Dietary Black Seed Effects on Growth Performance, Proximate Composition, Antioxidant and Histo-Biochemical Parameters of a Culturable Fish, Rohu (Labeo rohita)

 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Fish Culture and Diets Preparation
2.2. Water Quality Parameters
2.3. Fish Sampling
2.4. Preparation of Tissue Homogenates
2.5. Growth Performance
2.6. Proximate Composition Analysis
2.7. Oxidative Stress and Antioxidant Defense Markers Assessment
2.8. Liver and Kidney Functioning Tests
2.9. Histological Preparations
2.10. Data Analysis
2.11. Ethical Statement
3. Results
3.1. Growth Performance
3.2. Proximate Composition of Black Seed, Basal Diet and Dorsal Muscles of Rohu
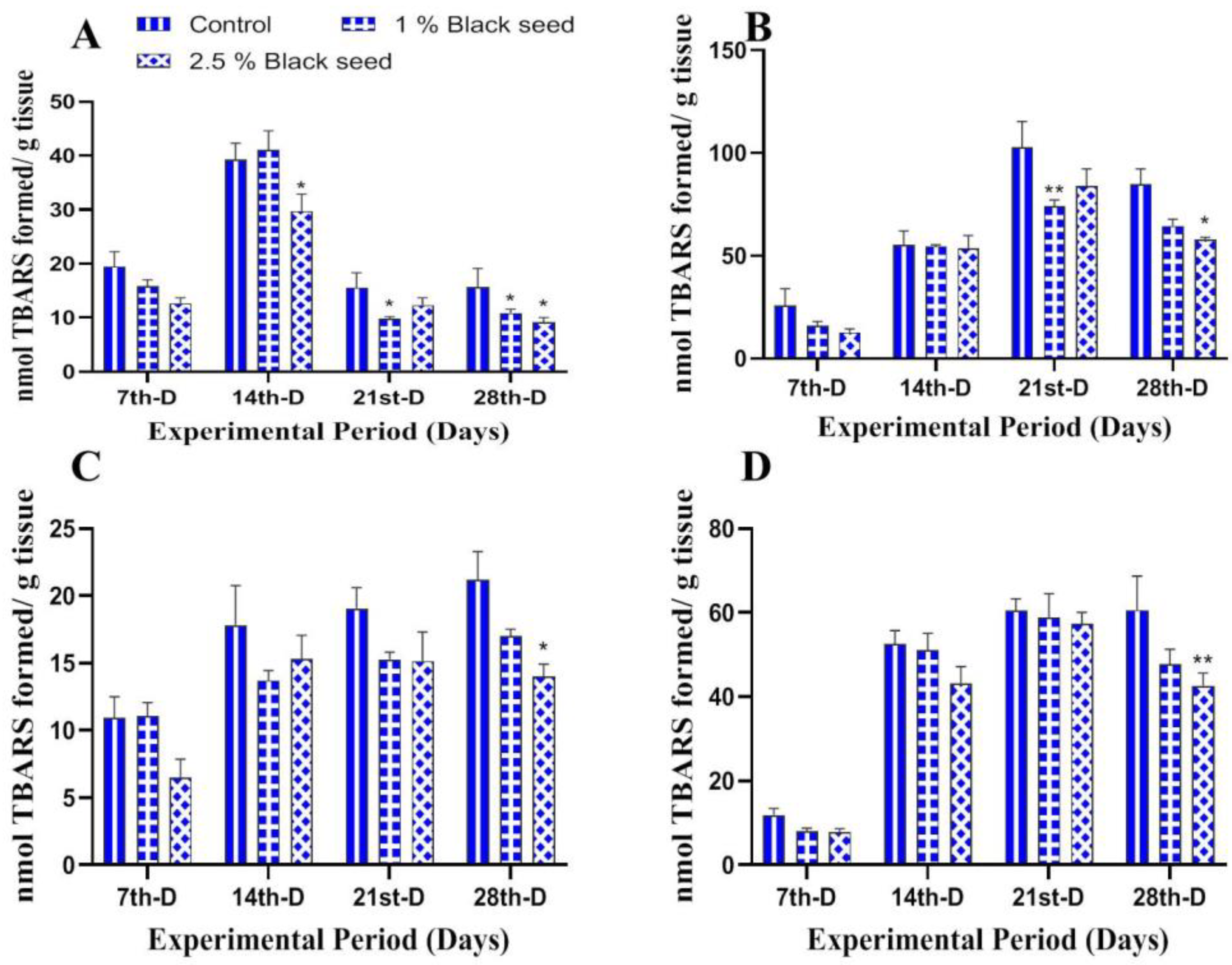
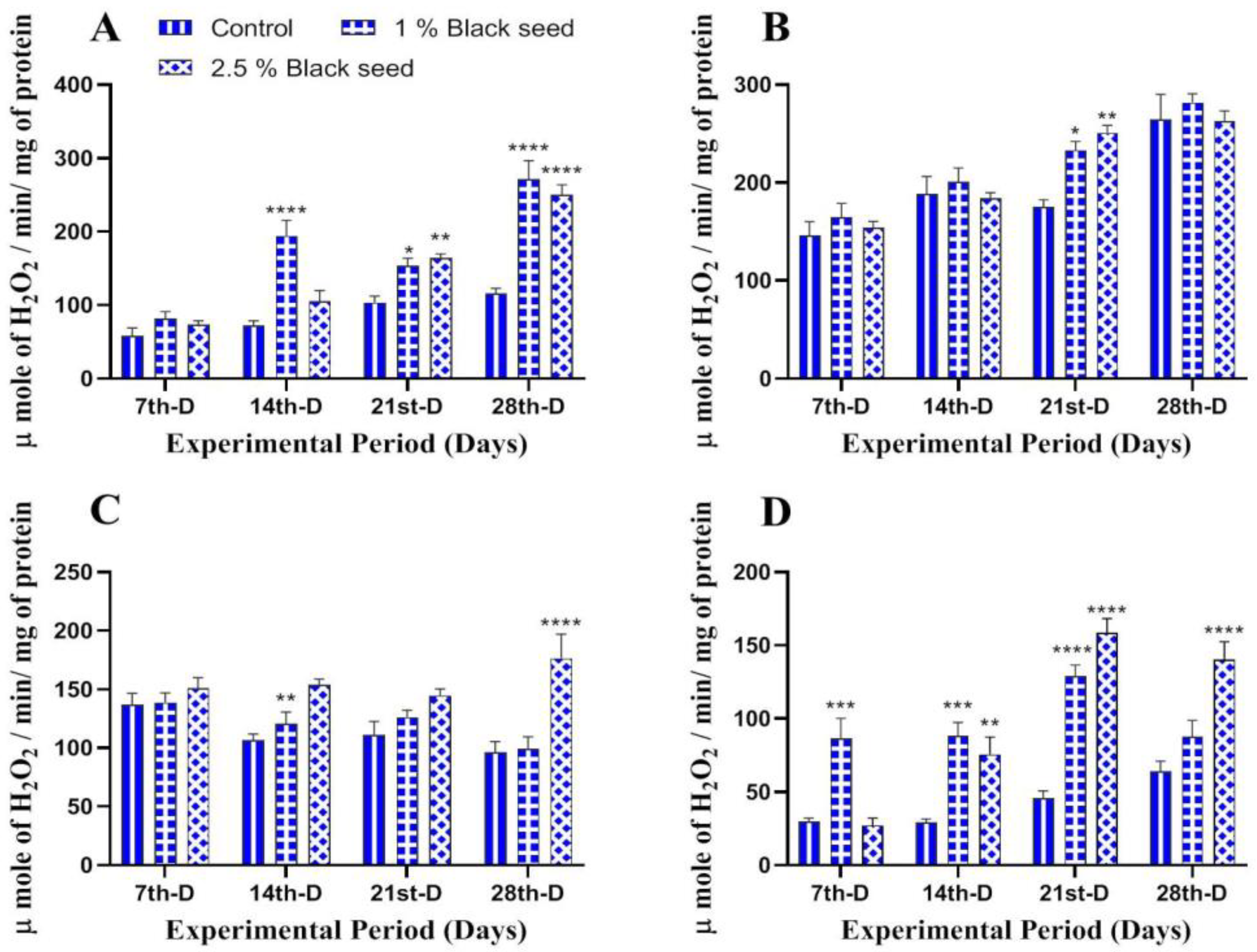
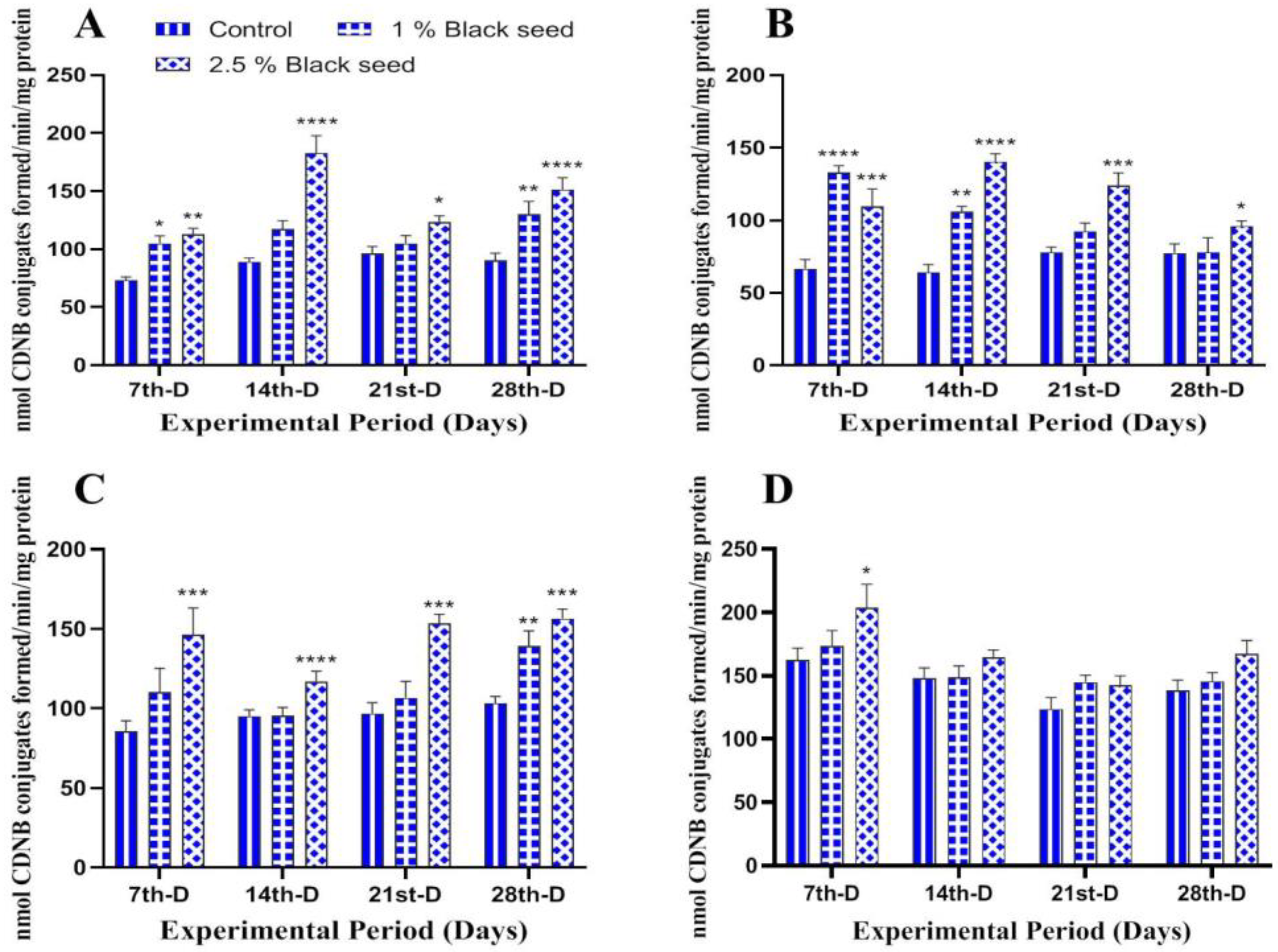
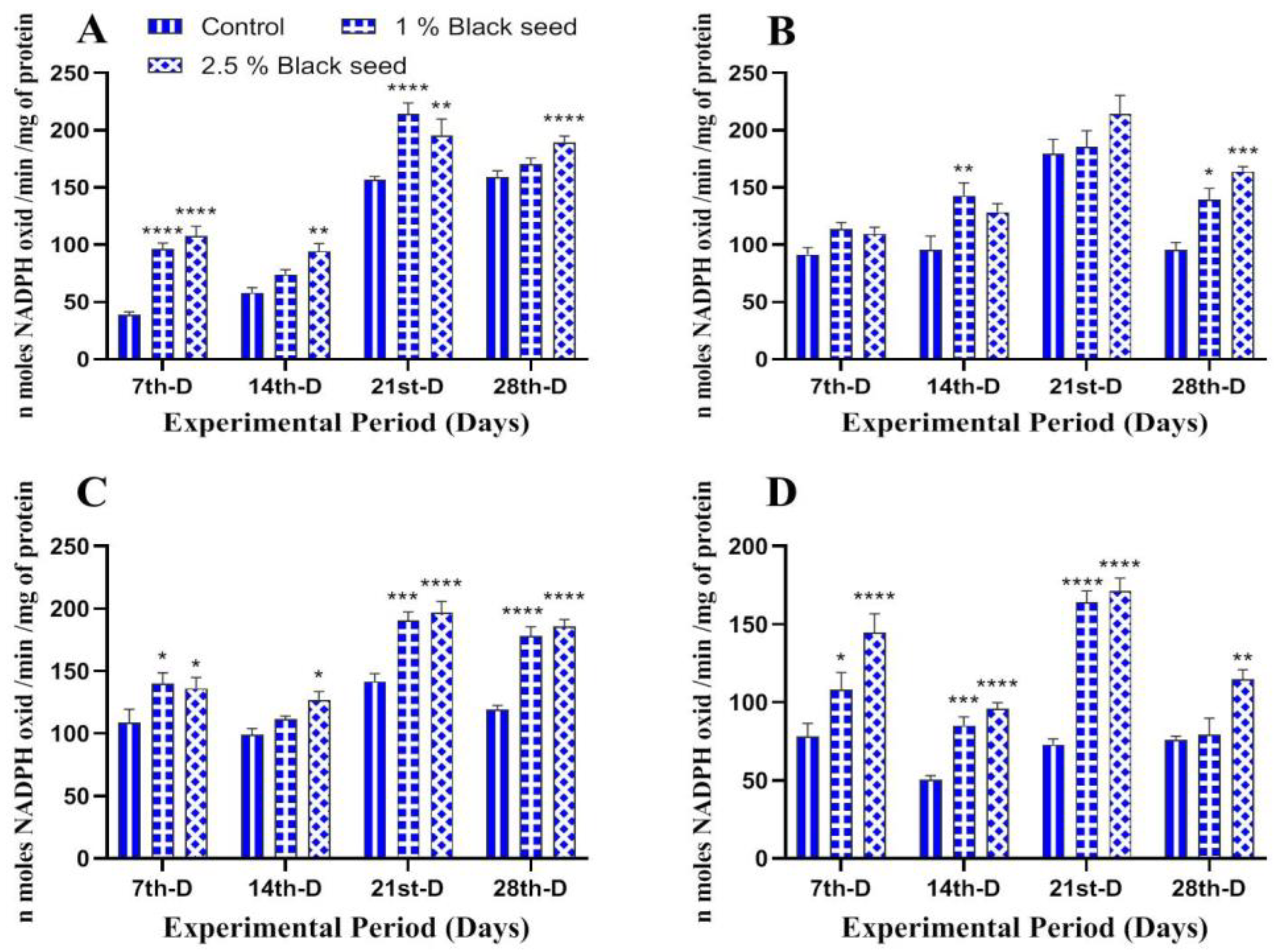
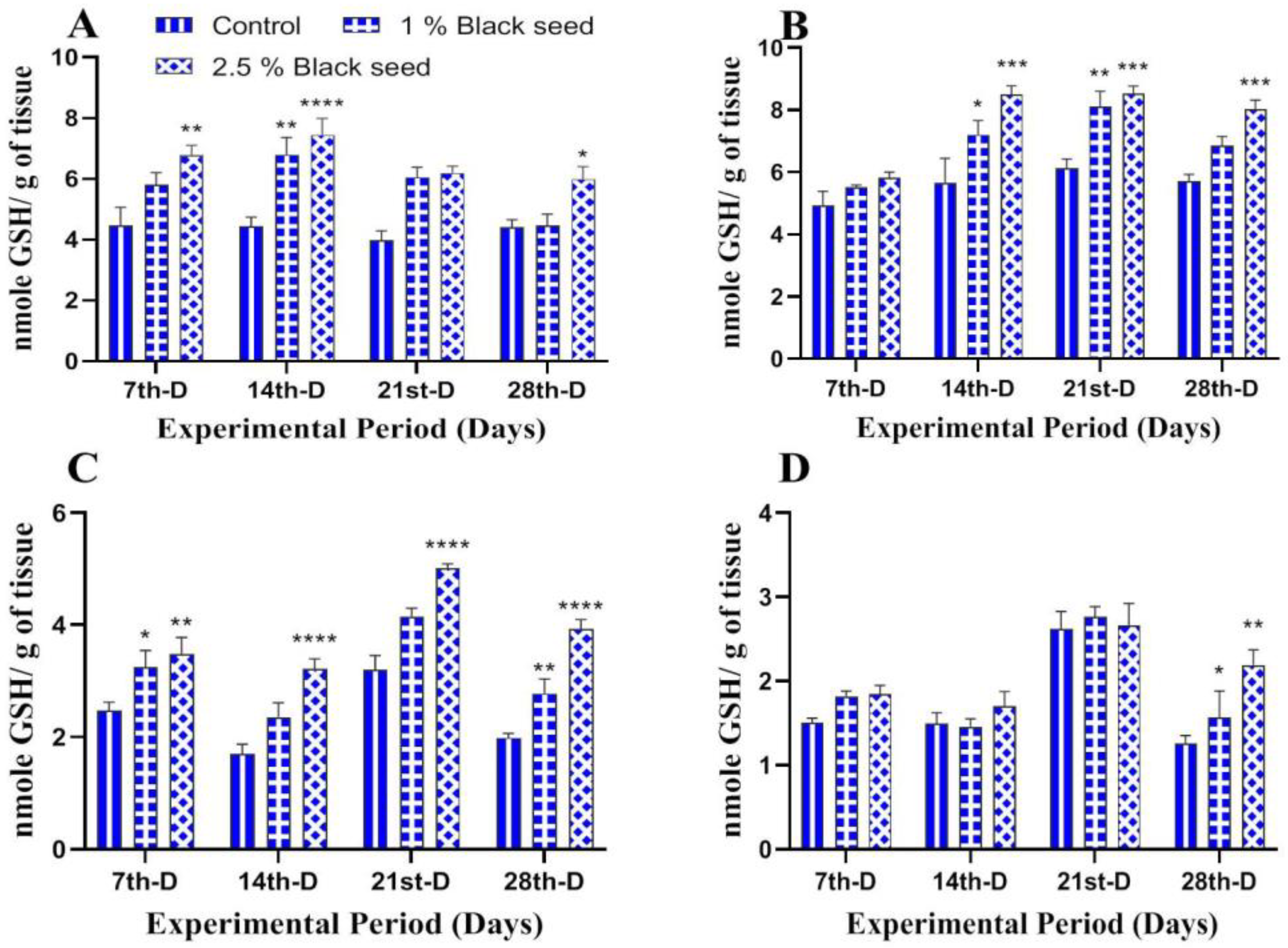
3.3. Antioxidant Status
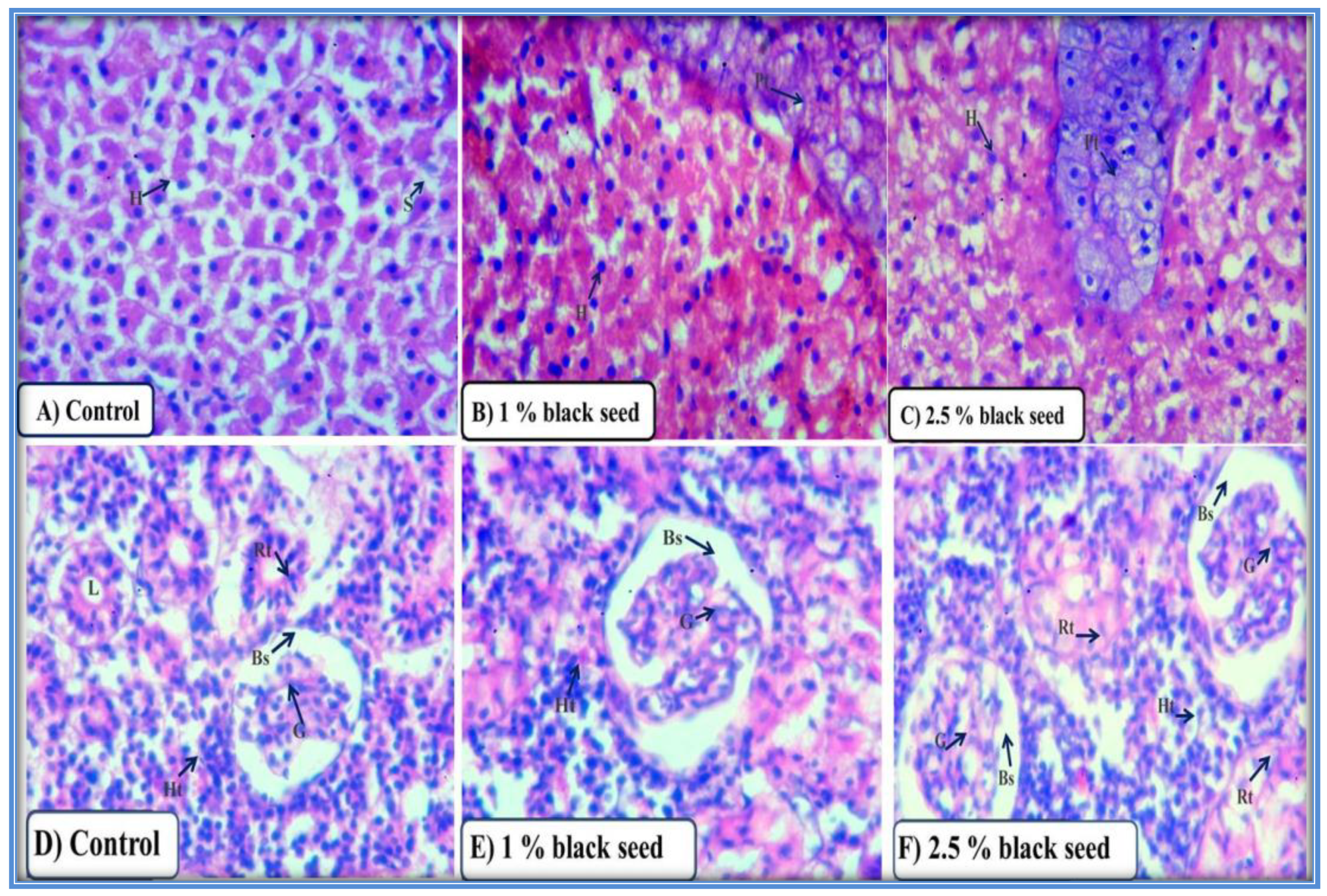
3.4. Liver and Kidney Histo-Biochemical Parameters
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Little, D.C.; Newton, R.W.; Beveridge, M.C.M. Aquaculture: A rapidly growing and significant source of sustainable food? Status, transitions and potential. Proc. Nutr. Soc. 2016, 75, 274–286. [Google Scholar] [CrossRef] [PubMed]
- Martinez, R.; Carpio, Y.; Morales, A.; Lugo, J.M.; Herrera, F.; Zaldivar, C.; Carrillo, O.; Arenal, A.; Pimentel, E.; Estrada, M.P. Oral administration of the growth hormone secretagogue-6 (GHRP-6) enhances growth and non-specific immune responses in tilapia (Oreochromis sp.). Aquaculture 2016, 452, 304–310. [Google Scholar] [CrossRef]
- Chuah, L.O.; Effarizah, M.E.; Goni, A.M.; Rusul, G. Antibiotic application and emergence of multiple antibiotic resistance (MAR) in global catfish aquaculture. Curr. Environ. Health. Reports. 2016, 3, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Bahadir, T.; Celebi, H.; Şimsek, I.; Tulun, S. Antibiotic applications in fish farms and environmental problems. Turkish J. Eng. 2019, 3, 60–67. [Google Scholar] [CrossRef] [Green Version]
- Dawood, M.A.O.; Koshio, S.; Esteban, M.A. Beneficial roles of feed additives as immunostimulants in aquaculture: A review. Rev. Aquac. 2018, 10, 950–974. [Google Scholar] [CrossRef]
- Awad, E.; Awaad, A. Role of medicinal plants on growth performance and immune status in fish. Fish Shellfis Immunol. 2017, 67, 40–54. [Google Scholar] [CrossRef] [PubMed]
- Bulfon, C.; Volpatti, D.; Galeotti, M. Current research on the use of plant-derived products in farmed fish. Aquac. Res. 2015, 46, 513–551. [Google Scholar] [CrossRef]
- Lamin, B.D.; Kumar, M.; Pailan, G.H.; Singh, D.K.; Bisawal, A. Effect of dietary supplementation of Zingiber officinale (ginger) on growth and nutrient utilization of Cyprinus carpio (Linnaeus, 1758). J. Exp. Zool. 2018, 21, 849–853. [Google Scholar]
- Kapinga, I.B.; Limbu, S.M.; Madalla, N.A.; Kimaro, W.H.; Mabiki, F.P.; Lamtane, H.A.; Tamatamah, R.A. Dietary Aspilia mossambicensis and Azadirachta indica supplementation alter gonadal characteristics and histology of juvenile Nile tilapia (Oreochromis niloticus). Aquac. Res. 2019, 50, 573–580. [Google Scholar] [CrossRef]
- Faheem, M.; Khaliq, S.; Mustafa, N.; Rani, S.; Lone, K.P. Dietary Moringa oleferia leaf meal induce growth, innate immunity and cytokine expression in grass carp, Ctenopharyngodon idella. Aquac. Nutr. 2020, 26, 1164–1172. [Google Scholar] [CrossRef]
- Yilmaz, E.; Tasbozan, O.; Erbas, C. Potential of medical herbal products to be used in aquaculture. Int. J. Sci. Eng. Res. 2018, 9, 16–23. [Google Scholar]
- Botnick, I.; Xue, W.; Bar, E.; Ibdah, M.; Schwartz, A.; Joel, D.M.; Lev, E.; Fait, A.; Lewinsohn, E. Distribution of primary and specialized metabolites in Nigella sativa seeds, a spice with vast traditional and historical uses. Molecules 2012, 17, 10159–10177. [Google Scholar] [CrossRef] [PubMed]
- Polat, R.; Satıl, F.; Cakıloglu, U. Medicinal plants and their use properties of sold in herbal market in Bingol (Turkey) district. Biol. Divers. Conserv. 2011, 4, 25–35. [Google Scholar]
- Tembhurne, S.V.; Feroz, S.; More, B.H.; Sakarkar, D.M. A review on therapeutic potential of Nigella sativa (kalonji) seeds. J. Med. Plants. Res. 2014, 8, 167–177. [Google Scholar]
- Ahmad, A.; Husain, A.; Mujeeb, M.; Khan, S.A.; Najmi, A.K.; Siddique, N.A.; Damanhouri, Z.A.; Anwar, F. A review on therapeutic potential of Nigella sativa: A miracle herb. Asian Pac. J. Trop. Biomed. 2013, 3, 337–352. [Google Scholar] [CrossRef] [Green Version]
- Hussein, M.M.A.; El-Feki, M.A.; Hatai, K.; Yamamoto, A. Inhibitory effects of thymoquinone from Nigella sativa on pathogenic Saprolegnia in fish. Biocontrol. Sci. 2002, 7, 31–35. [Google Scholar] [CrossRef]
- Mohammed, H.H.; Arias, C.R. Protective efficacy of Nigella sativa seeds and oil against columnaris disease in fishes. J. Fish. Dis. 2016, 39, 693–703. [Google Scholar] [CrossRef]
- Pes, T.S.; Saccol, E.M.H.; Ourique, G.M.; Londero, E.P.; Gressler, L.T.; Golombieski, J.I.; Glanzner, W.G.; Llesuy, S.F.; Gonçalves, P.B.D.; Neto, J.R.; et al. Quercetin in the diet of silver catfish: Effects on antioxidant status, blood parameters and pituitary hormone expression. Aquaculture 2016, 458, 100–106. [Google Scholar] [CrossRef]
- Altunoglu, C.Y.; Bilen, S.; Ulu, F.; Biswas, G. Immune responses to methanolic extract of black cumin (Nigella sativa) in rainbow trout (Oncorhynchus mykiss). Fish Shellfish Immunol. 2017, 67, 103–109. [Google Scholar] [CrossRef]
- Saad, T.T.; Abou El-Geit, E.N.; El-Hammady, A.K.I. Effect of black cumin seeds (Nigella sativa) and/or turmeric (curcumin) on hematological, biochemical and immunological parameters of sea bass vaccinated with Pseudomonas fluorescence Bacterin. Life Sci. J. 2013, 10, 1292–1303. [Google Scholar]
- Kotb, A.M.; Abd-Elkareem, M.; Abou Khalil, N.S.; Sayed, A.E.D.H. Protective effect of Nigella sativa on 4-nonylphenol-induced nephrotoxicity in Clarias gariepinus (Burchell, 1822). Sci. Total. Environ. 2018, 619, 692–699. [Google Scholar] [CrossRef] [PubMed]
- Alishahi, M.; Mesbah, M. Effects of Viscum album and Nigella sativa extracts on survival rate, growth factors and resistance to Aeromonas hydrophila infection in gold fish (Carassius auratus). J. Vet. Res. 2012, 67, 285–290. [Google Scholar]
- Elkamel, A.A.; Mosaad, G.M. Immunomodulation of Nile tilapia, Oreochromis niloticus, by Nigella sativa and Bacillus subtilis. J. Aquac. Res. Dev. 2012, 3, 1–4. [Google Scholar] [CrossRef]
- Abdelwahab, A.M.; El-Bahr, S.M. Influence of black cumin seeds (Nigella sativa) and turmeric (Curcuma longa Linn.) mixture on performance and serum biochemistry of Asian sea bass, Lates calcarifer. World J. Fish Mar. Sci. 2012, 4, 496–503. [Google Scholar] [CrossRef]
- Awad, E.; Austin, D.; Lyndon, A.R. Effect of black cumin seed oil (Nigella sativa) and nettle extract (Quercetin) on enhancement of immunity in rainbow trout, Oncorhynchus mykiss (Walbaum). Aquaculture 2013, 388, 193–197. [Google Scholar] [CrossRef]
- Awad, E.; Awaad, A.S.; Esteban, M.A. Effects of dihydroquercetin obtained from deodar (Cedrus deodara) on immune status of gilthead seabream (Sparus aurata L.). Fish Shellfish Immunol. 2015, 43, 43–50. [Google Scholar] [CrossRef] [Green Version]
- Khatun, A.; Yasmin, F.; Hossain, M.; Alam, M.; Rahman, M.; Islam, M. Effect of black cumin seed oil (Nigella sativa) on enhancement of immunity in the climbing perch, Anabas testudineus. Br. Microbiol. Res. J 2015, 6, 331–339. [Google Scholar] [CrossRef]
- Khondoker, S.; Hossain, M.M.M.; Hasan-UJ-Jaman, M.; Alam, M.E.; Zaman, M.F.U.; Tabassum, N. Effect of Nigella sativa (black cumin seed) to enhance the immunity of common carp (Cyprinus carpio) against Pseudomonas fluorescens. Am. J. Life Sci 2016, 4, 87–92. [Google Scholar] [CrossRef] [Green Version]
- Latif, M.; Ullah, M.Z.; Rasheed, A.; Hussain, M.; Naqvi, H.N.; Latif, S. Effect of varying dietary protein levels on the growth response of Labeo rohita (Rohu) fingerlings under semi-intensive culture system. Int. J. Biosci. 2018, 12, 82–87. [Google Scholar] [CrossRef]
- Agha, Q.; Ahmad, S.; Islam, M.; Gill, A.; Athar, M. Growth and production potential of three exotic herbs in highlands of Balochistan, Pakistan. J. Med. Plants Res. 2011, 5, 543–548. [Google Scholar]
- Rabbani, M.A.; Ghafoor, A.; Masood, M.S. NARC-Kalonji: An early maturing and high yielding variety of Nigella sativa released for cultivation in Pakistan. Pak. J. Bot. 2011, 43, 191–195. [Google Scholar]
- Sher, H.; Barkworth, M.E. Economic development through medicinal and aromatic plants (MAPs) cultivation in Hindu Kush Himalaya mountains of District Swat, Pakistan. J. Mt. Sci. 2015, 12, 1292–1301. [Google Scholar] [CrossRef]
- Bektas, Z.H.; Savaşer, S.; Akçimen, U.; Ceylan, M.; Yener, O.; Bulut, C. Using of black cumin seed powder (Nigella sativa) as immunostimulant and growth promoter in rainbow trout, Oncorhynchus mykiss (Walbaum). Turkish J. Fish Aquat. Sci. 2018, 19, 987–999. [Google Scholar]
- Latif, M.; Faheem, M.; Asmatullah; Hoseinifar, S.H.; Doan, H.V. Protective efficacy of Nigella sativa seeds against diethyl phthalate induced growth retardation, oxidative stress and histo-biochemical damages in Labeo rohita. Aquaculture 2020, 736065. [Google Scholar] [CrossRef]
- Faheem, M.; Lone, K.P. Oxidative stress and histopathologic biomarkers of exposure to bisphenol-A in the freshwater fish, Ctenopharyngodon idella. Brazilian J. Pharm. Sci. 2017, 53, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Latif, M.; Faheem, M.; Asmatullah. Study of oxidative stress and histo-biochemical biomarkers of diethyl phthalate induced toxicity in a cultureable fish, Labeo rohita. Pak. Vet. J. 2020, 40, 202–208. [Google Scholar] [CrossRef]
- Zaki, M.A.; Labib, E.M.; Nour, A.M.; Tonsy, H.D.; Mahmoud, S.H. Effect some medicinal plants diets on mono sex Nile tilapia (Oreochromis niloticus), growth performance, feed utilization and physiological parameters. APCBEE Procedia 2012, 4, 220–227. [Google Scholar] [CrossRef] [Green Version]
- Yilmaz, S.; Ergun, S.; Celik, E.S. Effects of herbal supplements on growth performance of sea bass (Dicentrarchus labrax): Change in body composition and some blood parameters. J. Biosci. Biotechnol. 2012, 1, 217–222. [Google Scholar]
- Gabriel, N.N.; Qiang, J.; He, J.; Ma, Y.X.; Kpundeh, M.D.; Xu, P. Dietary Aloe vera supplementation on growth performance, some haemato-biochemical parameters and disease resistance against Streptococcus iniae in tilapia (GIFT). Fish Shellfish Immunol. 2015, 44, 504–514. [Google Scholar] [CrossRef]
- Abdel-Tawwab, M.; Adeshina, I.; Jenyo-Oni, A.; Ajani, E.K.; Emikpe, B.O. Growth, physiological, antioxidants, and immune response of African catfish, Clarias gariepinus (B.), to dietary clove basil, Ocimum gratissimum, leaf extract and its susceptibility to Listeria monocytogenes infection. Fish Shellfish Immunol. 2018, 78, 346–354. [Google Scholar] [CrossRef]
- Abdel-Tawwab, M.; Samir, F.; Abd El-Naby, A.S.; Monier, M.N. Antioxidative and immunostimulatory effect of dietary cinnamon nanoparticles on the performance of Nile tilapia, Oreochromis niloticus (L.) and its susceptibility to hypoxia stress and Aeromonas hydrophila infection. Fish Shellfish Immunol. 2018, 74, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Guardiola, F.A.; Bahi, A.; Messina, C.M.; Mahdhi, A.; Santulli, A.; Arena, R.; Bakhrouf, A.; Esteban, M.A. Quality and antioxidant response of gilthead seabream (Sparus aurata L.) to dietary supplements of fenugreek (Trigonella foenum graecum) alone or combined with probiotic strains. Fish Shellfish Immunol. 2017, 63, 277–284. [Google Scholar] [CrossRef] [PubMed]
- Diab, A.S.; Aly, S.M.; John, G.; Abde-Hadi, Y.; Mohammed, M.F. Effect of garlic, black seed and Biogen as immunostimulants on the growth and survival of Nile tilapia, Oreochromis niloticus (Teleostei: Cichlidae), and their response to artificial infection with Pseudomonas fluorescens. African J. Aquat. Sci. 2008, 33, 63–68. [Google Scholar] [CrossRef]
- Al-Dubakel, A.; Al-Mhawe, B.; Majeed, M.; Shaeyal, L. Preliminary study on the effect of dietary black seed (Nigella sativa) on growth and blood glucose of common carp (Cyprinus carpio) fingerlings. Ira. J. Thi. Qar. Univ. Agricult. Res. 2012, 1, 41–50. [Google Scholar]
- Oz, M.; Dikel, S.; Durmus, M. Effect of black cumin oil (Nigella sativa) on the growth performance, body composition and fatty acid profile of rainbow trout (Oncorhynchus mykiss). Iran. J. Fish Sci. 2018, 17, 713–724. [Google Scholar] [CrossRef]
- Al-Jassir, M.S. Chemical composition and microflora of black cumin (Nigella sativa L.) seeds growing in Saudi Arabia. Food Chem. 1992, 45, 239–242. [Google Scholar] [CrossRef]
- Ahmed, H.A.; Abusina, G.; Elnady, A.; Abdelrahman, H.; Abedo, A.E. Evaluation of substituting black seed meal (Nigella sativa L.) as protein source in Nile tilapia diets. Biosci. Res. 2018, 15, 1191–1198. [Google Scholar]
- Zaefarian, A.; Yeganeh, S.; Adhami, B. Dietary effects of garlic powder (Allium sativum) on growth, blood indices, carcass composition, and lysozyme activity in brown trout (Salmo caspius) and resistance against Yersinia ruckeri infection. Aquac. Int. 2017, 25, 1987–1996. [Google Scholar] [CrossRef]
- Babayan, V.K.; Koottungal, D.; Halaby, G.A. proximate analysis, fatty acid and amino acid composition of Nigella sativa L. Seeds. J. Food Sci. 1978, 43, 1314–1315. [Google Scholar] [CrossRef]
- Lundebye, A.K.; Hovea, H.; Magea, A.; Bohneb, V.J.B.; Hamrea, K. Levels of synthetic antioxidants (ethoxyquin, butylated hydroxytoluene and butylated hydroxyanisole) in fish feed and commercially farmed fish. Food Addit. Contam. Part A Chem. Anal. Control. Expo. Risk. Assess 2010, 27, 1652–1657. [Google Scholar] [CrossRef]
- Merel, S.; Regueiro, J.; Berntssen, M.H.G.; Hannisdal, R.; Ornsrud, R.; Negreira, N. Identification of ethoxyquin and its transformation products in salmon after controlled dietary exposure via fish feed. Food Chem. 2019, 289, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Habibian, M.; Sadeghi, G.; Karimi, A. Comparative effects of powder, aqueous and methanolic extracts of purslane (Portulaca oleracea L.) on growth performance, antioxidant status, abdominal fat deposition and plasma lipids in broiler chickens. Anim. Prod. Sci. 2019, 59, 89–100. [Google Scholar] [CrossRef]
- Nagi, M.N.; Mansour, M.A. Protective effect of thymoquinone against doxorubicin—Induced cardiotoxicity in rats: A possible mechanism of protection. Pharmacol. Res. 2000, 41, 283–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lesser, M.P. Oxidative stress in marine environments: Biochemistry and physiological ecology. Annu. Rev. Physiol. 2006, 68, 253–278. [Google Scholar] [CrossRef] [Green Version]
- Ming, J.; Ye, J.; Zhang, Y.; Xu, Q.; Yang, X.; Shao, X.; Qiang, J.; Xu, P. Optimal dietary curcumin improved growth performance, and modulated innate immunity, antioxidant capacity and related genes expression of NF-κB and Nrf2 signaling pathways in grass carp (Ctenopharyngodon idella) after infection with Aeromonas hydrophila. Fish Shellfish Immunol. 2020, 97, 540–553. [Google Scholar] [CrossRef]
- Hadi, M.Y.; Mohammed, G.J.; Hameed, I.H. Analysis of bioactive chemical compounds of Nigella sativa using gas chromatography-mass spectrometry. J. Pharmacogn. Phyther. 2016, 8, 8–24. [Google Scholar] [CrossRef]
- Faheem, M.; Khaliq, S.; Lone, K.P. Effect of bisphenol-A on serum biochemistry and liver function in the freshwater fish, Catla catla. Pak. Vet. J. 2019, 39, 71–75. [Google Scholar] [CrossRef]
- Naeiji, N.; Shahsavani, D.; Baghshani, H. Effect of dietary garlic supplementation on lipid peroxidation and protein oxidation biomarkers of tissues as well as some serum biochemical parameters in common carp Cyprinus carpio. Fish Sci. 2013, 79, 699–705. [Google Scholar] [CrossRef]
- Mollazadeh, H.; Hosseinzadeh, H. The protective effect of Nigella sativa against liver injury: A review. Iran. J. Basic Med. Sci 2014, 17, 958–966. [Google Scholar] [CrossRef]
- Ijaz, A.; Ijaz, J.; Aslam, B.; Khan, J.A.; Khaliq, T.; Rahman, Z.; Khan, M.Z.; Naeem, M.A.; Ashraf, M.M. Nephroprotective and antioxidant effects of Moringa oleifera (sohanjna) in paracetamol induced nephrotoxic albino rabbits. Pak. Vet. J. 2016, 36, 292–296. [Google Scholar]
- Gholamnezhad, Z.; Havakhah, S.; Boskabady, M.H. Preclinical and clinical effects of Nigella sativa and its constituent, thymoquinone: A review. J. Ethnopharmacol. 2016, 190, 372–386. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Sampling Period (Days) | Dietary Black Seed Levels | ||
|---|---|---|---|---|
| Group-I (0%) | Group-II (1%) | Group-III (2.5%) | ||
| Initial weight (g/fish) | 0-D | 8.486 ± 0.138 | 8.517 ± 0.355 | 8.506 ± 0.228 |
| Final weight (g/fish) | 7th-D | 8.825 ± 0.136 | 9.693 ± 0.390 | 9.784 ± 0.246 * |
| 14th-D | 9.540 ± 0.178 | 11.32 ± 0.387 *** | 11.44 ± 0.297 *** | |
| 21st-D | 10.11 ± 0.244 | 11.29 ± 0.183 ** | 12.13 ± 0.204 *** | |
| 28th-D | 11.27 ± 0.180 | 11.84 ± 0.282 ** | 12.55 ± 0.206 *** | |
| %WG | 7th-D | 4.056 ± 0.154 | 14.13 ± 1.514 **** | 15.44 ± 1.927 **** |
| 14th-D | 12.58 ± 1.511 | 22.65 ± 1.700 **** | 34.77 ± 1.618 **** | |
| 21st-D | 22.96 ± 1.628 | 34.00 ± 2.012 **** | 36.08 ± 1.626 **** | |
| 28th-D | 32.03 ± 1.648 | 37.96 ± 2.043 **** | 40.50 ± 1.660 ** | |
| SGR (%/day) | 7th-D | 0.567 ± 0.021 | 1.866 ± 0.183 *** | 2.015 ± 0.231 *** |
| 14th-D | 1.667 ± 0.201 | 4.152 ± 0.208 **** | 4.243 ± 0.170 **** | |
| 21st-D | 2.927 ± 0.197 | 2.892 ± 0.191 * | 4.382 ± 0.169 **** | |
| 28th-D | 4.055 ± 0.182 | 4.253 ± 0.209 * | 4.839 ± 0.168 * | |
| ADWG | 7th-D | 0.048 ± 0.001 | 0.168 ± 0.016 **** | 0.182 ± 0.021 **** |
| 14th-D | 0.151 ± 0.017 | 0.401 ± 0.017 **** | 0.419 ± 0.021 **** | |
| 21st-D | 0.276 ± 0.018 | 0.412 ± 0.016 | 0.435 ± 0.021 **** | |
| 28t-D | 0.397 ± 0.017 | 0.437 ± 0.016 | 0.488 ± 0.021 **** | |
| PER | 7th-D | 0.012 ± 0.001 | 0.034 ± 0.004 | 0.047 ± 0.005 **** |
| 14th-D | 0.039 ± 0.004 | 0.104 ± 0.003 | 0.109 ± 0.005 **** | |
| 21st-D | 0.072 ± 0.005 | 0.069 ± 0.004 | 0.113 ± 0.005 **** | |
| 28th-D | 0.103 ± 0.005 | 0.107 ± 0.004 | 0.127 ± 0.005 ** | |
| CF (g/cm3) | 7th-D | 1.151 ± 0.059 | 1.176 ± 0.087 | 1.004 ± 0.049 |
| 14th-D | 1.411 ± 0.087 | 1.568 ± 0.150 | 1.283 ± 0.097 | |
| 21st-D | 1.507 ± 0.092 | 1.426 ± 0.110 | 1.327 ± 0.107 | |
| 28th-D | 1.538 ± 0.071 | 1.585 ± 0.137 | 1.360 ± 0.107 | |
| HSI (%) | 7th-D | 1.529 ± 0.076 | 1.536 ± 0.089 | 1.543 ± 0.075 |
| 14th-D | 1.534 ± 0.069 | 1.537 ± 0.112 | 1.545 ± 0.067 | |
| 21st-D | 1.549 ± 0.081 | 1.557 ± 0.064 | 1.562 ± 0.0452 | |
| 28th-D | 1.558 ± 0.054 | 1.568 ± 0.086 | 1.575 ± 0.060 | |
| Survival rate (%) | 7th–28th-D | 100 | 100 | 100 |
| Parameters | Sampling Period (Days) | Dietary Black Seed Levels | ||
|---|---|---|---|---|
| Group-I (0%) | Group-II (1%) | Group-III (2.5%) | ||
| Moisture (%) | 7th-D | 74.61 ± 0.105 | 74.46 ± 0.065 | 73.99 ± 0.025 |
| 14th-D | 74.31 ± 0.105 | 74.35 ± 0.070 | 73.92 ± 0.045 | |
| 21st-D | 74.25 ± 0.255 | 74.28 ± 0.050 | 73.78 ± 0.115 | |
| 28th-D | 74.08 ± 0.095 | 74.07 ± 0.050 | 73.73 ± 0.160 | |
| Fat (%) | 7th-D | 3.100 ± 0.100 | 3.035 ± 0.065 | 2.915 ± 0.025 |
| 14th-D | 3.050 ± 0.050 | 3.010 ± 0.030 | 2.980 ± 0.010 | |
| 21st-D | 2.995 ± 0.015 | 2.915 ± 0.045 | 2.910 ± 0.050 | |
| 28th-D | 3.025 ± 0.075 | 2.900 ± 0.040 | 2.825 ± 0.065 | |
| Ash (%) | 7th-D | 2.655 ± 0.005 | 2.925 ± 0.055 | 2.855 ± 0.025 |
| 14th-D | 2.855 ± 0.015 | 2.950 ± 0.030 | 2.970 ± 0.010 | |
| 21st-D | 2.815 ± 0.055 | 2.925 ± 0.035 * | 2.945 ± 0.035 * | |
| 28th-D | 2.845 ± 0.025 | 2.930 ± 0.010 ** | 2.990 ± 0.030 ** | |
| Protein (%) | 7th-D | 16.48 ± 0.050 | 16.79 ± 0.005 * | 16.90 ± 0.015 ** |
| 14th-D | 16.64 ± 0.025 | 16.93 ± 0.045 ** | 17.00 ± 0.010 ** | |
| 21st-D | 16.67 ± 0.075 | 16.90 ± 0.080 * | 17.08 ± 0.045 * | |
| 28th-D | 16.76 ± 0.080 | 17.00 ± 0.010 * | 17.28 ± 0.005 ** | |
| Parameters | Sampling Period (Days) | Dietary Black Seed Levels | ||
|---|---|---|---|---|
| Group-I (0%) | Group-II (1%) | Group-III (2.5%) | ||
| ALP (U/l) | 7th-D | 460.0 ± 0.512 | 322.0 ± 0.018 ** | 384.9 ± 0.047 |
| 14th-D | 500.5 ± 0.123 | 350.2 ± 0.312 ** | 390.4 ± 0.051 * | |
| 21st-D | 510.5 ± 0.145 | 440.1 ± 0.541 * | 386.7 ± 0.110 * | |
| 28th-D | 548.9 ± 0.312 | 413.7 ± 0.113 ** | 417.7 ± 0.111 ** | |
| ALT (U/l) | 7th-D | 24.81 ± 0.013 | 24.36 ± 0.045 | 27.72 ± 0.110 |
| 14th-D | 25.61 ± 0.009 | 23.64 ± 0.020 | 28.75 ± 0.009 | |
| 21st-D | 26.01 ± 0.101 | 22.08 ± 0.011 | 21.11 ± 0.017 | |
| 28th-D | 26.38 ± 0.002 | 26.01 ± 0.013 | 25.22 ± 0.005 | |
| AST (U/l) | 7th-D | 26.77 ± 1.012 | 26.48 ± 0.016 | 22.98 ± 0.079 |
| 14th-D | 27.35 ± 1.000 | 22.57 ± 0.078 | 18.07 ± 0.065 | |
| 21st-D | 27.94 ± 0.998 | 27.40 ± 0.015 | 25.15 ± 0.103 * | |
| 28th-D | 28.52 ± 1.145 | 22.65 ± 0.046 | 21.69 ± 0.078 * | |
| Urea (mg/dl) | 7th-D | 2.963 ± 0.213 | 2.618 ± 0.167 | 2.550 ± 0.078 |
| 14th-D | 3.376 ± 0.136 | 2.481 ± 0.210 | 2.687 ± 0.106 | |
| 21st-D | 3.032 ± 0.178 | 2.825 ± 0.198 | 2.550 ± 0.117 | |
| 28th-D | 3.032 ± 0.145 | 2.550 ± 0.166 | 2.274 ± 0.178 * | |
| Creatinine (mg/dl) | 7th-D | 0.152 ± 0.056 | 0.143 ± 0.045 | 0.118 ± 0.012 |
| 14th-D | 0.168 ± 0.033 | 0.134 ± 0.034 | 0.125 ± 0.036 * | |
| 21st-D | 0.172 ± 0.098 | 0.125 ± 0.076 | 0.110 ± 0.067 ** | |
| 28th-D | 0.152 ± 0.097 | 0.116 ± 0.045 * | 0.089 ± 0.045 ** | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Latif, M.; Faheem, M.; Asmatullah; Hoseinifar, S.H.; Van Doan, H. Dietary Black Seed Effects on Growth Performance, Proximate Composition, Antioxidant and Histo-Biochemical Parameters of a Culturable Fish, Rohu (Labeo rohita). Animals 2021, 11, 48. https://doi.org/10.3390/ani11010048
Latif M, Faheem M, Asmatullah, Hoseinifar SH, Van Doan H. Dietary Black Seed Effects on Growth Performance, Proximate Composition, Antioxidant and Histo-Biochemical Parameters of a Culturable Fish, Rohu (Labeo rohita). Animals. 2021; 11(1):48. https://doi.org/10.3390/ani11010048
Chicago/Turabian StyleLatif, Maria, Mehwish Faheem, Asmatullah, Seyed Hossein Hoseinifar, and Hien Van Doan. 2021. "Dietary Black Seed Effects on Growth Performance, Proximate Composition, Antioxidant and Histo-Biochemical Parameters of a Culturable Fish, Rohu (Labeo rohita)" Animals 11, no. 1: 48. https://doi.org/10.3390/ani11010048