The Probiotic Lactobacillus fermentum Biocenol CCM 7514 Moderates Campylobacter jejuni-Induced Body Weight Impairment by Improving Gut Morphometry and Regulating Cecal Cytokine Abundance in Broiler Chickens

 ,
,  ,
,  ,
,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Chickens and Experimental Scheme
2.2. Body Weight of Chickens
2.3. Morphometrical Analyses
2.4. RNA Extraction and RT-qPCR Assays
2.5. Statistical Analyes
3. Results
3.1. Chickens’ Body Weight
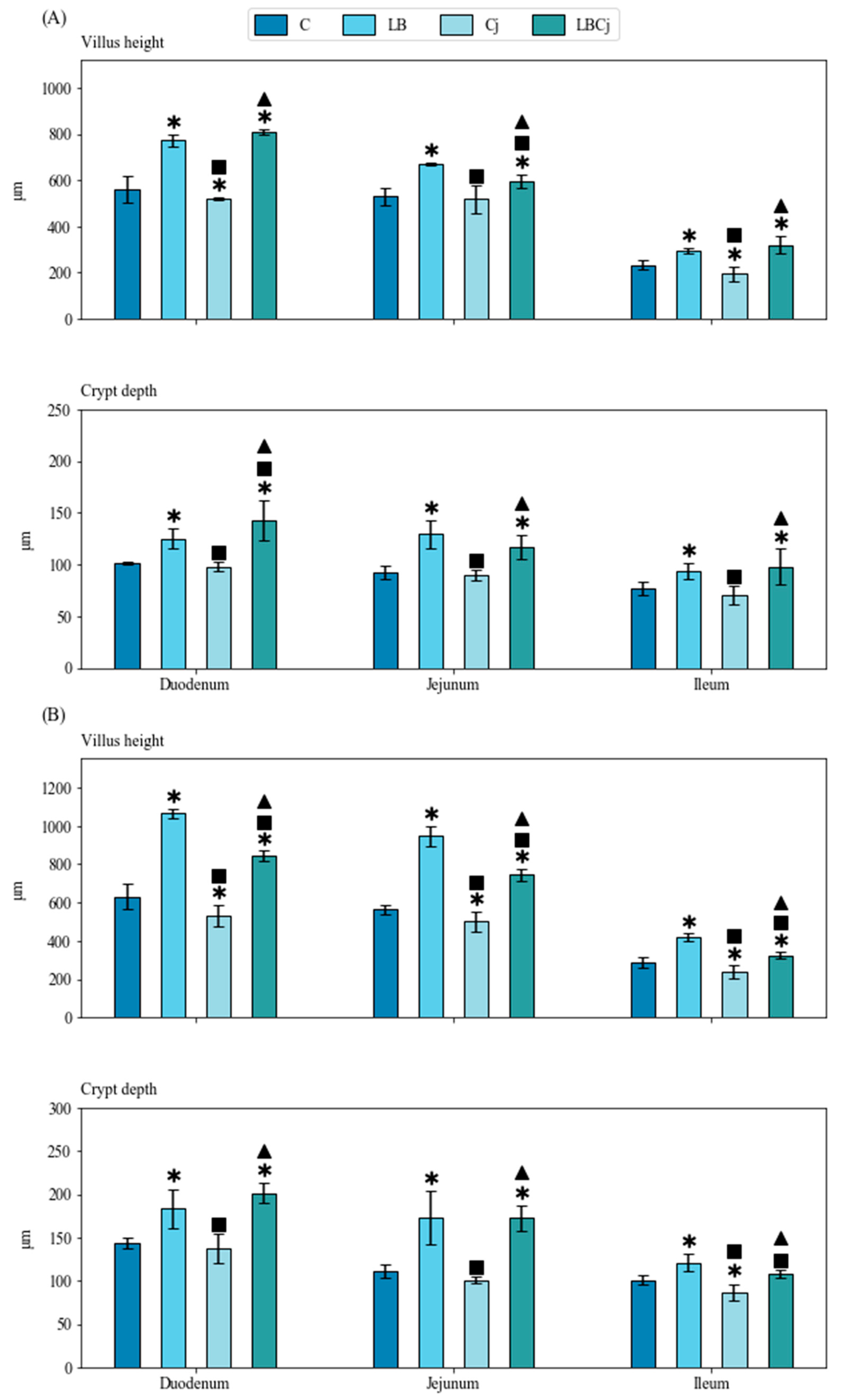
3.2. Morphometrical Analyses
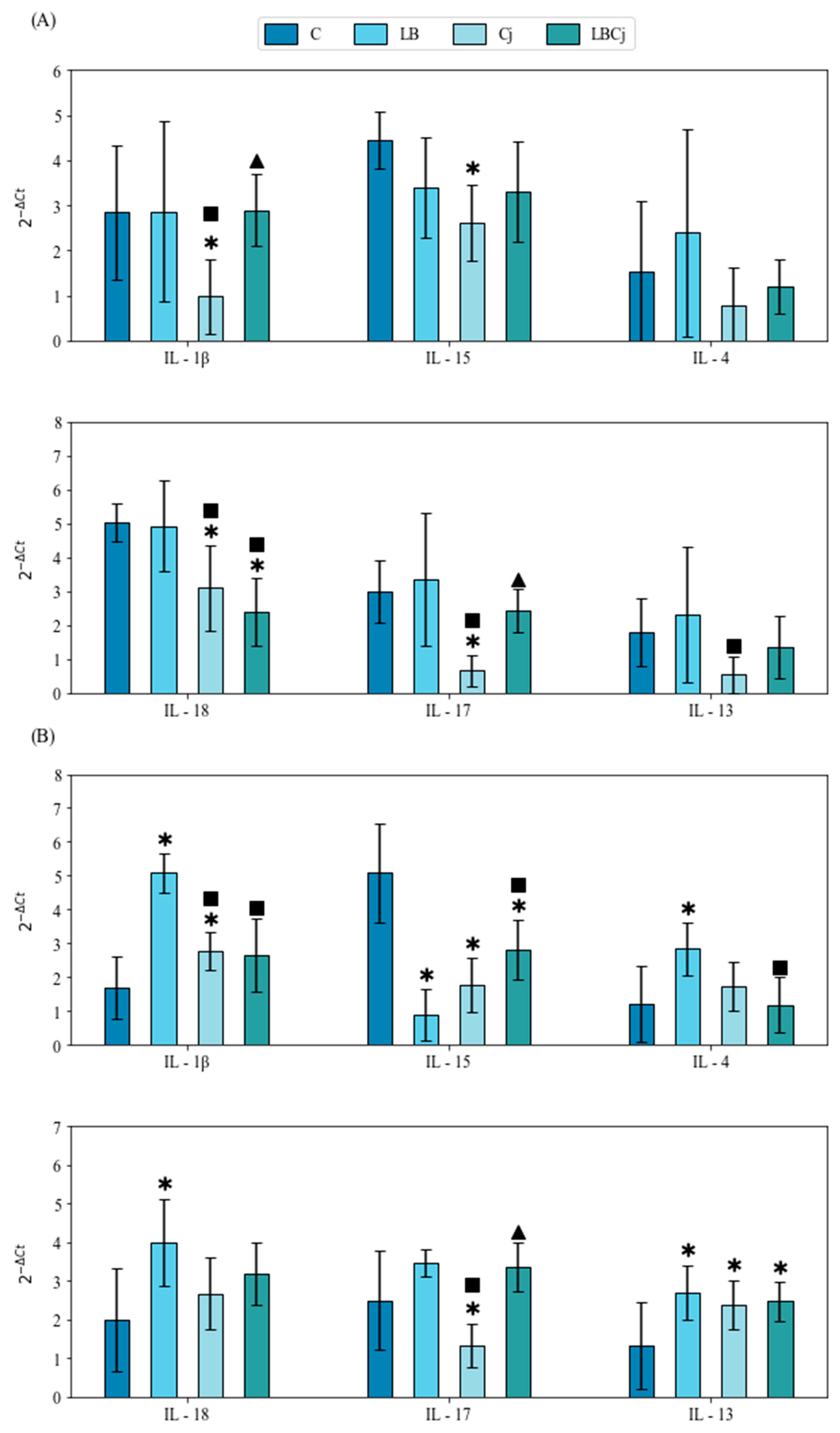
3.3. Cytokine Response
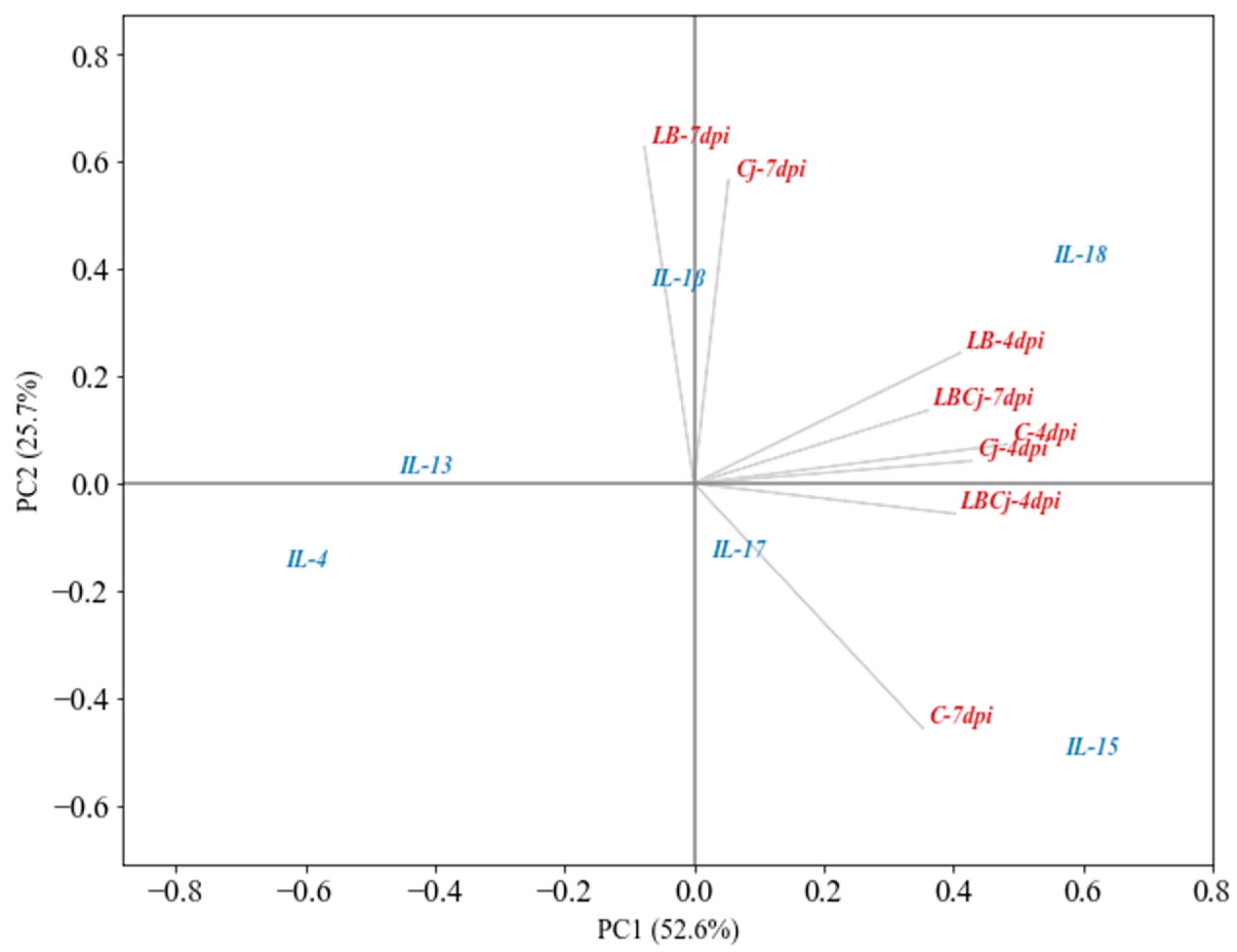
3.4. Correlation Analysis between Evaluated Indicators
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Pan, D.; Yu, Z. Intestinal microbiome of poultry and its interaction with host and diet. Gut Microbes 2014, 5, 108–119. [Google Scholar] [CrossRef]
- Awad, W.A.; Hess, C.; Hess, M. Enteric pathogens and their toxin-induced disruption of the intestinal barrier through alteration of tight junctions in chickens. Toxins 2017, 9, 60. [Google Scholar] [CrossRef] [Green Version]
- Kuhn, K.G.; Falkenhorst, G.; Emborg, H.D.; Ceper, T.; Torpdahl, M.; Krogfelt, K.A.; Ethelberg, S.; Mølbak, K. Epidemiological and serological investigation of a waterborne Campylobacter jejuni outbreak in a Danish town. Epidemiol. Infect. 2017, 145, 701–709. [Google Scholar] [CrossRef] [Green Version]
- Silva, W.C.; Targino, B.N.; Mendonça, R.S.; Sant’Ana, A.S.; Hungaro, H.M. Campylobacter: An overview of cases, occurrence in food, contamination sources, and antimicrobial resistance in Brazil. Food Rev. Int. 2018, 34, 364–389. [Google Scholar] [CrossRef]
- Connerton, P.L.; Richards, P.J.; Lafontaine, G.M.; O’Kane, P.M.; Ghaffar, N.; Cummings, N.J.; Smith, D.L.; Neville, M.F.; Connerton, I.F. The effect of the timing of exposure to Campylobacter jejuni on the gut microbiome and inflammatory responses of broiler chickens. Microbiome 2018, 6, 88. [Google Scholar] [CrossRef]
- Stephenson, H.N.; John, C.M.; Naz, N.; Gundogdu, O.; Dorrell, N.; Wren, B.W.; Jarvis, G.A.; Bajaj-Elliott, M. Campylobacter jejuni lipooligosaccharide sialylation, phosphorylation, and amide/ester linkage modifications fine-tune human Toll-like receptor 4 activation. J. Biol. Chem. 2013, 288, 19661–19672. [Google Scholar] [CrossRef] [Green Version]
- Shaughnessy, R.G.; Meade, K.G.; McGivney, B.A.; Allan, B.; O’Farrelly, C. Global gene expression analysis of chicken caecal response to Campylobacter jejuni. Vet. Immunol. Immunopathol. 2011, 142, 64–71. [Google Scholar] [CrossRef]
- Dunislawska, A.; Slawinska, A.; Stadnicka, K.; Bednarczyk, M.; Gulewicz, P.; Jozefiak, D.; Siwek, M. Synbiotics for broiler chickens—In vitro design and evaluation of the influence on host and selected microbiota populations following in ovo delivery. PLoS ONE 2017, 12, e0168587. [Google Scholar] [CrossRef] [Green Version]
- Lacharme-Lora, L.; Chaloner, G.; Gilroy, R.; Humphrey, S.; Gibbs, K.; Jopson, S.; Wright, E.; Reid, W.; Ketley, J.; Humphrey, T.; et al. B lymphocytes play a limited role in clearance of Campylobacter jejuni from the chicken intestinal tract. Sci. Rep. 2017, 7, 45090. [Google Scholar] [CrossRef]
- Karaffová, V.; Revajová, V.; Koščová, J.; Gancarčíková, S.; Nemcová, R.; Ševčíková, Z.; Herich, R.; Levkut, M. Local intestinal immune response including NLRP3 inflammasome in broiler chicken infected with Campylobacter jejuni after administration of Lactobacillus reuteri B1/1. Food Agric. Immunol. 2020, 31, 954–966. [Google Scholar] [CrossRef]
- Šefcová, M.; Larrea-Álvarez, M.; Larrea-Álvarez, C.; Revajová, V.; Karaffová, V.; Koščová, J.; Nemcová, R.; Ortega-Paredes, D.; Vinueza-Burgos, C.H.; Levkut, M.; et al. Effects of Lactobacillus Fermentum Supplementation on Body Weight and Pro-Inflammatory Cytokine Expression in Campylobacter Jejuni-Challenged Chickens. Vet. Sci. 2020, 7, 121. [Google Scholar] [CrossRef]
- Mortada, M.; Cosby, D.E.; Shanmugasundaram, R.; Selvaraj, R.K. In vivo and in vitro assessment of commercial probiotic and organic acid feed additives in broilers challenged with Campylobacter coli. J. Appl. Poult. Res. 2020, 29, 435–446. [Google Scholar] [CrossRef]
- Flaujac Lafontaine, G.M.; Richards, P.J.; Connerton, P.L.; O’Kane, P.M.; Ghaffar, N.M.; Cummings, N.J.; Fish, N.M.; Connerton, I.F. Prebiotic driven increases in IL-17A do not prevent Campylobacter jejuni colonization of chickens. Front. Microbiol. 2020, 10, 3030. [Google Scholar] [CrossRef] [Green Version]
- Nyati, K.K.; Prasad, K.N.; Agrawal, V.; Husain, N. Matrix metalloproteinases-2 and-9 in Campylobacter jejuni-induced paralytic neuropathy resembling Guillain-Barré syndrome in chickens. Microb. Pathog. 2017, 111, 395–401. [Google Scholar] [CrossRef]
- Johnson, T.J.; Shank, J.M.; Johnson, J.G. Current and potential treatments for reducing Campylobacter colonization in animal hosts and disease in humans. Front. Microbiol. 2017, 8, 487. [Google Scholar] [CrossRef] [Green Version]
- Kelly, C.; Gundogdu, O.; Pircalabioru, G.; Cean, A.; Scates, P.; Linton, M.; Pinkerton, L.; Magowan, E.; Stef, L.; Simiz, E.; et al. The in vitro and in vivo effect of carvacrol in preventing Campylobacter infection, colonization and in improving productivity of chicken broilers. Foodborne Pathog. Dis. 2017, 14, 341–349. [Google Scholar] [CrossRef]
- Abudabos, A.M.; Alyemni, A.H.; Dafalla, Y.M.; Khan, R.U. The effect of phytogenics on growth traits, blood biochemical and intestinal histology in broiler chickens exposed to Clostridium perfringens challenge. J. Appl. Anim. Res. 2018, 46, 691–695. [Google Scholar] [CrossRef] [Green Version]
- Rubio, L.A. Possibilities of early life programming in broiler chickens via intestinal microbiota modulation. Poult. Sci. 2019, 98, 695–706. [Google Scholar] [CrossRef]
- Baldwin, S.; Hughes, R.J.; Hao Van, T.T.; Moore, R.J.; Stanley, D. At-hatch administration of probiotic to chickens can introduce beneficial changes in gut microbiota. PLoS ONE 2018, 13, e0194825. [Google Scholar] [CrossRef]
- Manes-Lazaro, R.; Van Diemen, P.M.; Pin, C.; Mayer, M.J.; Stevens, M.P.; Narbad, A. Administration of Lactobacillus johnsonii FI9785 to chickens affects colonisation by Campylobacter jejuni and the intestinal microbiota. Br. Poult Sci. 2017, 58, 373–381. [Google Scholar] [CrossRef] [Green Version]
- Saint-Cyr, M.J.; Haddad, N.; Taminiau, B.; Poezevara, T.; Quesne, S.; Amelot, M.; Daubeb, G.; Chemalyc, M.; Dousset, X.; Guyard-Nicodèmec, M. Use of the potential probiotic strain Lactobacillus salivarius SMXD51 to control Campylobacter jejuni in broilers. Int. J. Food Microbiol. 2017, 247, 9–17. [Google Scholar] [CrossRef]
- Karaffová, V.; Marcinková, E.; Bobíková, K.; Herich, R.; Revajová, V.; Stašová, D.; Kavuľová, A.; Levkutová, M.; Levkut, M., Jr.; Lauková, A.; et al. TLR4 and TLR21 expression, MIF, IFN-β, MD-2, CD14 activation, and sIgA production in chickens administered with EFAL41 strain challenged with Campylobacter jejuni. Folia Microbiol. (Praha) 2017, 62, 89–97. [Google Scholar]
- Šefcová, M.; Larrea-Álvarez, M.; Larrea-Álvarez, C.; Karaffová, V.; Revajová, V.; Gancarčíková, S.; Ševčíková, Z.; Herich, R. Lactobacillus fermentum Administration Modulates Cytokine Expression and Lymphocyte Subpopulation Levels in Broiler Chickens Challenged with Campylobacter coli. Foodborne Pathog. Dis. 2020, 17, 485–493. [Google Scholar] [CrossRef]
- Cobb-Vantress. Broiler Management Guide. Available online: https://cobb-vantress.com (accessed on 4 December 2020).
- Nain, S.; Renema, R.A.; Zuidhof, M.J.; Korver, D.R. Effect of metabolic efficiency and intestinal morphology on variability in n-3 polyunsaturated fatty acid enrichment of eggs. Poult. Sci. 2012, 91, 888–898. [Google Scholar] [CrossRef]
- Šefcová, M.; Levkut, M.; Bobíková, K.; Karaffová, V.; Revajová, V.; Cingeľová Maruščáková, I.; Levkutová, M.; Ševčíková, Z.; Herich, R.; Levkut, M. Cytokine response after stimulation of culture cells by zinc and probiotic strain. In Vitr. Cell. Dev. Biol. Anim. 2019, 55, 830–837. [Google Scholar] [CrossRef]
- Champigny, M.J.; Sung, W.W.L.; Catana, V.; Salwan, R.; Summers, P.S.; Dudley, S.A.; Provart, N.J.; Cameron, R.K.; Golding, G.B.; Weretilnyk, E.A. RNA-Seq effectively monitors gene expression in Eutrema salsugineum plants growing in an extreme natural habitat and in controlled growth cabinet conditions. BMC Genom. 2013, 14, 578. [Google Scholar] [CrossRef] [Green Version]
- Oh, S.; Lillehoj, H.S.; Lee, Y.; Bravo, D.; Lillehoj, E. Dietary antibiotic growth promoters down-regulate intestinal inflammatory cytokine expression in chickens challenged with LPS or co-infected with Eimeria maxima and Clostridium perfringens. Front. Vet. Sci. 2019, 6, 420. [Google Scholar] [CrossRef] [Green Version]
- Sureshkumar, S.; Jung, S.K.; Kim, D.; Oh, K.B.; Yang, H.; Lee, H.C.; Jin, Y.J.; Lee, H.S.; Lee, S.; Byun, S.J. Oral administration of Lactobacillus reuteri expressing a 3D8 single-chain variable fragment (scFv) enhances chicken growth and conserves immune homeostasis. 3 Biotech. 2019, 9, 282. [Google Scholar] [CrossRef]
- Forte, C.; Manuali, E.; Abbate, Y.; Papa, P.; Vieceli, L.; Tentellini, M.; Trabalza-Marinucci, M.; Moscati, L. Dietary Lactobacillus acidophilus positively influences growth performance, gut morphology, and gut microbiology in rurally reared chickens. Poult. Sci. 2018, 97, 930–936. [Google Scholar] [CrossRef]
- Biasato, I.; Ferrocino, I.; Biasibetti, E.; Grego, E.; Dabbou, S.; Sereno, A.; Gai, F.; Gasco, L.; Schiavone, A.; Cocolin, l.; et al. Modulation of intestinal microbiota, morphology and mucin composition by dietary insect meal inclusion in free-range chickens. BMC Vet. Res. 2018, 14, 383. [Google Scholar] [CrossRef] [Green Version]
- Swaggerty, C.L.; Callaway, T.R.; Kogut, M.H.; Piva, A.; Grilli, E. Modulation of the immune response to improve health and reduce foodborne pathogens in poultry. Microorganisms 2019, 7, 65. [Google Scholar] [CrossRef] [Green Version]
- Castro, M.S.; Molina, M.A.; Azpiroz, M.B.; Díaz, A.M.; Ponzio, R.; Sparo, M.D.; Manghi, M.A.; Canellada, A.M. Probiotic activity of Enterococcus faecalis CECT 7121: Effects on mucosal immunity and intestinal epithelial cells. J. Appl. Microbiol. 2016, 121, 1117–1129. [Google Scholar] [CrossRef] [Green Version]
- Zhai, Z.; Torres-Fuentes, C.; Heeney, D.D.; Marco, M.L. Synergy between probiotic Lactobacillus casei and milk to maintain barrier integrity of intestinal epithelial cells. J. Agric. Food Chem. 2019, 67, 1955–1962. [Google Scholar] [CrossRef]
- Shini, S.; Zhang, D.; Aland, R.C.; Li, X.; Dart, P.J.; Callaghan, M.J.; Speight, R.E.; Bryden, W.L. Probiotic Bacillus amyloliquefaciens H57 ameliorates subclinical necrotic enteritis in broiler chicks by maintaining intestinal mucosal integrity and improving feed efficiency. Poult. Sci. 2020, 99, 4278–4293. [Google Scholar] [CrossRef]
- Ho, S.W.; El-Nezami, H.; Shah, N.P. The protective effects of enriched citrulline fermented milk with Lactobacillus helveticus on the intestinal epithelium integrity against Escherichia coli infection. Sci. Rep. 2020, 10, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Dela Cruz, P.J.D.; Dagaas, C.T.; Mangubat, K.M.M.; Angeles, A.A.; Abanto, O.D. Dietary effects of commercial probiotics on growth performance, digestibility, and intestinal morphometry of broiler chickens. Trop. Anim. Health Prod. 2019, 51, 1105–1115. [Google Scholar] [CrossRef]
- Bogucka, J.; Ribeiro, D.M.; Bogusławska-Tryk, M.; Dankowiakowska, A.; da Costa, R.P.R.; Bednarczyk, M. Microstructure of the small intestine in broiler chickens fed a diet with probiotic or synbiotic supplementation. J. Anim. Physiol. Anim. Nutr. (Berl) 2019, 103, 1785–1791. [Google Scholar] [CrossRef]
- Saenphoom, P.; Liang, J.B.; Ho, Y.W.; Loh, T.C.; Rosfarizan, M. Effects of enzyme treated palm kernel expeller on metabolizable energy, growth performance, villus height and digesta viscosity in broiler chickens. Asian-Australasian J. Anim. Sci. 2013, 26, 537. [Google Scholar] [CrossRef] [Green Version]
- Silva, M.A.; Sousa Pessotti, B.M.; Zanini, S.F.; Colnago, G.L.; Alves Rodrigues, M.R.; Carvalho Nunes, L.; Santos Zanini, M.; Freire Martins, I.V. Intestinal mucosa structure of broiler chickens infected experimentally with Eimeria tenella and treated with essential oil of oregano. Ciência Rural 2009, 39, 1471–1477. [Google Scholar] [CrossRef] [Green Version]
- Matur, E.; Eraslan, E. The impact of probiotics on the gastrointestinal physiology. New Adv. Basic Clin. Gastroenterol. 2012, 1, 51–74. [Google Scholar]
- Wu, H.; Ye, L.; Lu, X.; Xie, S.; Yang, Q.; Yu, Q. Lactobacillus acidophilus Alleviated Salmonella-Induced Goblet Cells Loss and Colitis by Notch Pathway. Mol. Nutr. Food Res. 2018, 62, 1800552. [Google Scholar] [CrossRef]
- Lu, X.; Xie, S.; Ye, L.; Zhu, L.; Yu, Q. Lactobacillus protects against S. Typhimurium–induced intestinal inflammation by determining the fate of epithelial proliferation and differentiation. Mol. Nutr. Food Res. 2020, 64, 1900655. [Google Scholar] [CrossRef]
- Fonseca, B.B.; Fernandez, H.; Rossi, D.A. Campylobacter spp. and Related Organisms in Poultry; Springer: Cham, Switzerland, 2016. [Google Scholar]
- Smith, C.K.; AbuOun, M.; Cawthraw, S.A.; Humphrey, T.J.; Rothwell, L.; Kaiser, P.; Barrow, P.A.; Jones, M.A. Campylobacter colonization of the chicken induces a proinflammatory response in mucosal tissues. FEMS Immunol. Med. Microbiol. 2008, 54, 114–121. [Google Scholar] [CrossRef] [Green Version]
- Hermans, D.; Pasmans, F.; Heyndrickx, M.; Van Immerseel, F.; Martel, A.; Van Deun, K.; Haesebrouck, F. A tolerogenic mucosal immune response leads to persistent Campylobacter jejuni colonization in the chicken gut. Crit. Rev. Microbiol. 2012, 38, 17–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bron, P.A.; Van Baarlen, P.; Kleerebezem, M. Emerging molecular insights into the interaction between probiotics and the host intestinal mucosa. Nat. Rev. Microbiol. 2012, 10, 66–78. [Google Scholar] [CrossRef] [PubMed]
- Barjesteh, N.; Hodgins, D.C.; Paul, M.S.; Quinteiro-Filho, W.M.; DePass, C.; Monteiro, M.A.; Sharif, S. Induction of chicken cytokine responses in vivo and in vitro by lipooligosaccharide of Campylobacter jejuni HS: 10. Vet. Microbiol. 2013, 164, 122–130. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhao, H.; Shao, Y.; Liu, J.; Li, J.; Xing, M. Copper or/and arsenic induce oxidative stress-cascaded, nuclear factor kappa B-dependent inflammation and immune imbalance, trigging heat shock response in the kidney of chicken. Oncotarget 2017, 8, 98103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Castejon, G.; Brough, D. Understanding the mechanism of IL-1β secretion. Cytokine Growth Factor Rev. 2011, 22, 189–195. [Google Scholar] [CrossRef]
- Miettinen, M.; Pietilä, T.E.; Kekkonen, R.A.; Kankainen, M.; Latvala, S.; Pirhonen, J.; Österlund, P.; Korpela, R.; Julkunen, I. Nonpathogenic Lactobacillus rhamnosus activates the inflammasome and antiviral responses in human macrophages. Gut Microbes 2012, 3, 510–522. [Google Scholar] [CrossRef] [Green Version]
- He, H.; Genovese, K.J.; Kogut, M.H. Modulation of chicken macrophage effector function by TH1/TH2 cytokines. Cytokine 2011, 53, 363–369. [Google Scholar] [CrossRef]
- Brisbin, J.T.; Gong, J.; Parvizi, P.; Sharif, S. Effects of lactobacilli on cytokine expression by chicken spleen and cecal tonsil cells. Clin. Vaccine Immunol. 2010, 17, 1337–1343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mensikova, M.; Stepanova, H.; Faldyna, M. Interleukin-17 in veterinary animal species and its role in various diseases: A review. Cytokine 2013, 64, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Konjar, Š.; Ferreira, C.; Blankenhaus, B.; Veldhoen, M. Intestinal barrier interactions with specialized CD8 T cells. Front. Immunol. 2017, 8, 1281. [Google Scholar] [CrossRef] [PubMed]
- Al-Banna, N.A.; Cyprian, F.; Albert, M.J. Cytokine responses in campylobacteriosis: Linking pathogenesis to immunity. Cytokine Growth Factor Rev. 2018, 41, 75–87. [Google Scholar] [CrossRef] [PubMed]
- Monge, S.; Teunis, P.; Friesema, I.; Franz, E.; Wim, A.; van Pelt, W.; Mughini-Grasad, L. Immune response-eliciting exposure to Campylobacter vastly exceeds the incidence of clinically overt campylobacteriosis but is associated with similar risk factors: A nationwide serosurvey in the Netherlands. J. Infect. 2018, 77, 171–177. [Google Scholar] [CrossRef]
- Havelaar, A.H.; van Pelt, W.; Ang, C.W.; Wagenaar, J.A.; van Putten, J.P.M.; Gross, U.; Newell, D.G. Immunity to Campylobacter: Its role in risk assessment and epidemiology. Crit. Rev. Microbiol. 2009, 35, 1–22. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, R.; Song, M.; Hu, Y.; Pan, B.; Cai, J.; Wang, M. Eimeria tenella: Interleukin 17 contributes to host immunopathology in the gut during experimental infection. Exp. Parasitol. 2013, 133, 121–130. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Day | Control (Birds) | L fermentum Treatment, per os (Birds) | C. jejuni Treatment, per os (Birds) | Coexposure Treatment, per os (Birds) |
|---|---|---|---|---|
| 0 d | 18 | 18 | 18 | 18 |
| 1–3 d | 18 | 18 109 CFU/0.2 mL (Probiotic) | 18 | 18 109 CFU/0.2 mL (Probiotic) |
| 4 d | 18 | 18 109 CFU/0.2 mL (Probiotic) | 18 108 CFU/0.2 mL (C. jejuni) | 18 109 CFU/0.2 mL (Probiotic) + 108 CFU/0.2 mL (C. jejuni) |
| 5–7 d | 18 | 18 109 CFU/0.2 mL (Probiotic) | 18 | 18 109 CFU/0.2 mL (Probiotic) per os |
| 8 d (4 dpi) sample collection | 9 | 9 | 9 | 9 |
| 9–10 d | 9 | 9 | 9 | 9 |
| 11 d (7 dpi) sample collection | 9 | 9 | 9 | 9 |
| Small Intestine Sections | Experimental Groups | |||
|---|---|---|---|---|
| C | LB | Cj | LBCj | |
| 4 dpi | ||||
| Duodenum | 5.53 ± 0.55 | 6.20 ± 0.37 | 5.29 ± 0.37 ■ | 5.68 ± 0.83 |
| Jejunum | 5.76 ± 0.47 | 5.19 ± 0.64 | 5.80 ± 0.61 | 5.13 ± 0.63 |
| Ileum | 3.04 ± 0.33 | 3.18 ± 0.23 | 2.87 ± 0.74 | 3.35 ± 0.64 |
| 7 dpi | ||||
| Duodenum | 4.40 ± 0.51 | 5.88 ± 0.60 * | 3.95 ± 0.66 ■ | 4.19 ± 0.21 ■ |
| Jejunum | 5.09 ± 0.44 | 5.69 ± 1.4 | 4.96 ± 0.56 | 4.35 ± 0.37 ■ |
| Ileum | 2.87 ± 0.34 | 3.50 ± 0.39* | 2.75 ± 0.34 ■ | 3.00 ± 0.16 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Šefcová, M.A.; Larrea-Álvarez, M.; Larrea-Álvarez, C.M.; Karaffová, V.; Ortega-Paredes, D.; Vinueza-Burgos, C.; Ševčíková, Z.; Levkut, M.; Herich, R.; Revajová, V. The Probiotic Lactobacillus fermentum Biocenol CCM 7514 Moderates Campylobacter jejuni-Induced Body Weight Impairment by Improving Gut Morphometry and Regulating Cecal Cytokine Abundance in Broiler Chickens. Animals 2021, 11, 235. https://doi.org/10.3390/ani11010235
Šefcová MA, Larrea-Álvarez M, Larrea-Álvarez CM, Karaffová V, Ortega-Paredes D, Vinueza-Burgos C, Ševčíková Z, Levkut M, Herich R, Revajová V. The Probiotic Lactobacillus fermentum Biocenol CCM 7514 Moderates Campylobacter jejuni-Induced Body Weight Impairment by Improving Gut Morphometry and Regulating Cecal Cytokine Abundance in Broiler Chickens. Animals. 2021; 11(1):235. https://doi.org/10.3390/ani11010235
Chicago/Turabian StyleŠefcová, Miroslava Anna, Marco Larrea-Álvarez, César Marcelo Larrea-Álvarez, Viera Karaffová, David Ortega-Paredes, Christian Vinueza-Burgos, Zuzana Ševčíková, Mikuláš Levkut, Róbert Herich, and Viera Revajová. 2021. "The Probiotic Lactobacillus fermentum Biocenol CCM 7514 Moderates Campylobacter jejuni-Induced Body Weight Impairment by Improving Gut Morphometry and Regulating Cecal Cytokine Abundance in Broiler Chickens" Animals 11, no. 1: 235. https://doi.org/10.3390/ani11010235