challenge
2.3. Growth Rate Parameters and Animal Sampling
2.4. Plasma Corticosterone Level Determination
2.5. Determination of Telomere Length Using Real-Time Quantitative PCR (qRT-PCR) Analysis
2.6. Gene Expression Analysis of Telomere Length Regulatory Genes
2.7. Statistical Analysis
3. Result
3.1. Growth Performance and Mortality
3.2. Absolute Weight of Organs
3.3. Relative Weight of Organs
3.4. Plasma Level of Corticosterone
3.5. Absolute Telomere Length
3.6. Telomere Regulatory Genes
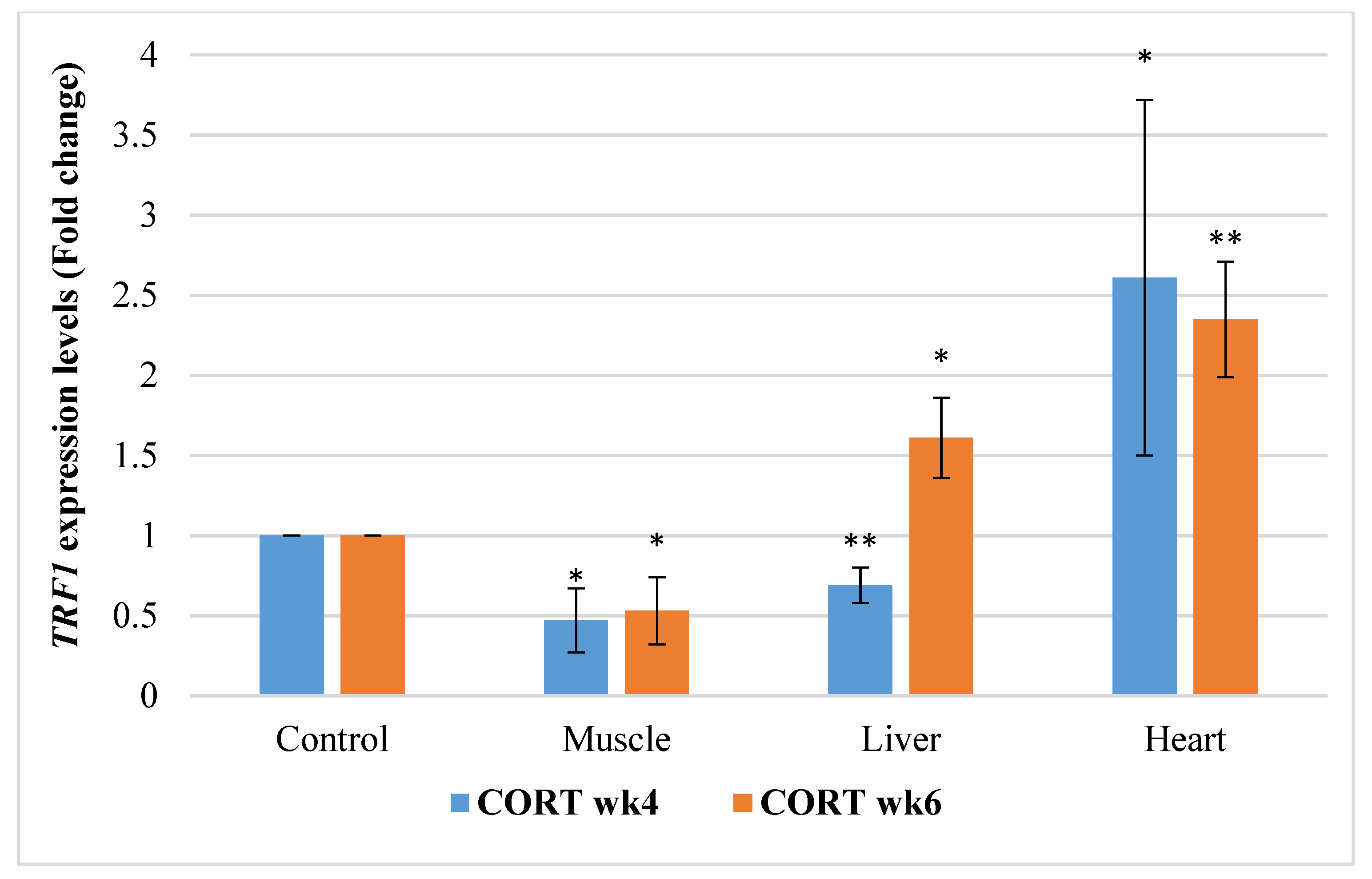
3.6.1. Telomeric Repeat Transcriptional Factor 1 (TRF1)
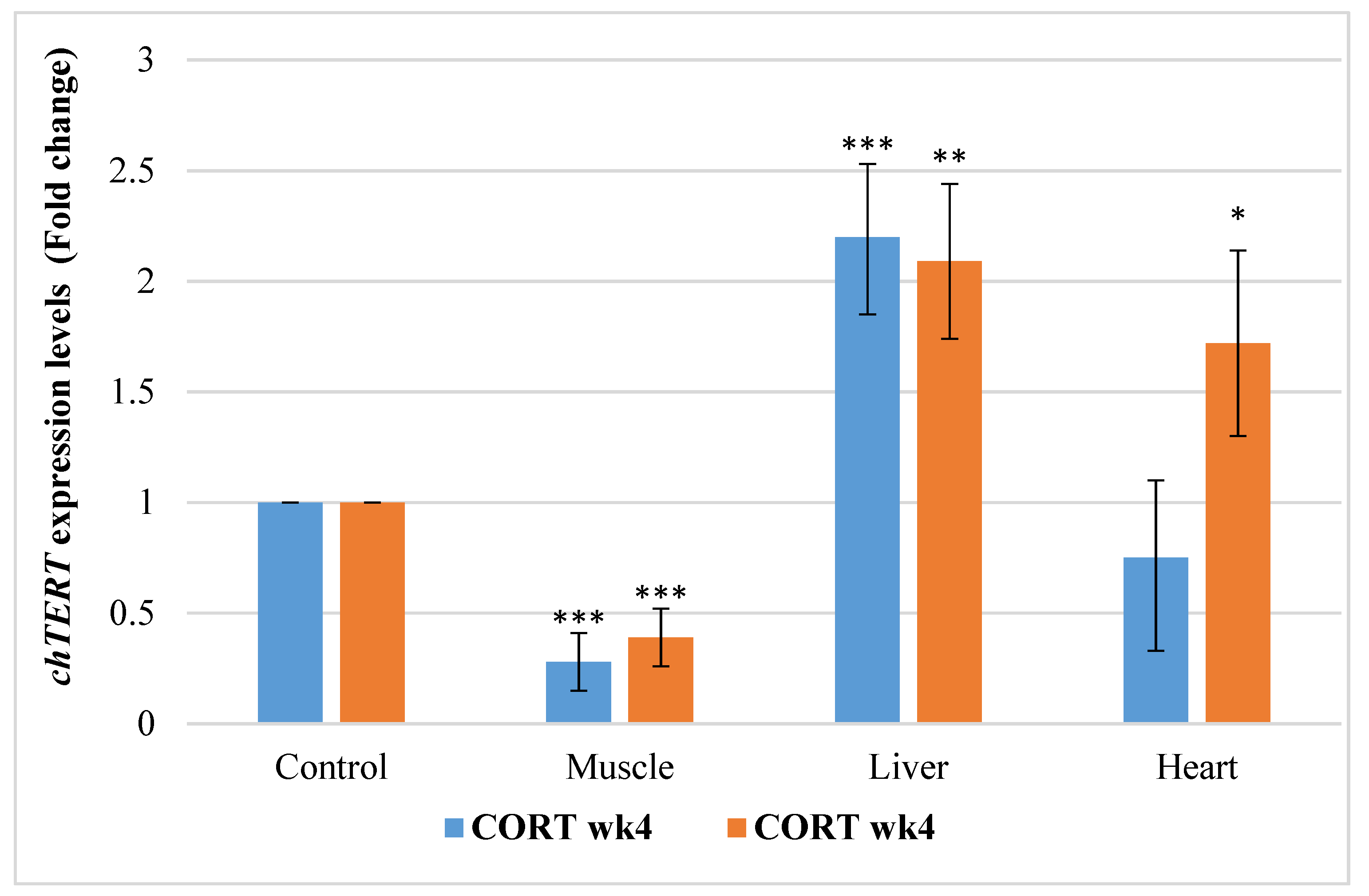
3.6.2. Chicken Telomerase Reverse Transcription Factors (chTERT)
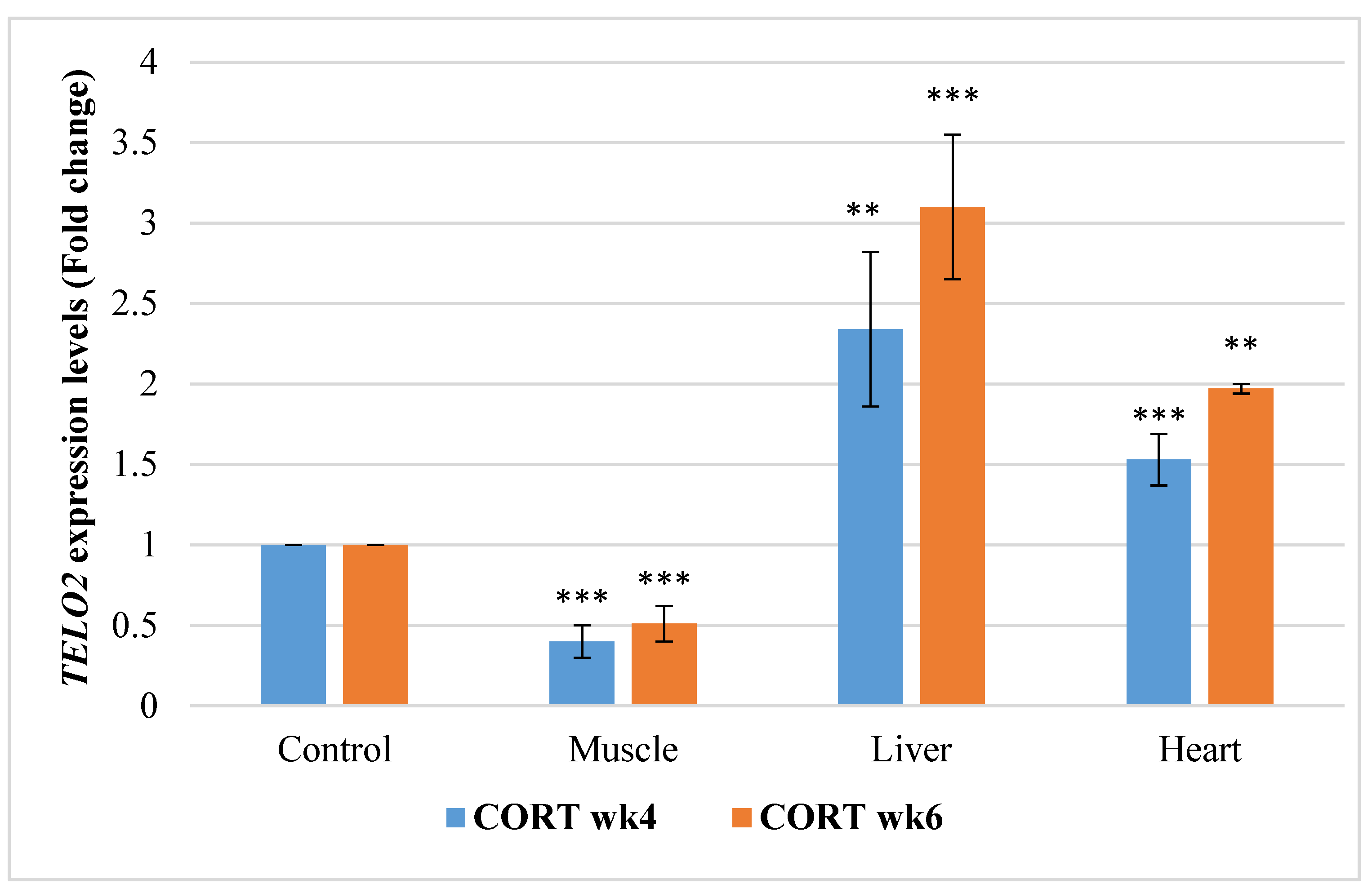
3.6.3. Telomere Maintenance Gene 2 (TELO2)
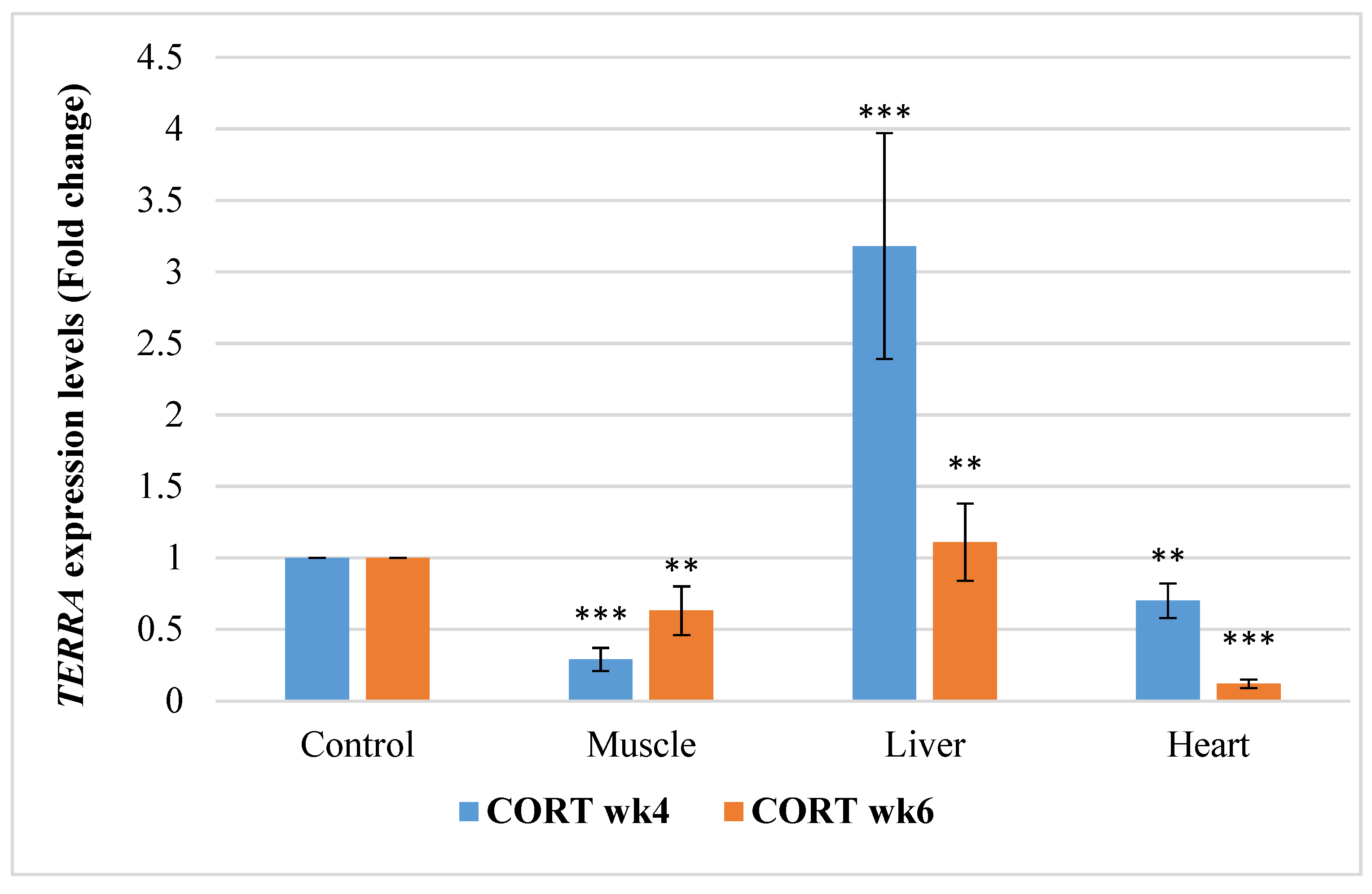
3.6.4. Telomeric Repeat-Containing RNA (TERRA)
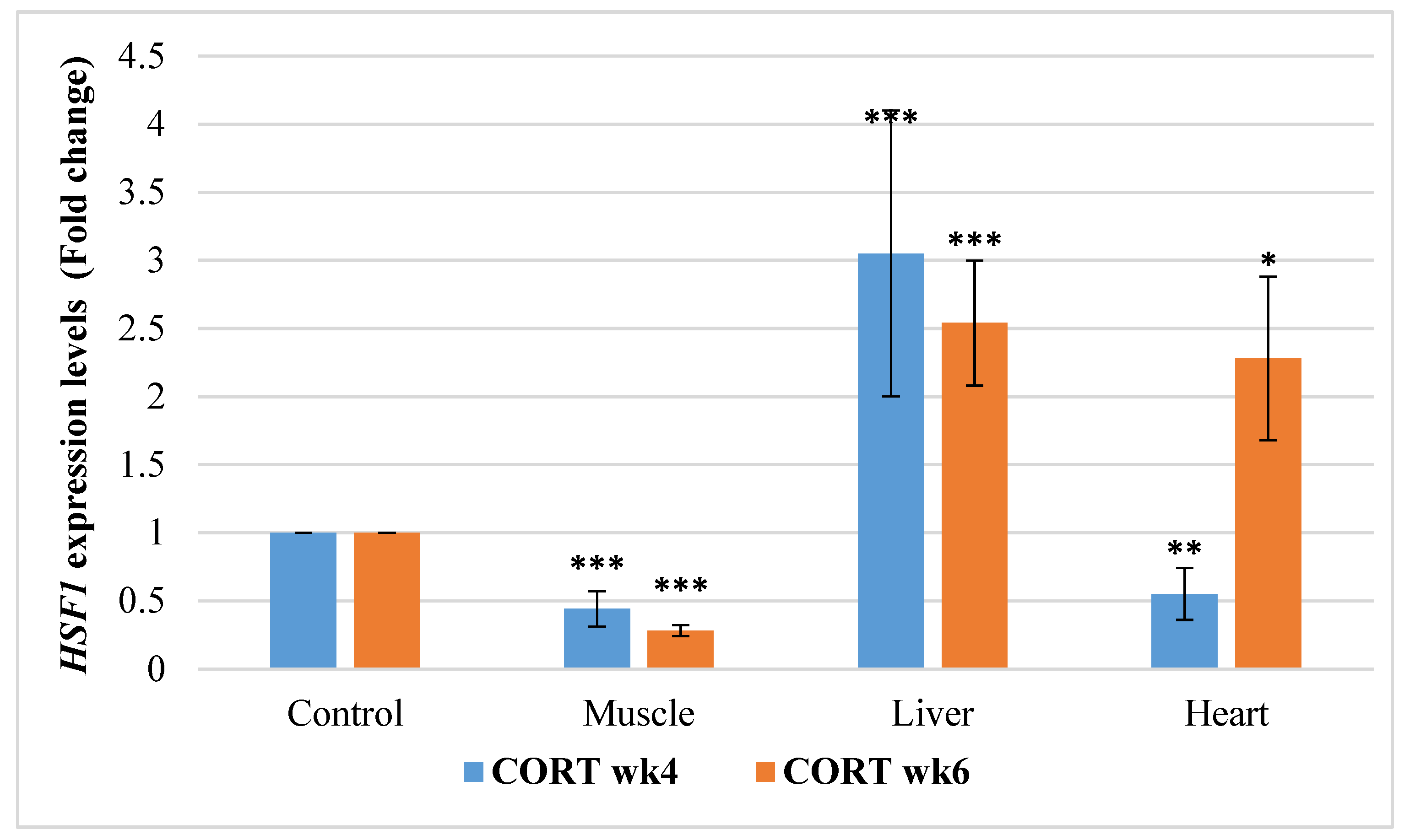
3.6.5. Heat Shock Transcriptional Factor 1 (HSF1)
4. Discussion
4.1. Growth Performance
4.2. Absolute and Relative Weights of Organ
4.3. Plasma Corticosterone Levels
4.4. Telomere Length
4.5. Telomere Length Regulatory Genes
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rushen, J. Problems associated with the interpretation of physiological data in the assessment of animal welfare. Appl. Anim. Behav. Sci. 1991, 28, 381–386. [Google Scholar] [CrossRef]
- Fairhurst, G.D.; Marchant, T.A.; Soos, C.; Machin, K.L.; Clark, R.G. Experimental relationships between levels of corticosterone in plasma and feathers in a free-living bird. J. Exp. Biol. 2013, 4071–4081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhabhar, F.S.; McEwen, B.S. Acute stress enhances while chronic stress suppresses cell-mediated immunity in vivo: A potential role for leukocyte trafficking. Brain Behav. Immun. 1997, 11, 286–306. [Google Scholar] [CrossRef] [Green Version]
- Dhabhar, F.S. Acute stress enhances while chronic stress suppresses skin immunity. The role of stress hormones and leukocyte trafficking. Ann. N. Y. Acad. Sci. 2000, 917, 876–893. [Google Scholar] [CrossRef]
- Romero, L.M. Physiological stress in ecology: Lessons from biomedical research. Trends Ecol. Evol. 2004, 19, 249–255. [Google Scholar] [CrossRef]
- Spiers, J.G.; Chen, H.C.; Sernia, C.; Lavidis, N.A. Activation of the hypothalamic-pituitary-adrenal stress axis induces cellular oxidative stress. Front. Neurosci. 2015, 8, 456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bennett, M.C.; Mlady, G.W.; Fleshner, M.; Rose, M.G. Synergy between chronic corticosterone and sodium azide treatments in producing a spatial learning deficit and inhibiting cytochrome oxidase activity. Proc. Natl. Acad. Sci. USA 1996, 93, 1330–1334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barna, J.; Csermely, P.; Vellai, T. Roles of heat shock factor 1 beyond the heat shock response. Cell. Mol. Life Sci. 2018, 75, 2897–2916. [Google Scholar] [CrossRef]
- Jones, T.J.; Li, D.; Wolf, I.M.; Wadekar, S.A.; Periyasamy, S.; Sánchez, E.R. Enhancement of glucocorticoid receptor-mediated gene expression by constitutively active heat shock factor 1. Mol. Endocrinol. 2004, 18, 509–520. [Google Scholar] [CrossRef] [Green Version]
- Houbon, J.M.J.; Moonen, H.J.J.; van Schooter, F.J.; Hageman, G.J. Telomere length assessment: Biomarkers of chronic oxidative state? Free Radic. Biol Med. 2007, 144, 235–246. [Google Scholar] [CrossRef] [PubMed]
- Moyzis, R.K.; Buckingham, J.M.; Cram, L.S.; Dani, M.; Deaven, L.L.; Jones, M.D.; Meyne, J.; Ratliff, R.L.; Wu, J.R. A highly conserved repetitive DNA sequence, (TTAGGG), present at the telomeres of human chromosomes. Proc. Natl. Acad. Sci. USA 1988, 85, 6622–6626. [Google Scholar] [CrossRef] [Green Version]
- Kotrschal, A.; Ilmonen, P.; Penn, D.J. Stress impacts telomere dynamics. Biol. Lett. 2007, 3, 128–130. [Google Scholar] [CrossRef]
- Angelier, F.; Vleck, C.M.; Holberton, R.L.; Marra, P.P. Telomere length, non-breeding habitat and return rate in male American redstarts. Funct. Ecol. 2013, 27, 342–350. [Google Scholar] [CrossRef]
- Heidinger, B.J.; Blount, J.D.; Boner, W.; Griffiths, K.; Metcalfe, N.B.; Monaghan, P. Telomere length in early life predicts lifespan. Proc. Natl. Acad. Sci. USA 2012, 109, 1743–1748. [Google Scholar] [CrossRef] [Green Version]
- Blackburn, E.H.; Chan, S.; Chang, J.; Fulton, T.B.; Krauskopf, A.; Mceachern, M.; Prescott, J.; Roy, J.; Smith, C.; Wang, H. Molecular manifestations and molecular determinants of telomere capping. Cold Spring Harb. Symp. Quant. Biol. 2000, 65, 253–263. [Google Scholar] [CrossRef]
- Richter, T.; Proctor, C. The role of intracellular peroxide levels on the development and maintenance of telomere-dependent senescence. Exp. Gerontol. 2007, 42, 1043–1052. [Google Scholar] [CrossRef] [PubMed]
- Sohn, S.H.; Subramani, V.K. Dynamics of telomere length in the chicken. World Poult. Sci. J. 2014, 70, 721–736. [Google Scholar] [CrossRef]
- Costantini, D.; Marasco, V.; Møller, A.P. A meta-analysis of glucocorticoids as modulators of oxidative stress in vertebrates. J. Comp. Physiol. Biol. 2011, 181, 447–456. [Google Scholar] [CrossRef] [PubMed]
- Von Zglinicki, T. Oxidative stress shortens telomeres. Trends Biochem. Sci. 2002, 27, 339–344. [Google Scholar] [CrossRef]
- Elia, M. Organ and tissue contribution to metabolic rate. In Energy Metabolism. Tissue Determinants and Cellular Corollaries; Kinney, J.M., Tucker, H.N., Eds.; Raven Press: New York, NY, USA, 1992; pp. 61–77. [Google Scholar]
- Holliday, M.A. Body composition and energy needs during growth. In Human Growth: A Comprehensive Treatise; Fulkner, F., Tanner, J.M., Eds.; Plenum Press: New York, NY, USA, 1986; pp. 101–117. [Google Scholar]
- Hsu, A.; Heshka, S.; Janumala, I.; Song, M.; Horlick, M.; Krasnow, N.; Gallagher, D. Larger mass of high-metabolic-rate organs does not explain higher resting energy expenditure in children. Am. J. Clin. Nutr. 2003, 1–3, 1506–1511. [Google Scholar] [CrossRef] [Green Version]
- Bianchi, A.; Smith, S.; Chong, L.; Elias, P.; de Lange, T. TRF1 is a dimer and bends telomeric DNA. EMBO J. 1997, 16, 1785–1794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Lange, T. Shelterin: The protein complex that shapes and safeguards human telomeres. Genes Dev. 2005, 19, 2100–2110. [Google Scholar] [CrossRef] [Green Version]
- Ye, J.Z.S.; Donigian, J.R.; van Overbeek, M.; Loayza, D.; Luo, Y.; Krutchinsky, A.N.; Chait, B.T.; de Lange, T. TIN2 binds TRF1 and TRF2 simultaneously and stabilizes the TRF2 complex on telomeres. J. Biol. Chem. 2004, 279, 47264–47271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blackburn, E.H. Structure and function of telomeres. Nature 1991, 350, 569–573. [Google Scholar] [CrossRef]
- Greider, C.W.; Blackburn, E.H. Identification of a specific telomere terminal transferase activity in Tetrahymena extracts. Cell 1985, 43, 405–413. [Google Scholar] [CrossRef]
- Lansdorp, P.M. Telomeres, stem cells, and hematology. ASH 50th anniversary review. Blood 2008, 111, 1759–1766. [Google Scholar] [CrossRef] [Green Version]
- Taylor, H.A.; Delany, M.E. Ontogeny of telomerase in chicken: Impact of downregulation on pre- and postnatal telomere length in vivo. Dev. Growth Diff. 2000, 42, 613–621. [Google Scholar] [CrossRef] [PubMed]
- Gaudet, P.; Livestock, M.S.; Lweis, S.E.; Thomas, P.D. Phylogenetic-based propagation of functional annotations, within the Gene ontology consortium. Brief. Bioinform. 2011, 12, 449–462. [Google Scholar] [CrossRef] [Green Version]
- Graf, M.; Bonetti, D.; Lockhart, A.; Serhal, K.; Kellner, V.; Maicher, A.; Joilivet, P.; Teixeira, M.T.; Luke, B. Telomere length determines TERRA and R-loop regulation through the cell cycle. Cell 2017, 170, 72–85. [Google Scholar] [CrossRef] [Green Version]
- Koskas, S.; Decottignies, A.; Dufour, S.; Pezet, M.; Verdel, A.; Vourch, C.; Faure, V. Heat shock factor 1 promotes TERRA transcription and telomere protection upon heat stress. Nucleic Acids Res. 2017, 45, 6321–6333. [Google Scholar] [CrossRef] [Green Version]
- Blasco, M.; Schoeftner, S. Developmentally regulated transcription of mammalian telomeres by DNA-dependent RNA polymerase II. Nat. Cell Biol. 2008, 10, 228–236. [Google Scholar]
- Romano, G.H.; Harari, Y.; Yehuda, T.; Podhorzer, A.; Rubinstein, L.; Shamir, R.; Gottlieb, A.; Silberberg, Y.; Pe’er, D.; Ruppin, E.; et al. Environmental stresses disrupt telomere length homeostasis. PLoS Genet. 2013, 9, e1003721. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.F.; Guo, Y.M.; Huang, B.Y.; Zhang, L.B.; Bun, S.; Liu, D. Effect of corticosterone administration on small intestinal weight and expression of small intestinal nutrient transporter mRNA of broiler chickens. Asian Aust. J. Anim. Sci. 2010, 23, 175–181. [Google Scholar] [CrossRef]
- Cawthon, R.M. Telomere measurement by quantitative PCR. Nucleic Acids Res. 2002, 30, e47. [Google Scholar] [CrossRef]
- Herborn, K.A.; Heidinger, B.J.; Boner, W.; Noguera, J.C.; Adam, A.; Daunt, F.; Monaghan, P. Stress exposure in early post-natal life reduces telomere length: An experimental demonstration in a long-lived seabird. Proc. R. Soc. Lond. 2014, 281, 20133151. [Google Scholar] [CrossRef] [Green Version]
- O’Callaghan, N.J.; Fenech, M. A quantitative PCR method for measuring absolute telomere length. BMC 2011, 13, 3. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- SAS Publishing. Statistical Analysis System Multiple Incorporation. Users Guide Statistical Version; SAS Institute Inc.: Cary, NC, USA, 2002. [Google Scholar]
- Gross, W.B.; Siegel, P.B. Some effects of feeding deoxycorticosterone to chickens. Poult. Sci. 1981, 60, 2232–2239. [Google Scholar] [CrossRef]
- Zulkifli, I.; Najafi, P.; Nurfarahin, A.J.; Soleimani, A.F.; Kumari, S.; Anna Aryan, A.; O’ Reilly, E.L.; Eckersalli, P.D. Acute phase proteins, interleukin 6, and heat shock protein 70 in broiler chickens administered with corticosterone. Poult. Sci. 2014, 93, 3112–3118. [Google Scholar] [CrossRef]
- Lin, H.; Sui, J.; Jiao, H.; Buyse, H. Impaired development of broiler chickens by stress mimicked by corticosterone exposure. Comp. Biochem. Phys. A 2006, 143, 400–405. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, K.; Kaneda, S.; Otsuka, A.; Tomita, Y. Effects of ambient temperature and thyroxine on protein turnover and oxygen consumption inchicken squeletal muscle. In Proceedings of the 19th World’s Poultry Congress, Amsterdam, The Netherlands, 19–24 September 1992; Volume 2, pp. 93–96. [Google Scholar]
- Gross, W.B.; Siegel, P.B.; DuBose, R.T. Some effects of feeding corticosterone to chicken. Poult. Sci. 1980, 59, 516–522. [Google Scholar] [CrossRef]
- Ziegler, T.R.; Evans, M.E.; Fernandez-Estivariz, C.; Jones, D.P. Trophic and cytoprotective nutrition for intestinal adaptation, mucosal repair, and barrier function. Annu. Rev. Nutr. 2003, 23, 229–261. [Google Scholar] [CrossRef]
- Vágási, C.I.; Pătraș, L.; Pap, P.L.; Vincze, O.; Mureșan, C.; Németh, J.; Lendvai, Á.Z. Experimental increase in baseline corticosterone level reduces oxidative damage and enhances innate immune response. PONE 2018, 13, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Kawanishi, S.; Oikawa, S. Mechanism of telomere shortening by oxidative stress. Ann. N. Y. Acad. Sci. 2004, 1019, 278–284. [Google Scholar] [CrossRef] [PubMed]
- Kurt, W.; Vera, E.; Martinez-nevado, E.; Sanpera, C.; Blasco, M.A. Telomere shortening rate predicts species life span. Proc. Natl. Acad. Sci. USA 2019, 116, 15122–15127. [Google Scholar] [CrossRef] [Green Version]
- Klaunig, J.E.; Zemin, W.; Xinzhu, P.; Shaoyu, Z. Oxidative stress and oxidative damage in chemical carcinogenesis and multistage carcinogenesis. Toxicol. Appl. Pharmacol. 2011, 254, 86–99. [Google Scholar] [CrossRef]
- Reichert, S. Determinants of Telomere Length and Implications in Life History Trade-Offs. Ph.D. Thesis, University De Strasbourg, Strasbourg, France, 2013; p. 344. [Google Scholar]
- Kim, Y.J.; Subramani, V.K.; Sohn, S.H. Age prediction in the chickens using telomere quantity by quantitative fluorescence in situ hybridisation technique. Asian Aust. J. Anim. Sci. 2011, 24, 603–609. [Google Scholar] [CrossRef]
- Harley, C.B.; Futcher, A.B.; Greider, C.W. Telomeres shorten during ageing of human fibroblasts. Nature 1990, 345, 458–460. [Google Scholar] [CrossRef] [PubMed]
- Martin, T.E.; Riordan, M.M.; Repin, R.; Mouton, J.C.; Blake, W.M.; Fleishman, E. Apparent annual survival estimates of tropical songbirds better reflect life history variation when based on intensive field methods. Glob. Ecol. Biogeogr. 2017, 26, 1386–1397. [Google Scholar] [CrossRef]
- Muñoz, A.P.; Kéry, M.; Martins, P.V.; Ferraz, G. Age effects on survival of Amazon forest birds and the latitudinal gradient in bird survival. Auk Ornithol. Adv. 2018, 135, 299–313. [Google Scholar] [CrossRef] [Green Version]
- Wilbourn, R.V.; Moatt, J.P.; Froy, H.; Wally, C.A.; Nussey, D.H.; Broonekamp, J.J. The relationship between telomere length and mortality Risk in non-model vertebrate system: A Meta Analysis. Phil. Trans. R. Soc. B Biol. Sci. 2018, 373, 20160447. [Google Scholar] [CrossRef] [PubMed]
- Tricola, G.M.; Simons, M.J.P.; Atema, E.; Boughton, R.K.; Brown, J.L.; Dearborn, D.C.; Divoky, G.; Eimes, J.A.; Huntington, C.E.; Kitaysky, A.E.; et al. The rate of telomere loss is related to maximum lifespan in birds. Phil. Trans. R. Soc. B Biol. Sci. 2018, 373, 1741. [Google Scholar] [CrossRef] [PubMed]
- Blackburn, E.H. Switching and signalling at the telomere. Cell 2001, 106, 661–673. [Google Scholar] [CrossRef] [Green Version]
- Dunham, M.A.; Neumann, A.A.; Fasching, C.L.; Reddel, R.R. Telomere maintenance by recombination in human cells. Nat. Genet. 2000, 26, 447–450. [Google Scholar] [CrossRef]
- Greider, C.W. Telomerase activity, cell proliferation, and cancer. Proc. Natl. Acad. Sci. USA 1998, 95, 90–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez, P.; Blasco, M.A. Heart-Breaking telomeres. Circ. Res. 2018, 123, 787–802. [Google Scholar] [CrossRef] [PubMed]
- Olkowski, A.A. Pathophysiology of heart failure in broiler chickens: Structural, biochemical and molecular characteristics. Poult. Sci. 2007, 85, 999–1005. [Google Scholar] [CrossRef] [PubMed]
- Strong, M.A.; Vidal-Cadenas, S.L.; Karim, B.; Yu, H.; Guo, N.; Greider, C.W. Phenotypes in mTERT+/− and mTERT-/- mice are due to short telomeres, not telomere-independent functions of telomerase reverse transcriptase. Mol. Cell Biol. 2011, 31, 2369–2379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armanios, M. Telomeres and age-related disease: How telomere biology informs clinical paradigm. J. Clin. Investig. 2013, 123, 996–1002. [Google Scholar] [CrossRef] [Green Version]
- Booth, S.A.; Charchar, F.J. Cardiac telomere length in heart development, function and disease. Physiol. Genom. 2017, 49, 368–384. [Google Scholar] [CrossRef] [PubMed]
- Yip, B.W.; Mok, H.O.; Peterson, D.R.; Wan, M.T.; Taniguchi, Y.; Ge, W.; Au, D.W. Sex-dependent telomere shortening, telomerase activity and oxidative damage in marine medaka Oryzias melastigma during aging. Mar. Pollut. Bull. 2017, 124, 701–709. [Google Scholar] [CrossRef] [PubMed]
- Epel, E.S.; Blackburn, E.H.; Lin, J.; Dhabhar, F.S.; Adler, N.E.; Morrow, J.D.; Cawthon, R.M. Accelerated telomere shortening in response to life stress. Proc. Natl. Acad. Sci. USA 2004, 101, 17312–17315. [Google Scholar] [CrossRef] [Green Version]
- Monaghan, P.; Haussmann, M.F. Do telomere dynamics link lifestyle and lifespan? Trends Ecol. Evol. 2006, 21, 47–53. [Google Scholar] [CrossRef]
- Monaghan, P. Telomeres and life histories: The long and the short of it. Ann. N. Y. Acad. Sci. 2010, 1206, 130–142. [Google Scholar] [CrossRef] [PubMed]
- Bauch, C.; Becker, P.H.; Verhulst, S. Telomere length reflects phenotypic quality and costs of reproduction in a long-lived seabird. Proc. R. Soc. Lond. 2013, 280, 20122540. [Google Scholar] [CrossRef] [Green Version]
- Yip, S.H.; Sham, P.C.; Wang, J. Evaluation of tools for highly variable gene discovery from single-cell RNA-seq data evaluation of tools for highly variable gene discovery from single-cell RNA-seq data. Brief. Bioinform. 2018, 20, 1583–1589. [Google Scholar] [CrossRef] [PubMed]
- Cusanelli, E.; Angelica, C.; Romero, P.; Chartrand, P. Telomeric noncoding RNA TERRA is induced by telomere shortening to nucleate telomerase molecules at short telomeres. Mol. Cell 2013, 51, 780–791. [Google Scholar] [CrossRef] [Green Version]
- Okamoto, K.; Iwano, T.; Tachibana, M.; Shinkai, Y. Distinct roles of TRF1 in the regulation of telomere structure and lengthening. J. Biol. Chem. 2008, 283, 23981–23988. [Google Scholar] [CrossRef] [Green Version]
- Kolquist, K.A.; Ellisen, L.W.; Counter, C.M.; Meyerson, M.; Tan, L.K.; Weinberg, R.A.; Haber, D.A.; Gerald, W.L. Expression of TERT in early premalignant lesions and a subset cells in normal tissues. Nat. Genet. 1998, 19, 182–186. [Google Scholar] [CrossRef]
- Ludlow, A.T.; Spangenburg, E.E.; Chin, E.R.; Cheng, W.H.; Roth, S.M. Telomeres shorten in response to oxidative stress in mouse skeletal muscle fibers. J. Gerontol. Biol. Sci. Med. Sci. J. 2014, 69, 821–830. [Google Scholar] [CrossRef] [Green Version]
- Schmutz, I.; de Lange, T. Shelterin. Curr. Biol. 2016, 26, R397–R399. [Google Scholar] [CrossRef]
- Bejarano, L.; Schuhmacher, A.J.; Squatrito, M.; Blasco, M.A. Inhibition of TRF1 telomere protein impairs tumor initiation and progression in glioblastoma mouse models and patient-derived xenografts. Cancer Cell 2017, 32, 590–607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- RGD (Rat Genome Database). (N.D). Telomere Maintenance 2 Gene. 2020. Available online: https://www.rgd.mcw.edu (accessed on 3 June 2019).
- Scheibe, M.; Arnoult, N.; Kappei, D.; Bucholz, F.; Decottignies, A.; Butter, F.; Mann, M. Quantitative interaction screen of telomeric repeat-containing RNA reveal novel TERRA regulators. Genome Res. 2013, 23, 2149–2157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maicher, A.; Kastner, L.; Dees, M.; Luke, B. Deregulated telomere transcription causes replication—Dependent telomere shortening and promotes cellular senescence. Nucleic Acid Res. 2012, 40, 6649–6659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prowse, K.R.; Greider, C.W. Development and tissue specific regulation of mouse telomerase and telomere length. Proc. Natl. Acad. Sci. USA 1995, 92, 4818–4822. [Google Scholar] [CrossRef] [Green Version]
- Shay, J.W.; Bacchetti, S. A survey of telomerase activity in human cancer. Eur. J. Cancer 1997, 33, 787–791. [Google Scholar] [CrossRef]
- Goichberg, P.; Chang, J.; Liao, R.; Leri, A. Cardiac stem cells: Biology and clinical applications. Antioxid. Redox Signal. 2014, 21, 2002–2017. [Google Scholar] [CrossRef] [PubMed]
- Bettin, N.; Pegorar, C.O.; Cusanelli, E. The emerging roles of TERRA in telomere maintenance and genome stability. Cells 2019, 8, 246. [Google Scholar] [CrossRef] [Green Version]
- Yifan, C.; Tianzheng, Y. Mouse liver is more resistant than skeletal muscle to heat-induced apoptosis. Cell Stress Chaperones 2021, 26, 275–281. [Google Scholar] [CrossRef]
- Lu, A.L.; Li, X.; Gu, Y.; Wright, P.M.; Chang, D.Y. Repair of oxidative DNA damage:mechanisms and functions. Cell Biochem. Biophys. 2001, 35, 141–170. [Google Scholar] [CrossRef]
- Marnett, L.J. Oxyradicals and DNA damage. Carcinogenesis 2000, 21, 361–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Composition | Starter 1 | Finisher 2 |
|---|---|---|
| Crude protein (%) | 23.00 | 19.00 |
| Crude fiber (%) | 5.00 | 5.00 |
| Crude fat (%) | 5.00 | 5.00 |
| Moisture (%) | 13.00 | 13.00 |
| Ash (%) | 8.00 | 8.00 |
| Calcium (%) | 0.80 | 0.80 |
| Phosphorous (%) | 0.40 | 0.40 |
| No | Gene | Primers’ Sequence | Accession Numbers | Amplicon Sizes (bp) |
|---|---|---|---|---|
| 1 | Telomere | F—GGTTTTTGAGGGTGAGGGTGAGGGTGAGGGTGAGGGT R—TCCCGACTATCCCTATCCCTATCCCTATCCCTATCCCTA | NA | 79 |
| 2 | TRF1 | F—GGAGGAACGGTTTCCCTAAG R—CTGATGCTGCCCACAGTAGA | NC-006089.5 | 178 |
| 3 | TERRA | F—GGCCACTGTAAATGGCTGTT R—GTTTGCACAAGGGTCTCCAT | NC-006127.5 | 219 |
| 4 | HSF1 | F—TCTCTGGGTGTCCTTCTGCT R—CTCCTTCCACAGAGCACCTC | NC-006089.5 | 151 |
| 5 | TELO2 | F—GGATGACCCTCAGAGATGGA R—ATTGGTGTGACCAGGAAAGCe | NC-006101.5 | 249 |
| 6 | chTERT | F—AGGTGCCCAAAACTGAACAC R—CTTCCAAGGGAGACTTGCAG | NC-006089.5 | 184 |
| 7 | GAPDH | F—ACTATGCGGTTCCCAGTGTC R—TGCCACCATCAGAAAAATGA | NC-006088.5 | 215 |
| Age/Traits | CTRL | CORT-Fed Chicken | SEM | p-Values |
|---|---|---|---|---|
| Body weight (g) | ||||
| Week 2 | 541.88 | 539.76 | 8.76 | 0.8585 |
| Week 4 | 1509.00 | 1054.21 | 23.48 | 0.001 |
| Week 6 | 2363.55 | 1479.18 | 34.62 | 0.001 |
| Feed consumption (g/bird/week) | ||||
| Week 2 | 422.00 | 439.80 | 8.22 | 0.1730 |
| Week 4 | 990.00 | 843.30 | 17.65 | 0.0004 |
| Week 6 | 1057.70 | 822.30 | 22.57 | 0.0001 |
| Weight gain (g/bird/week) | ||||
| Week 2 | 347.56 | 344.83 | 5.41 | 0.735 |
| Week 4 | 496.42 | 259.55 | 17.39 | 0.0001 |
| Week 6 | 1056.67 | 822.33 | 22.57 | 0.0001 |
| FCR (feed/gain) | ||||
| Week 2 | 1.21 | 1.27 | 0.02 | 0.0557 |
| Week 4 | 2.00 | 3.27 | 0.12 | 0.0001 |
| Week 6 | 2.27 | 3.60 | 0.11 | 0.0001 |
| Mortality (Week 0–Week 6) | 1.00 | 7.00 | 0.09 | 0.024 |
| Age | Treatment | Heart (g) | Liver (g) | Smallint (g) | AbdFat (g) | Gizzard (g) |
|---|---|---|---|---|---|---|
| Week 4 | CTRL | 7.85 b | 36.27 c | 76.55 c | 17.87 d | 29.36 b |
| CORT | 7.22 b | 55.21 b | 66.09 c | 27.24 c | 36.13 ab | |
| Week 6 | CTRL | 11.24 a | 49.17 b | 123.56 a | 52.03 b | 29.36 b |
| CORT | 11.74 a | 75.02 a | 93.63 b | 63.12 a | 42.42 a | |
| SEM | 0.54 | 3.42 | 2.48 | 2.48 | 2.95 | |
| p values | Age | <0.0001 | <0.0001 | <0.0001 | <0.0001 | 0.359 |
| Treatment | 0.907 | <0.0001 | <0.0001 | <0.0001 | 0.004 | |
| Age × Treatment | 0.319 | 0.363 | 0.041 | 0.74 | 0.287 |
| Age | Treatment | Heart (%) | Liver (%) | Smallint (%) | AbdFat (%) | Gizzard (%) |
|---|---|---|---|---|---|---|
| Week 4 | CTRL | 0.50 b | 2.31 b | 4.88 b | 1.14 d | 1.90 c |
| CORT | 0.68 a | 5.15 a | 6.21 a | 2.55 b | 3.35 a | |
| Week 6 | CTRL | 0.44 b | 1.93 b | 4.87 b | 2.06 c | 1.17 d |
| CORT | 0.74 a | 4.59 a | 5.72 a | 3.83 a | 2.61 b | |
| SEM | 0.03 | 0.20 | 0.22 | 0.14 | 0.20 | |
| p values | Age | 0.756 | 0.041 | 0.268 | <0.0001 | 0.0016 |
| Treatment | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | |
| Age × Treatment | 0.212 | 0.667 | 0.282 | 0.213 | 0.999 |
| Age | Week 4 | Week 6 | p Values | |||||
|---|---|---|---|---|---|---|---|---|
| Treatment | CTRL | CORT | CTRL | CORT | SEM | Age | Treatment | Age × Treatment |
| Plasma CORT (ng/mL) | 6.80 b | 7.65 a | 6.73 b | 7.94 a | 0.37 | 0.685 | 0.0004 | 0.830 |
| Age | Treatment | Whole Blood | Muscle | Liver | Heart |
|---|---|---|---|---|---|
| Week 4 | CTRL | 526.40 a | 446.68 a | 564.96 a | 576.89 a |
| CORT | 334.50 bc | 313.84 b | 346.84 b | 264.43 b | |
| Week 6 | CTRL | 417.96 ab | 463.41 a | 481.07 ab | 355.72 ab |
| CORT | 260.09 c | 282.39 b | 467.59 ab | 364.98 ab | |
| SEM | 37.27 | 54.90 | 58.33 | 56.03 | |
| p values | Age | 0.016 | 0.779 | 0.807 | 0.316 |
| Treatment | <0.0001 | 0.005 | 0.028 | 0.009 | |
| Age × Treatment | 0.921 | 0.817 | 0.047 | 0.018 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Badmus, K.A.; Idrus, Z.; Meng, G.Y.; Sazili, A.Q.; Mamat-Hamidi, K. Telomere Length and Regulatory Genes as Novel Stress Biomarkers and Their Diversities in Broiler Chickens (Gallus gallus domesticus) Subjected to Corticosterone Feeding. Animals 2021, 11, 2759. https://doi.org/10.3390/ani11102759
Badmus KA, Idrus Z, Meng GY, Sazili AQ, Mamat-Hamidi K. Telomere Length and Regulatory Genes as Novel Stress Biomarkers and Their Diversities in Broiler Chickens (Gallus gallus domesticus) Subjected to Corticosterone Feeding. Animals. 2021; 11(10):2759. https://doi.org/10.3390/ani11102759
Chicago/Turabian StyleBadmus, Kazeem Ajasa, Zulkifli Idrus, Goh Yong Meng, Awis Qurni Sazili, and Kamalludin Mamat-Hamidi. 2021. "Telomere Length and Regulatory Genes as Novel Stress Biomarkers and Their Diversities in Broiler Chickens (Gallus gallus domesticus) Subjected to Corticosterone Feeding" Animals 11, no. 10: 2759. https://doi.org/10.3390/ani11102759