Clostridium perfringens as Foodborne Pathogen in Broiler Production: Pathophysiology and Potential Strategies for Controlling Necrotic Enteritis

 ,
,  ,
,  ,
,
Abstract
:Simple Summary
Abstract

1. Introduction
2. C. perfringens as a Foodborne Pathogen
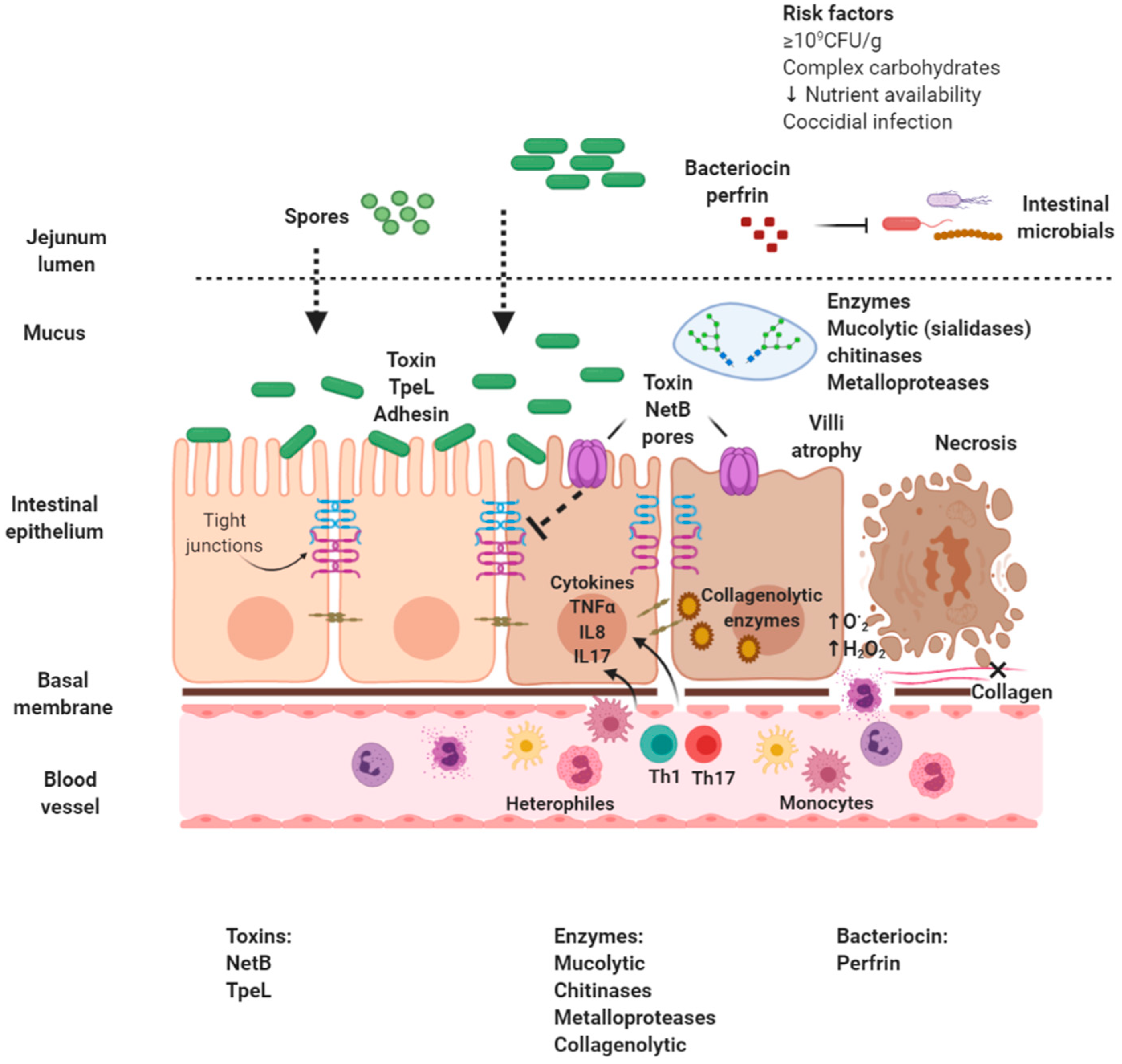
3. Necrotic Enteritis Pathophysiology
3.1. Clinical Alterations
3.2. Immune System Activity
4. Detection Mechanisms
4.1. Histopathological Detection
4.2. Immunological Detection
4.3. Molecular Detection
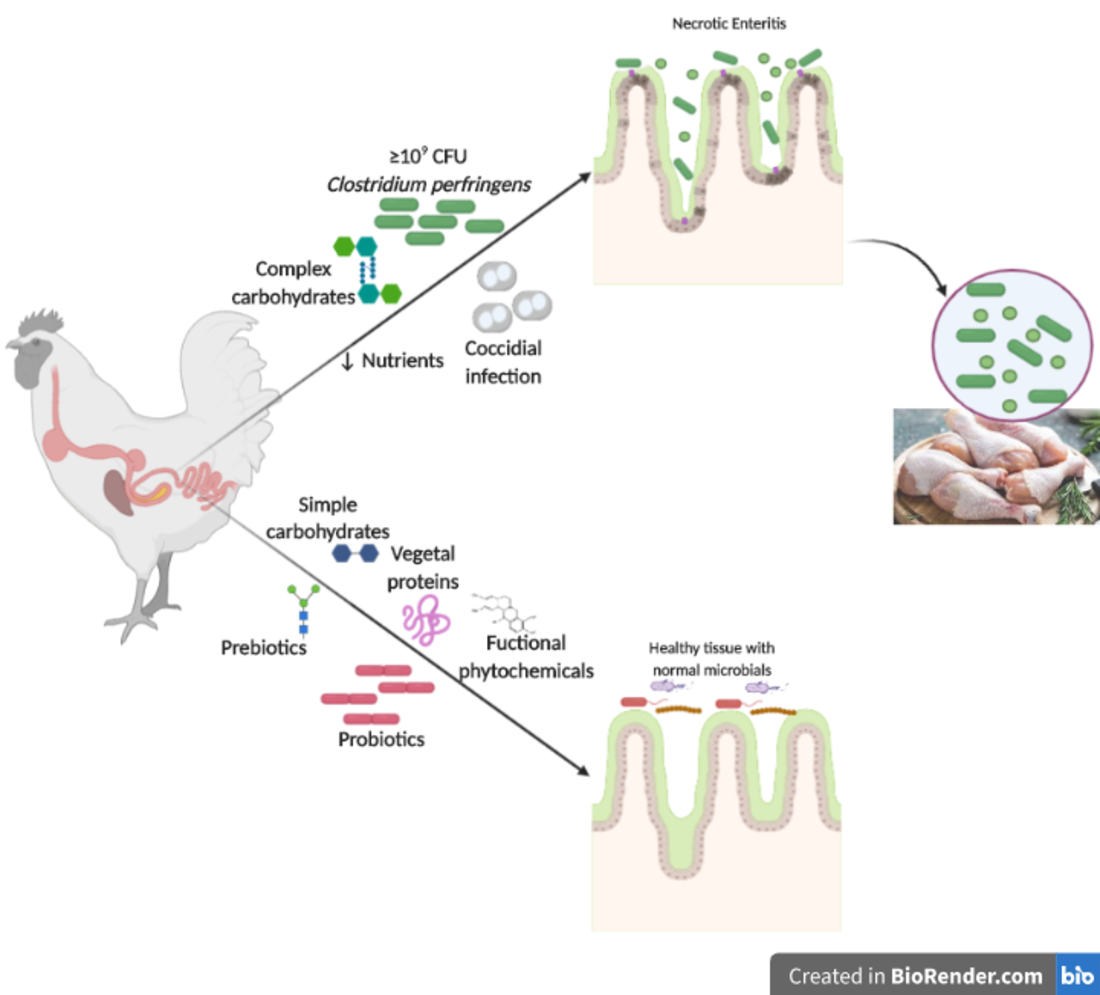
5. Control
5.1. Probiotics
5.2. Prebiotics
5.3. Synbiotics
5.4. Phytogenics
5.5. Organic Acids
5.6. Dietary Modifications and Enzymes
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Sarker, M.R.; Shivers, R.P.; Sparks, S.G.; Juneja, V.K.; McClane, B.A. Comparative experiments to examine the effects of heating on vegetative cells and spores of Clostridium perfringens isolates carrying plasmid genes versus chromosomal enterotoxin genes. Appl. Environ. Microbiol. 2000, 66, 3234–3240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juneja, V.K.; Novak, J.S.; Labbe, R.J. Clostridium perfringens. In Pathogens and Toxins in Foods: Challenges and Interventions; Juneja, V.K., Sofos, J.N., Eds.; ASM Press: Washington, DC, USA, 2010; Volume 34, pp. 53–70. [Google Scholar]
- Mcclane, B.A.; Uzal, F.A.; Fernandez, M.; Lyerly, D.; Wilkins, T. The Enterotoxic Clostridia. In The Prokaryotes. Bacteria: Firmicutes, Cyanobacteria; Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.-H., Stackebrandt, E., Eds.; Springer Science: Singapore, 2006; p. 1186. ISBN 978-0-387-25494-4. [Google Scholar]
- Al-Khaldi, S. Clostridium perfringens, phytohaemagglutinin (kidney bean lectin),Yersinia species. In Bad Bug Book. Foodborne Pathogenic Microorganisms and Natural Toxins; Lampel, K.A., Al-Khaldi, S., Cahill, S.M., Eds.; International Medical Publishing: McLean, VA, USA, 2012; ISBN 9780323401814. [Google Scholar]
- Geissmann, T.A.; Teuber, M.; Meile, L. Transcriptional analysis of the rubrerythrin and superoxide dismutase genes of Clostridium perfringens. J. Bacteriol. 1999, 181, 7136–7139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jean, D.; Briolat, V.; Reysset, G. Oxidative stress response in Clostridium perfringens. Microbiology 2004, 150, 1649–1659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johansson, A.; Aspan, A.; Bagge, E.; Båverud, V.; Engström, B.E.; Johansson, K.E. Genetic diversity of Clostridium perfringens type A isolates from animals, food poisoning outbreaks and sludge. BMC Microbiol. 2006, 6, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saito, M. Production of enterotoxin by Clostridium perfringens derived from humans, animals, foods, and the natural environment in Japan. J. Food Prot. 1990, 53, 115–118. [Google Scholar] [CrossRef] [PubMed]
- Grass, J.; Gould, L.H.; Mahon, B. Epidemiology of Foodborne Disease Outbreaks Caused by Clostridium perfringens, United States, 1998–2010. Foodborne Pathog. Dis. 2013, 10, 131–136. [Google Scholar] [CrossRef] [Green Version]
- Mellou, K.; Kyritsi, M.; Chrysostomou, A.; Sideroglou, T.; Georgakopoulou, T.; Hadjichristodoulou, C. Clostridium perfringens foodborne outbreak during an athletic event in northern Greece, June 2019. Int. J. Environ. Res. Public Health 2019, 16, 3967. [Google Scholar] [CrossRef] [Green Version]
- Monma, C.; Hatakeyama, K.; Obata, H.; Yokoyama, K.; Konishi, N.; Itoh, T.; Kai, A. Four foodborne disease outbreaks caused by a new type of enterotoxin-producing Clostridium perfringens. J. Clin. Microbiol. 2015, 53, 859–867. [Google Scholar] [CrossRef] [Green Version]
- Shaltout, A.; Zakaria, M.; Nabil, E. Detection and typing of Clostridium perfringens in some retail chicken meat products. Benha Vet. Med. J. 2017, 33, 283–291. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.A.; Labbé, R. Distribution of enterotoxin- and epsilon-positive Clostridium perfringens spores in U.S. retail spices. J. Food Prot. 2018, 81, 394–399. [Google Scholar] [CrossRef]
- Butler, A.J.; Thomas, M.K.; Pintar, K.D.M. Expert elicitation as a means to attribute 28 enteric pathogens to foodborne, waterborne, animal contact, and person-to-person transmission routes in Canada. Foodborne Pathog. Dis. 2015, 12, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Fohler, S.; Klein, G.; Hoedemaker, M.; Scheu, T.; Seyboldt, C.; Campe, A.; Jensen, K.C.; Abdulmawjood, A. Diversity of Clostridium perfringens toxin-genotypes from dairy farms. BMC Microbiol. 2016, 16, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Centers for Disease Control and Prevention (CDC) Surveillance for Foodborne Disease Outbreaks United States, 2017: Annual Report. U.S. Department of Health and Human Services: Atlanta, GA, USA, 2019; Volume 62, pp. 1–10.
- May, F.J.; Polkinghorne, B.G.; Fearnley, E.J. Epidemiology of bacterial toxin—mediated foodborne gastroenteritis outbreaks in Australia, 2001 to 2013. CDI 2016, 40, 460–469. [Google Scholar]
- Makkar, H.P.S.; Ankers, P. Towards sustainable animal diets: A survey-based study. Anim. Feed Sci. Technol. 2014, 198, 309–322. [Google Scholar] [CrossRef]
- Kay, S.; Edwards, J.; Brown, J.; Dixon, R. Galleria mellonella infection model identifies both high and low lethality of Clostridium perfringens toxigenic strains and their response to antimicrobials. Front. Microbiol. 2019, 10, 1–11. [Google Scholar] [CrossRef]
- Kiu, R.; Hall, L.J. An update on the human and animal enteric pathogen Clostridium perfringens. Emerg. Microbes Infect. 2018, 7. [Google Scholar] [CrossRef] [Green Version]
- Rood, J.I.; Adams, V.; Lacey, J.; Lyras, D.; Mcclane, B.A.; Stephen, B.; Moore, R.J.; Popoff, M.R.; Sarker, M.R.; Songer, J.G.; et al. Expansion of the Clostridium perfringens toxin-based typing scheme. Anaerobe 2018, 5–10. [Google Scholar] [CrossRef]
- Uzal, F.A.; Freedman, J.C.; Shrestha, A.; Theoret, J.R.; Garcia, J.; Awad, M.M.; Adams, V.; Moore, R.J.; Rood, J.I.; Mcclane, B.A. Towards an understanding of the role of Clostridium perfringens toxins in human and animal disease. Future Microbiol. 2014, 9, 361–377. [Google Scholar] [CrossRef] [Green Version]
- Uzal, F.A.; Navarro, M.A.; Li, J.J.; Freedman, J.C.; Shrestha, A.; McClane, B.A. Comparative pathogenesis of enteric clostridial infections in humans and animals. Anaerobe 2018, 53, 11–20. [Google Scholar] [CrossRef]
- Sayeed, S.; Uzal, F.A.; Fisher, D.J.; Saputo, J.; Vidal, J.E.; Chen, Y.; Gupta, P.; Rood, J.I.; McClane, B.A. Beta toxin is essential for the intestinal virulence of Clostridium perfringens type C disease isolate CN3685 in a rabbit ileal loop model. Mol. Microbiol. 2008, 67, 15–30. [Google Scholar] [CrossRef]
- Ghoneim, N.H.; Hamza, D.A. Epidemiological studies on Clostridium perfringens food poisoning in retail foods. Rev. Sci. Tech. Off. Int. Epiz 2017, 36, 1025–1032. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, A.; Uzal, F.A.; McClane, B.A. Enterotoxic Clostridia: Clostridium perfringens Enteric Diseases. Gram-Positive Pathog. 2018, 6, 977–990. [Google Scholar] [CrossRef]
- Miyamoto, K.; Li, J.; McClane, B.A. Enterotoxigenic Clostridium perfringens: Detection and identification. Microbes Environ. 2012, 27, 343–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villarruel-López, A.; Ruíz-Quezada, S.L.; Castro-Rosas, J.; Gomez-Aldapa, C.A.; Olea-Rodríguez, M.A.; Nuño, K.; Navarro-Hidalgo, V.; Torres-Vitela, M.R. Behavior and inactivation of enterotoxin-positive Clostridium perfringens in pork picadillo and tamales filled with pork picadillo under different cooking, storage, and reheating conditions. J. Food Prot. 2016, 79, 741–747. [Google Scholar] [CrossRef] [PubMed]
- DuPont, H.L. Bacterial Diarrhea. N. Engl. J. Med. 2009, 361, 1560–1569. [Google Scholar] [CrossRef]
- Wong, S.; Santullo, P.; O’driscoll, J.; Jamous, A.; Hirani, S.P.; Saif, M. Use of antibiotic and prevalence of antibiotic-associated diarrhoea in-patients with spinal cord injuries: A UK national spinal injury centre experience. Spinal Cord 2017, 55, 583–587. [Google Scholar] [CrossRef] [Green Version]
- Gui, L.; Subramony, C.; Fratkin, J.; Hughson, M.D. Fatal enteritis necroticans (Pigbel) in a diabetic adult. Mod. Pathol. 2002, 15, 66–70. [Google Scholar] [CrossRef] [Green Version]
- Stevens, D.L.; Bryant, A.E. Necrotizing soft-tissue infections. N. Engl. J. Med. 2017, 377, 2253–2265. [Google Scholar] [CrossRef]
- Zúñiga-Pereira, A.M.; Santamaría, C.; Gutierrez, J.M.; Alape-Girón, A.; Flores-Díaz, M. Deficient skeletal muscle regeneration after injury induced by a Clostridium perfringens strain associated with gas gangrene. Infect. Immun. 2019, 87, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Zahoor, I.; Ghayas, A.; Basheer, A. Genetics and genomics of susceptibility and immune response to necrotic enteritis in chicken: A review. Mol. Biol. Rep. 2018, 45, 31–37. [Google Scholar] [CrossRef]
- Annett, C.B.; Viste, J.R.; Chirino-Trejo, M.; Classen, H.L.; Middleton, D.M.; Simko, E. Necrotic enteritis: Effect of barley, wheat and corn diets on proliferation of Clostridium perfringens type A. Avian Pathol. 2002, 31, 598–601. [Google Scholar] [CrossRef] [PubMed]
- Mejia, D.B.; Peñuela, -S.L.M.; Sanmiguel, R.A. El gran impacto de Clostridium perfringens en aves de corral. Pubvet 2018, 12, 1–9. [Google Scholar] [CrossRef]
- Van Immerseel, F.; De Buck, J.; Pasmans, F.; Huyghebaert, G.; Haesebrouck, F.; Ducatelle, R. Clostridium perfringens in poultry: An emerging threat for animal and public health. Avian Pathol. 2004, 33, 537–549. [Google Scholar] [CrossRef] [PubMed]
- Gharib-Naseri, K.; Kheravii, S.K.; Keerqin, C.; Morgan, N.; Swick, R.A.; Choct, M.; Wu, S.B. Two different Clostridium perfringens strains produce different levels of necrotic enteritis in broiler chickens. Poult. Sci. 2019, 98, 6422–6432. [Google Scholar] [CrossRef] [PubMed]
- Timbermont, L.; Haesebrouck, F.; Ducatelle, R.; Van Immerseel, F. Necrotic enteritis in broilers: An updated review on the pathogenesis. Avian Pathol. 2011, 40, 341–347. [Google Scholar] [CrossRef]
- Timbermont, L.; Lanckriet, A.; Dewulf, J.; Nollet, N.; Schwarzer, K.; Haesebrouck, F.; Ducatelle, R.; van Immerseel, F. Control of clostridium perfringens-induced necrotic enteritis in broilers by target-released butyric acid, fatty acids and essential oils. Avian Pathol. 2010, 39, 117–121. [Google Scholar] [CrossRef] [Green Version]
- Rood, J.I.; Keyburn, A.L.; Moore, R.J. NetB and necrotic enteritis: The hole movable story. Avian Pathol. 2016, 45, 295–301. [Google Scholar] [CrossRef] [Green Version]
- Van Immerseel, F.; Rood, J.I.; Moore, R.J.; Titball, R.W. Rethinking our understanding of the pathogenesis of necrotic enteritis in chickens. Trends Microbiol. 2009, 17, 32–36. [Google Scholar] [CrossRef] [Green Version]
- Moore, R.J. Necrotic enteritis predisposing factors in broiler chickens. Avian Pathol. 2016, 45, 275–281. [Google Scholar] [CrossRef] [Green Version]
- Timbermont, L.; De Smet, L.; Van Nieuwerburgh, F.; Parreira, V.R.; Van Driessche, G.; Haesebrouck, F.; Ducatelle, R.; Prescott, J.; Deforce, D.; Devreese, B.; et al. Perfrin, a novel bacteriocin associated with netB positive Clostridium perfringens strains from broilers with necrotic enteritis. Vet. Res. 2014, 45, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Flores-Díaz, M.; Barquero-Calvo, E.; Ramírez, M.; Alape-Girón, A. Role of Clostridium perfringens Toxins in Necrotic Enteritis in Poultry. Microb. Toxins 2016, 1–16. [Google Scholar] [CrossRef]
- Prescott, J.F.; Parreira, V.R.; Mehdizadeh Gohari, I.; Lepp, D.; Gong, J. The pathogenesis of necrotic enteritis in chickens: What we know and what we need to know: A review. Avian Pathol. 2016, 45, 288–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Razmyar, J.; Peighambari, S.M.; Zamani, A.H. Detection of a Newly Described Bacteriocin, Perfrin, among Clostridium perfringens Isolates from Healthy and Diseased Ostriches and Broiler Chickens in Iran. Avian Dis. 2017, 61, 387–390. [Google Scholar] [CrossRef] [PubMed]
- Wade, B.; Keyburn, A.L.; Seemann, T.; Rood, J.I.; Moore, R.J. Binding of Clostridium perfringens to collagen correlates with the ability to cause necrotic enteritis in chickens. Vet. Microbiol. 2015, 180, 299–303. [Google Scholar] [CrossRef] [PubMed]
- Cheung, J.K.; Keyburn, A.L.; Carter, G.P.; Lanckriet, A.L.; Van Immerseel, F.; Moore, R.J.; Rood, J.I. The VirSR two-component signal transduction system regulates NetB toxin production in Clostridium perfringens. Infect. Immun. 2010, 78, 3064–3072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Q.; Lepp, D.; Gohari, I.M.; Wu, T.; Zhou, H.; Yin, X.; Yu, H.; Prescott, J.F.; Nie, S.P.; Xie, M.Y.; et al. The Agr-like quorum sensing system is required for pathogenesis of necrotic enteritis caused by Clostridium perfringens in poultry. Infect. Immun. 2017, 85, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.H. Sialidases From Clostridium perfringens and Their Inhibitors. Front. Cell. Infect. Microbiol. 2020, 9, 1–11. [Google Scholar] [CrossRef]
- Lepp, D.; Roxas, B.; Parreira, V.R.; Marri, P.R.; Rosey, E.L.; Gong, J.; Songer, J.G.; Vedantam, G.; Prescott, J.F. Identification of novel pathogenicity loci in Clostridium perfringens strains that cause Avian necrotic enteritis. PLoS ONE 2010, 5. [Google Scholar] [CrossRef]
- Savva, C.G.; Da Costa, S.P.F.; Bokori-Brown, M.; Naylor, C.E.; Cole, A.R.; Moss, D.S.; Titball, R.W.; Basak, A.K. Molecular architecture and functional analysis of NetB, a pore-forming toxin from Clostridium perfringens. J. Biol. Chem. 2013, 288, 3512–3522. [Google Scholar] [CrossRef] [Green Version]
- Popoff, M.R.; Bouvet, P. Clostridial toxins. Future Microbiol. 2009, 4, 1021–1064. [Google Scholar] [CrossRef]
- Navarro, M.A.; McClane, B.A.; Uzal, F.A. Mechanisms of action and cell death associated with Clostridium perfringens toxins. Toxins 2018, 10, 212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paiva, D.; McElroy, A. Necrotic enteritis: Applications for the poultry industry. J. Appl. Poult. Res. 2014, 23, 557–566. [Google Scholar] [CrossRef]
- Edelblum, K.L.; Turner, J.R. The Tight Junction in Inflammatory Disease: Communication Breakdown. Curr. Opin. Pharmacol. 2009, 9, 715–720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Llanco, L.A.; Nakano, V.; Moraes, C.T.P.d.; Piazza, R.M.F.; Avila-Campos, M.J. Adhesion and invasion of Clostridium perfringens type A into epithelial cells. Brazilian J. Microbiol. 2017, 48, 764–768. [Google Scholar] [CrossRef] [PubMed]
- Coursodon, C.F.; Glock, R.D.; Moore, K.L.; Cooper, K.K.; Songer, J.G. TpeL-producing strains of Clostridium perfringens type A are highly virulent for broiler chicks. Anaerobe 2012, 18, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Schorch, B.; Song, S.; Van Diemen, F.R.; Bock, H.H.; May, P.; Herz, J.; Brummelkamp, T.R.; Papatheodorou, P.; Aktories, K. LRP1 is a receptor for Clostridium perfringens TpeL toxin indicating a two-receptor model of clostridial glycosylating toxins. Proc. Natl. Acad. Sci. USA 2014, 111, 6431–6436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alshamy, Z.; Richardson, K.C.; Hünigen, H.; Hafez, H.M.; Plendl, J.; Al Masri, S. Comparison of the gastrointestinal tract of a dual-purpose to a broiler chicken line: A qualitative and quantitative macroscopic and microscopic study. PLoS ONE 2018, 13, e0204921. [Google Scholar] [CrossRef] [Green Version]
- Laledashti, M.A.; Saki, A.A.; Rafati, A.A.; Abdolmaleki, M. Effect of in-ovo feeding of iron nanoparticles and methionine hydroxy analogue on broilers chickens small intestinal characteristics. Acta Sci. Anim. Sci. 2020, 42. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Jia, Z. Microbiome modulates intestinal homeostasis against inflammatory diseases. Vet. Immunol. Immunopathol. 2018, 205, 97–105. [Google Scholar] [CrossRef]
- Cooper, K.K.; Songer, J.G.; Uzal, F.A. Diagnosing clostridial enteric disease in poultry. J. Vet. Diagnostic Investig. 2013, 25, 314–327. [Google Scholar] [CrossRef] [Green Version]
- Redondo, L.M.; Redondo, E.A.; Delgado, F.; La Sala, L.F.; Fernández Miyakawa, M.E. An Experimental Reproduction of Necrotic Enteritis in Broiler Chickens. J. Vet. Sci. Med. 2016, 4, 1–5. [Google Scholar] [CrossRef]
- Smyth, J.A. Pathology and diagnosis of necrotic enteritis: Is it clear-cut? Avian Pathol. 2016, 45, 282–287. [Google Scholar] [CrossRef] [PubMed]
- Awad, W.A.; Hess, C.; Hess, M. Enteric pathogens and their toxin-induced disruption of the intestinal barrier through alteration of tight junctions in chickens. Toxins 2017, 9, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Meyer, F.; Eeckhaut, V.; Ducatelle, R.; Dhaenens, M.; Daled, S.; Dedeurwaerder, A.; De Gussem, M.; Haesebrouck, F.; Deforce, D.; Van Immerseel, F. Host intestinal biomarker identification in a gut leakage model in broilers. Vet. Res. 2019, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez-Lopez, A.; Behnsen, J.; Nuccio, S.P.; Raffatellu, M. Mucosal immunity to pathogenic intestinal bacteria. Nat. Rev. Immunol. 2016, 16, 135–148. [Google Scholar] [CrossRef] [PubMed]
- Gholamiandehkordi, A.R.; Timbermont, L.; Lanckriet, A.; Van Den Broeck, W.; Pedersen, K.; Dewulf, J.; Pasmans, F.; Haesebrouck, F.; Ducatelle, R.; Van Immerseel, F. Quantification of gut lesions in a subclinical necrotic enteritis model. Avian Pathol. 2007, 36, 375–382. [Google Scholar] [CrossRef] [Green Version]
- Goossens, E.; Valgaeren, B.R.; Pardon, B.; Haesebrouck, F.; Ducatelle, R.; Deprez, P.R.; Van Immerseel, F. Rethinking the role of alpha toxin in Clostridium perfringens-associated enteric diseases: A review on bovine necro-haemorrhagic enteritis. Vet. Res. 2017, 48, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Sanchez, L.C. Disorders of the Gastrointestinal System. Equine Intern. Med. 2018, 709–842. [Google Scholar] [CrossRef]
- Clavijo, V.; Flórez, M.J.V. The gastrointestinal microbiome and its association with the control of pathogens in broiler chicken production: A review. Poult. Sci. 2018, 97, 1006–1021. [Google Scholar] [CrossRef]
- Lacey, J.A.; Stanley, D.; Keyburn, A.L.; Ford, M.; Chen, H.; Johanesen, P.; Lyras, D.; Moore, R.J. Clostridium perfringens-mediated necrotic enteritis is not influenced by the pre-existing microbiota but is promoted by large changes in the post-challenge microbiota. Vet. Microbiol. 2018, 227, 119–126. [Google Scholar] [CrossRef]
- Villagran-de la Mora, Z.; Nuño, K.; Olga, V.; Avalos, H.; Castro-rosas, J.; Carlos, G.; Angulo, C.; Ascencio, F. Effect of a Synbiotic Mix on Intestinal Structural Changes, and Salmonella Typhimurium and Clostridium Perfringens Colonization in Broiler Chickens. Animals 2019, 9, 777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keyburn, A.L.; Sheedy, S.A.; Ford, M.E.; Williamson, M.M.; Awad, M.M.; Rood, J.I.; Moore, R.J. Alpha-toxin of Clostridium perfringens is not an essential virulence factor in necrotic enteritis in chickens. Infect. Immun. 2006, 74, 6496–6500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shojadoost, B.; Vince, A.R.; Prescott, J.F. The successful experimental induction of necrotic enteritis in chickens by Clostridium perfringens: A critical review. Vet. Res. 2012, 43, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chuang, W.Y.; Lin, W.C.; Hsieh, Y.C.; Huang, C.M.; Chang, S.C.; Lee, T.T. Evaluation of the combined use of Saccharomyces cerevisiae and Aspergillus oryzae with phytase fermentation products on growth, inflammatory, and intestinal morphology in broilers. Animals 2019, 9, 1051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.W.; Lillehoj, H.S.; Jeong, W.; Jeoung, H.Y.; An, D.J. Avian necrotic enteritis: Experimental models, host immunity, pathogenesis, risk factors, and vaccine development. Poult. Sci. 2011, 90, 1381–1390. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Kim, W.H.; Lee, S.J.; Lillehoj, H.S. Detection of chicken interleukin-10 production in intestinal epithelial cells and necrotic enteritis induced by Clostridium perfringens using capture ELISA. Vet. Immunol. Immunopathol. 2018, 204, 52–58. [Google Scholar] [CrossRef]
- Wise, M.G.; Siragusa, G.R. Quantitative detection of Clostridium perfringens in the broiler fowl gastrointestinal tract by real-time PCR. Appl. Environ. Microbiol. 2005, 71, 3911–3916. [Google Scholar] [CrossRef] [Green Version]
- Bailey, M.A.; Macklin, K.S.; Krehling, J.T. Use of a Multiplex PCR for the Detection of Toxin-Encoding Genes netB and tpeL in Strains of Clostridium perfringens. ISRN Vet. Sci. 2013, 2013, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Food and Agriculture Organization, (FAO); World Health Organization, (WHO). Probiotics in Food Health and Nutritional Properties and Guidelines for Evaluation; FAO, WHO, Eds.; FAO WHO: Rome, Italy, 2006; Volume 85, ISBN 9251055130. [Google Scholar]
- Jin, L.Z.; Ho, Y.W.; Abdullah, N.; Jalaludin, S. Probiotics in poultry: Modes of action. Worlds. Poult. Sci. J. 1997, 53, 362–368. [Google Scholar] [CrossRef]
- Park, Y.H.; Hamidon, F.; Rajangan, C.; Soh, K.P.; Gan, C.Y.; Lim, T.S.; Abdullah, W.N.W.; Liong, M.T. Application of probiotics for the production of safe and high-quality poultry meat. Korean J. Food Sci. Anim. Resour. 2016, 36, 567–576. [Google Scholar] [CrossRef] [Green Version]
- Ma, T.; Suzuki, Y.; Guan, L.L. Dissect the mode of action of probiotics in affecting host-microbial interactions and immunity in food producing animals. Vet. Immunol. Immunopathol. 2018, 205, 35–48. [Google Scholar] [CrossRef] [PubMed]
- Azad, M.A.K.; Sarker, M.; Li, T.; Yin, J. Probiotic Species in the Modulation of Gut Microbiota: An Overview. Biomed Res. Int. 2018, 2018. [Google Scholar] [CrossRef] [Green Version]
- Popova, T. Effect of probiotics in poultry for improving meat quality. Curr. Opin. Food Sci. 2017, 14, 72–77. [Google Scholar] [CrossRef]
- Carlander, D.; Stålberg, J.; Larsson, A. Chicken antibodies: A clinical chemistry perspective. Ups. J. Med. Sci. 1999, 104, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Khalique, A.; Zeng, D.; Shoaib, M.; Wang, H.; Qing, X.; Rajput, D.S.; Pan, K.; Ni, X. Probiotics mitigating subclinical necrotic enteritis (SNE) as potential alternatives to antibiotics in poultry. AMB Express 2020, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menconi, A.; Sokale, A.O.; Mendoza, S.M.; Whelan, R.; Doranalli, K. Effect of Bacillus subtilis DSM 32315 under different Necrotic Enteritis models in broiler chickens: A meta-analysis of 5 independent research trials. Avian Dis. 2020. [Google Scholar] [CrossRef]
- Johnson, T.J.; Youmans, B.P.; Noll, S.; Cardona, C.; Evans, N.P.; Peter Karnezos, T.; Ngunjiri, J.M.; Abundo, M.C.; Lee, C.W. A consistent and predictable commercial broiler chicken bacterial microbiota in antibiotic-free production displays strong correlations with performance. Appl. Environ. Microbiol. 2018, 84, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Stanley, D.; Hughes, R.J.; Geier, M.S.; Moore, R.J. Bacteria within the gastrointestinal tract microbiota correlated with improved growth and feed conversion: Challenges presented for the identification of performance enhancing probiotic bacteria. Front. Microbiol. 2016, 7, 1–13. [Google Scholar] [CrossRef] [Green Version]
- La Ragione, R.M.; Narbad, A.; Gasson, M.J.; Woodward, M.J. In vivo characterization of Lactobacillus johnsonii FI9785 for use as a defined competitive exclusion agent against bacterial pathogens in poultry. Lett. Appl. Microbiol. 2004, 38, 197–205. [Google Scholar] [CrossRef]
- Gervasi, T.; Lo Curto, R.; Minniti, E.; Narbad, A.; Mayer, M.J. Application of Lactobacillus johnsonii expressing phage endolysin for control of Clostridium perfringens. Lett. Appl. Microbiol. 2014, 59, 355–361. [Google Scholar] [CrossRef]
- Kim, C.H.; Shin, K.S.; Woo, K.C.; Paik, I.K. Effect of Dietary Oligosaccharides on the Performance, Intestinal Microflora and Serum Immunoglobulin Contents in Laying Hens. Korean J. Poult. Sci. 2009, 36, 125–131. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Shao, Y.; Song, B.; Zhen, W.; Wang, Z.; Guo, Y.; Shahid, M.S.; Nie, W. Effects of Bacillus coagulans supplementation on the growth performance and gut health of broiler chickens with Clostridium perfringens-induced necrotic enteritis. J. Anim. Sci. Biotechnol. 2018, 9, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Lin, Y.; Zeng, D.; Zhou, M.; Zeng, Y.; Wang, H.; Zhou, Y.; Zhu, H.; Pan, K.; Jing, B.; et al. Bacillus licheniformis normalize the ileum microbiota of chickens infected with necrotic enteritis. Sci. Rep. 2018, 8, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zeng, D.; Wang, H.; Qing, X.; Sun, N.; Xin, J.; Luo, M.; Khalique, A.; Pan, K.; Shu, G.; et al. Dietary Probiotic Bacillus licheniformis H2 Enhanced Growth Performance, Morphology of Small Intestine and Liver, and Antioxidant Capacity of Broiler Chickens Against Clostridium perfringens–Induced Subclinical Necrotic Enteritis. Probiotics Antimicrob. Proteins 2019. [Google Scholar] [CrossRef] [PubMed]
- Aljumaah, M.R.; Alkhulaifi, M.M.; Abudabos, A.M.; Aljumaah, R.S.; Alsaleh, A.N.; Stanley, D. Bacillus subtilis PB6 based probiotic supplementation plays a role in the recovery after the necrotic enteritis challenge. PLoS ONE 2020, 15, e0232781. [Google Scholar] [CrossRef] [PubMed]
- Ramlucken, U.; Ramchuran, S.O.; Moonsamy, G.; Lalloo, R.; Thantsha, M.S.; Jansen van Rensburg, C. A novel Bacillus based multi-strain probiotic improves growth performance and intestinal properties of Clostridium perfringens challenged broilers. Poult. Sci. 2020, 99, 331–341. [Google Scholar] [CrossRef]
- Wang, H.; Ni, X.; Qing, X.; Liu, L.; Xin, J.; Luo, M.; Khalique, A.; Dan, Y.; Pan, K.; Jing, B.; et al. Probiotic lactobacillus johnsonii BS15 improves blood parameters related to immunity in broilers experimentally infected with subclinical necrotic enteritis. Front. Microbiol. 2018, 9, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Cao, L.; Wu, X.H.; Bai, Y.L.; Wu, X.Y.; Gu, S.B. Anti-inflammatory and antioxidant activities of probiotic powder containing Lactobacillus plantarum 1.2567 in necrotic enteritis model of broiler chickens. Livest. Sci. 2019, 223, 157–163. [Google Scholar] [CrossRef]
- Cao, L.; Yang, X.J.; Li, Z.J.; Sun, F.F.; Wu, X.H.; Yao, J.H. Reduced lesions in chickens with Clostridium perfringens-induced necrotic enteritis by Lactobacillus fermentum 1.2029. Poult. Sci. 2012, 91, 3065–3071. [Google Scholar] [CrossRef]
- Guo, S.; Liu, D.; Zhang, B.; Li, Z.; Li, Y.; Ding, B.; Guo, Y. Two Lactobacillus species inhibit the growth and α-toxin production of Clostridium perfringens and induced proinflammatory factors in chicken intestinal epithelial cells in vitro. Front. Microbiol. 2017, 8, 1–12. [Google Scholar] [CrossRef]
- Xu, T.; Chen, Y.; Yu, L.; Wang, J.; Huang, M.; Zhu, N. Effects of Lactobacillus plantarum on intestinal integrity and immune responses of egg-laying chickens infected with Clostridium perfringens under the free-range or the specific pathogen free environment. BMC Vet. Res. 2020, 16, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.; Zhen, W.; Geng, Y.; Wang, Z.; Guo, Y. Pretreatment with probiotic Enterococcus faecium NCIMB 11181 ameliorates necrotic enteritis-induced intestinal barrier injury in broiler chickens. Sci. Rep. 2019, 9, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.R.; Dong, X.F.; Liu, S.; Tong, J.M. Effects of long-term Bacillus subtilis CGMCC 1.921 supplementation on performance, egg quality, and fecal and cecal microbiota of laying hens. Poult. Sci. 2017, 96, 1280–1289. [Google Scholar] [CrossRef] [PubMed]
- Jayaraman, S.; Thangavel, G.; Kurian, H.; Mani, R.; Mukkalil, R.; Chirakkal, H. Bacillus subtilis PB6 improves intestinal health of broiler chickens challenged with Clostridium perfringens-induced necrotic enteritis. Poult. Sci. 2013, 92, 370–374. [Google Scholar] [CrossRef]
- Forte, C.; Acuti, G.; Manuali, E.; Casagrande Proietti, P.; Pavone, S.; Trabalza-Marinucci, M.; Moscati, L.; Onofri, A.; Lorenzetti, C.; Franciosini, M.P. Effects of two different probiotics on microflora, morphology, and morphometry of gut in organic laying hens. Poult. Sci. 2016, 95, 2528–2535. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.M.; Begum, M.; Kim, I.H. Effect of Bacillus subtilis, Clostridium butyricum and Lactobacillus acidophilus endospores on growth performance, nutrient digestibility, meat quality, relative organ weight, microbial shedding and excreta noxious gas emission in broilers. Vet. Med. 2015, 60, 77–86. [Google Scholar] [CrossRef] [Green Version]
- Olnood, C.G.; Beski, S.S.M.; Iji, P.A.; Choct, M. Delivery routes for probiotics: Effects on broiler performance, intestinal morphology and gut microflora. Anim. Nutr. 2015, 1, 192–202. [Google Scholar] [CrossRef]
- Rantala, M.; Nurmi, E. Prevention of the growth of salmonella infantis in chicks by the flora of the alimentary tract of chickens. Br. Poult. Sci. 1973, 14, 627–630. [Google Scholar] [CrossRef]
- Caly, D.L.; D’Inca, R.; Auclair, E.; Drider, D. Alternatives to antibiotics to prevent necrotic enteritis in broiler chickens: A microbiologist’s perspective. Front. Microbiol. 2015, 6, 1336. [Google Scholar] [CrossRef] [Green Version]
- Craven, A.S.E.; Stern, N.J.; Cox, N.A.; Bailey, J.S.; Berrang, M.; Craven, S.E.; Stern, N.J.; Cox, N.A.; Bailey, J.S.; Berrang, M. Cecal Carriage of Clostridium perfringens in Broiler Chickens Given Mucosal Starter Culture. Avian Dis. 2018, 43, 484–490. [Google Scholar] [CrossRef]
- Rubio, L.A. Possibilities of early life programming in broiler chickens via intestinal microbiota modulation. Poult. Sci. 2019, 98, 695–706. [Google Scholar] [CrossRef] [PubMed]
- Ghareeb, K.; Awad, W.A.; Mohnl, M.; Porta, R.; Biarnés, M.; Böhm, J.; Schatzmayr, G. Evaluating the efficacy of an avian-specific probiotic to reduce the colonization of Campylobacter jejuni in broiler chickens. Poult. Sci. 2012, 91, 1825–1832. [Google Scholar] [CrossRef] [PubMed]
- Froebel, L.K.; Jalukar, S.; Lavergne, T.A.; Lee, J.T.; Duong, T. Administration of dietary prebiotics improves growth performance and reduces pathogen colonization in broiler chickens. Poult. Sci. 2019, 98, 6668–6676. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.A.; Jang, M.J.; Kim, S.Y.; Yang, Y.; Pavlidis, H.O.; Ricke, S.C. Potential for prebiotics as feed additives to limit foodborne Campylobacter establishment in the poultry gastrointestinal tract. Front. Microbiol. 2019, 10, 1–12. [Google Scholar] [CrossRef]
- Bucław, M. Inulin in poultry production. Worlds. Poult. Sci. J. 2017, 73, 301–308. [Google Scholar] [CrossRef]
- Teng, P.; Kim, W.K. Review: Roles of Prebiotics in Intestinal Ecosystem of Broilers. Front. Vet. Sci. 2018, 5, 1–18. [Google Scholar] [CrossRef]
- Ghasemi, H.A.; Shivazad, M.; Mirzapour Rezaei, S.S.; Torshizi, M.A.K. Effect of synbiotic supplementation and dietary fat sources on broiler performance, serum lipids, muscle fatty acid profile and meat quality. Br. Poult. Sci. 2015, 57. [Google Scholar] [CrossRef]
- Huyghebaert, G.; Ducatelle, R.; Van Immerseel, F. Van An update on alternatives to antimicrobial growth promoters for broilers. Vet. J. 2011, 187, 182–188. [Google Scholar] [CrossRef] [Green Version]
- Hashim, M.M.; Arsenault, R.J.; Byrd, J.A.; Kogut, M.H.; Al-Ajeeli, M.; Bailey, C.A. Influence of different yeast cell wall preparations and their components on performance and immune and metabolic pathways in Clostridium perfringens-challenged broiler chicks. Poult. Sci. 2018, 97, 203–210. [Google Scholar] [CrossRef]
- Guaragni, A.; Boiago, M.M.; Bottari, N.B.; Morsch, V.M.; Lopes, T.F.; Schafer da Silva, A. Feed supplementation with inulin on broiler performance and meat quality challenged with Clostridium perfringens: Infection and prebiotic impacts. Microb. Pathog. 2020, 139, 103889. [Google Scholar] [CrossRef]
- Ricke, S.C.; Lee, S.I.; Kim, S.A.; Park, S.H.; Shi, Z. Prebiotics and the poultry gastrointestinal tract microbiome. Poult. Sci. 2020, 99, 670–677. [Google Scholar] [CrossRef] [PubMed]
- Ajuwon, K.M. Toward a better understanding of mechanisms of probiotics and prebiotics action in poultry species. J. Appl. Poult. Res. 2016, 25, 277–283. [Google Scholar] [CrossRef]
- Kheravii, S.K.; Swick, R.A.; Choct, M.; Wu, S.B. Effect of oat hulls as a free choice feeding on broiler performance, short chain fatty acids and microflora under a mild necrotic enteritis challenge. Anim. Nutr. 2018, 4, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Mtei, A.W.; Abdollahi, M.R.; Schreurs, N.; Girish, C.K.; Ravindran, V. Dietary inclusion of fibrous ingredients and bird type influence apparent ileal digestibility of nutrients and energy utilization. Poult. Sci. 2019, 98, 6702–6712. [Google Scholar] [CrossRef] [PubMed]
- Gibson, G.R.; Roberfroid, M.B. Dietary modulation of the human colonic microbiota: Introducing the concept of prebiotics. J. Nutr. 1995, 125, 1401–1412. [Google Scholar] [CrossRef]
- Peña, A.S. Flora intestinal, probióticos, prebióticos, simbióticos y alimentos novedosos. Rev. Esp. Enferm. Dig. 2007, 99, 653–658. [Google Scholar] [CrossRef] [Green Version]
- Pandey, K.R.; Naik, S.R.; Vakil, B.V. Probiotics, prebiotics and synbiotics—A review. J. Food Sci. Technol. 2015, 52, 7577–7587. [Google Scholar] [CrossRef]
- Gadde, U.; Kim, W.H.; Oh, S.T.; Lillehoj, H.S. Alternatives to antibiotics for maximizing growth performance and feed efficiency in poultry: A review. Anim. Heal. Res. Rev. 2017, 18, 26–45. [Google Scholar] [CrossRef]
- Markowiak, P.; Ślizewska, K. The role of probiotics, prebiotics and synbiotics in animal nutrition. Gut Pathog. 2018, 10, 1–20. [Google Scholar] [CrossRef]
- Baffoni, L.; Gaggìa, F.; Garofolo, G.; Di Serafino, G.; Buglione, E.; Di Giannatale, E.; Di Gioia, D. Evidence of Campylobacter jejuni reduction in broilers with early synbiotic administration. Int. J. Food Microbiol. 2017, 251, 41–47. [Google Scholar] [CrossRef]
- Koc, F.; Samli, H.; Okur, A.; Ozduven, M.; Akyurek, H.; Senkoylu, N. Effects of Saccharomyces cerevisiae and/or mannanoligosaccharide on performance, blood parameters and intestinal microbiota of broiler chicks. Bulg. J. Agric. Sci. 2010, 16, 643–650. [Google Scholar]
- Mookiah, S.; Sieo, C.C.; Ramasamy, K.; Abdullah, N.; Ho, Y.W. Effects of dietary prebiotics, probiotic and synbiotics on performance, caecal bacterial populations and caecal fermentation concentrations of broiler chickens. J. Sci. Food Agric. 2014, 94, 341–348. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Ghany, W.A. Comparative evaluation on the effect of coccidiostate and synbiotic preparations on prevention of Clostridium perfringens in broiler chickens. Glob. Vet. 2010, 5, 324–333. [Google Scholar]
- Krueger, L.A.; Spangler, D.A.; Vandermyde, D.R.; Sims, M.D.; Ayangbile, G.A. Avi-Lution® supplemented at 1.0 or 2.0 g/kg in feed improves the growth performance of broiler chickens during challenge with bacitracin-resistant Clostridium perfringens. Poult. Sci. 2017, 96, 2595–2600. [Google Scholar] [CrossRef] [PubMed]
- Shanmugasundaram, R.; Markazi, A.; Mortada, M.; Ng, T.T.; Applegate, T.J.; Bielke, L.R.; Syed, B.; Pender, C.M.; Curry, S.; Murugesan, G.R.; et al. Research Note: Effect of synbiotic supplementation on caecal Clostridium perfringens load in broiler chickens with different necrotic enteritis challenge models. Poult. Sci. 2020, 99, 2452–2458. [Google Scholar] [CrossRef]
- Yang, C.; Chowdhury, M.A.K.; Hou, Y.; Gong, J. Phytogenic compounds as alternatives to in-feed antibiotics: Potentials and challenges in application. Pathogens 2015, 4, 137–156. [Google Scholar] [CrossRef] [Green Version]
- Pham, V.H.; Kan, L.; Huang, J.; Geng, Y.; Zhen, W.; Guo, Y.; Abbas, W.; Wang, Z. Dietary encapsulated essential oils and organic acids mixture improves gut health in broiler chickens challenged with necrotic enteritis. J. Anim. Sci. Biotechnol. 2020, 11, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Granstad, S.; Kristoffersen, A.B.; Benestad, S.L.; Sjurseth, S.K.; David, B.; Sørensen, L.; Fjermedal, A.; Edvardsen, D.H.; Sanson, G.; Løvland, A.; et al. Effect of feed additives as alternatives to in-feed antimicrobials on production performance and intestinal Clostridium perfringens counts in broiler chickens. Animals 2020, 10, 240. [Google Scholar] [CrossRef] [Green Version]
- Abudabos, A.M. Use of a competitive exclusion product (Aviguard) to prevent Clostridium perfringens colonization in broiler chicken under induced challenge. Pak. J. Zool. 2013, 45, 371–376. [Google Scholar]
- Hussein, E.O.S.; Ahmed, S.H.; Abudabos, A.M.; Aljumaah, M.R.; Alkhlulaifi, M.M.; Nassan, M.A.; Suliman, G.M.; Naiel, M.A.E.; Swelum, A.A. Effect of antibiotic, phytobiotic and probiotic supplementation on growth, blood indices and intestine health in broiler chicks challenged with Clostridium perfringens. Animals 2020, 10, 507. [Google Scholar] [CrossRef] [Green Version]
- Tzora, A.; Giannenas, I.; Karamoutsios, A.; Papaioannou, N.; Papanastasiou, D.; Bonos, E.; Skoufos, S.; Bartzanas, T.; Skoufos, I. Effects of oregano, attapulgite, benzoic acid and their blend on chicken performance, intestinal microbiology and intestinal morphology. J. Poult. Sci. 2017, 54, 218–227. [Google Scholar] [CrossRef] [PubMed]
- Kirkpinar, F.; Ünlü, H.B.; Özdemir, G. Effects of oregano and garlic essential oils on performance, carcase, organ and blood characteristics and intestinal microflora of broilers. Livest. Sci. 2011, 137, 219–225. [Google Scholar] [CrossRef]
- Du, E.; Wang, W.; Gan, L.; Li, Z.; Guo, S.; Guo, Y. Effects of thymol and carvacrol supplementation on intestinal integrity and immune responses of broiler chickens challenged with Clostridium perfringens. J. Anim. Sci. Biotechnol. 2016, 7, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdelli, N.; Pérez, J.F.; Vilarrasa, E.; Luna, I.C.; Melo-Duran, D.; D’angelo, M.; Solà-Oriol, D. Targeted-release organic acids and essential oils improve performance and digestive function in broilers under a necrotic enteritis challenge. Animals 2020, 10, 259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Q.; Liu, D.; Guo, S.; Chen, Y.; Guo, Y. Effects of dietary essential oil and enzyme supplementation on growth performance and gut health of broilers challenged by Clostridium perfringens. Anim. Feed Sci. Technol. 2015, 207, 234–244. [Google Scholar] [CrossRef]
- Mannelli, F.; Minieri, S.; Tosi, G.; Secci, G.; Daghio, M.; Massi, P.; Fiorentini, L.; Galigani, I.; Lancini, S.; Rapaccini, S.; et al. Effect of chestnut tannins and short chain fatty acids as anti-microbials and as feeding supplements in broilers rearing and meat quality. Animals 2019, 9, 659. [Google Scholar] [CrossRef] [Green Version]
- Dahiya, J.P.; Wilkie, D.C.; Van Kessel, A.G.; Drew, M.D. Potential strategies for controlling necrotic enteritis in broiler chickens in post-antibiotic era. Anim. Feed Sci. Technol. 2006, 129, 60–88. [Google Scholar] [CrossRef]
- Lensing, M.; van der Klis, J.D.; Fabri, T.; Cazemier, A.; Else, A.J. Efficacy of a lactylate on production performance and intestinal health of broilers during a subclinical Clostridium perfringens infection. Poult. Sci. 2010, 89, 2401–2409. [Google Scholar] [CrossRef]
- Drew, M.D.; Syed, N.A.; Goldade, B.G.; Laarveld, B.; Van Kessel, A.G. Effects of dietary protein source and level on intestinal populations of Clostridium perfringens in broiler chickens. Poult. Sci. 2004, 83, 414–420. [Google Scholar] [CrossRef]
- Kumar, S.; Adhikari, P.; Oakley, B.; Kim, W.K. Changes in cecum microbial community in response to total sulfur amino acid (TSAA: DL-methionine) in antibiotic-free and supplemented poultry birds. Poult. Sci. 2019, 98, 5809–5819. [Google Scholar] [CrossRef]
- Sieo, C.C.; Abdullah, N.; Tan, W.S.; Ho, Y.W. Influence of β-glucanase-producing lactobacillus strains on intestinal characteristics and feed passage rate of broiler chickens. Poult. Sci. 2005, 84, 734–741. [Google Scholar] [CrossRef]
- Tsiouris, V.; Georgopoulou, I.; Batzios, C.; Pappaioannou, N.; Ducatelle, R.; Fortomaris, P. Temporary feed restriction partially protects broilers from necrotic enteritis. Avian Pathol. 2014, 43, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Wilkie, D.C.; Van Kessel, A.G.; White, L.J.; Laarveld, B.; Drew, M.D. Dietary amino acids affect intestinal Clostridium perfringens populations in broiler chickens. Can. J. Anim. Sci. 2005, 85, 185–193. [Google Scholar] [CrossRef] [Green Version]
- Singh, Y.; Amerah, A.M.; Ravindran, V. Whole grain feeding: Methodologies and effects on performance, digestive tract development and nutrient utilisation of poultry. Anim. Feed Sci. Technol. 2014, 190, 1–18. [Google Scholar] [CrossRef]
- Tsiouris, V. Poultry management: A useful tool for the control of necrotic enteritis in poultry. Avian Pathol. 2016, 45, 323–325. [Google Scholar] [CrossRef]
- Kiarie, E.; Romero, L.F.; Nyachoti, C.M. The role of added feed enzymes in promoting gut health in swine and poultry. Nutr. Res. Rev. 2013, 26, 71–88. [Google Scholar] [CrossRef] [Green Version]
- Barekatain, M.R.; Antipatis, C.; Rodgers, N.; Walkden-Brown, S.W.; Iji, P.A.; Choct, M. Evaluation of high dietary inclusion of distillers dried grains with solubles and supplementation of protease and xylanase in the diets of broiler chickens under necrotic enteritis challenge. Poult. Sci. 2013, 92, 1579–1594. [Google Scholar] [CrossRef]
- Wu, Y.B.; Ravindran, V. Influence of whole wheat inclusion and xylanase supplementation on the performance, digestive tract measurements and carcass characteristics of broiler chickens. Anim. Feed Sci. Technol. 2004, 116, 129–139. [Google Scholar] [CrossRef]
- Latorre, J.D.; Hernandez-Velasco, X.; Kuttappan, V.A.; Wolfenden, R.E.; Vicente, J.L.; Wolfenden, A.D.; Bielke, L.R.; Prado-Rebolledo, O.F.; Morales, E.; Hargis, B.M.; et al. Selection of Bacillus spp. for cellulase and xylanase production as direct-fed microbials to reduce digesta viscosity and Clostridium perfringens proliferation using an in vitro digestive model in different poultry diets. Front. Vet. Sci. 2015, 2, 1–8. [Google Scholar] [CrossRef]
- Choct, M.; Sinlae, M.; Al-Jassim, R.A.M.; Pettersson, D. Effects of xylanase supplementation on between-bird variation in energy metabolism and the number of Clostridium perfringens in broilers fed a wheat-based diet. Aust. J. Agric. Res. 2006, 57, 1017–1021. [Google Scholar] [CrossRef]
- Wang, G.; Wang, Y.; Yu, L.; Jiang, Y.; Liu, J.; Cheng, Z. New pathogenetic characters of reticuloendotheliosis virus isolated from Chinese partridge in specific-pathogen-free chickens. Microb. Pathog. 2012, 53, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Borda-Molina, D.; Seifert, J.; Camarinha-Silva, A. Current Perspectives of the Chicken Gastrointestinal Tract and Its Microbiome. Comput. Struct. Biotechnol. J. 2018, 16, 131–139. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Toxins | ||||||
|---|---|---|---|---|---|---|
| Type | Alpha (α) | Beta (β) | Epsilon (ε) | Iota (ι) | CPE | NetB |
| (plc o cpa) * | (cpb) * | (etx) * | (iap y ibp) * | (cpe) * | (netB) * | |
| A | + | − | − | − | − | − |
| B | + | + | + | − | − | − |
| C | + | + | − | − | +/− | − |
| D | + | − | + | − | +/− | - |
| E | + | − | − | + | +/− | − |
| F | + | − | − | − | + | − |
| G | + | − | − | − | − | + |
| Type of Toxin | Main Toxin | Diseases that Cause |
|---|---|---|
| A | α | Wound infection in humans (gas gangrene or clostridial myonecrosis), necrotic enteritis in birds, ulcerative abomasitis, mild necrotizing enteritis in piglets, and endotoxemia in South American camelids. |
| α, CPE | Food poisoning in humans, non-food gastrointestinal diseases in humans, and diarrhea in animals such as dogs, pigs, and foals. | |
| α, β2 | Gastrointestinal disease in swine. | |
| B | α, β, ε | Dysentery and hemorrhagic enteritis in lambs and kids. |
| C | α, β | Necrotizing enteritis in humans, enteritis in dogs, chickens, and South American camelids. |
| α, β, β2 | Gastrointestinal disease in swine. | |
| D | α, ε | Enterotoxemia in sheep and goats (pulpy kidney disease). |
| E | α, ι | Enterotoxemia in rabbits, dogs, cattle, and sheep. |
| F | α, CPE | Human food poisoning and non-food associated diarrhea. |
| G | α, NetB | Subclinical necrotic enteritis in chickens. |
| Lesion | Characteristics |
|---|---|
| Necrosis | Mucous discolored, thick, coarse granular texture, moderately firm and adherent or smooth, and moist. Areas of intensely eosinophilic villi covered with clostridia delimited by heterophilic infiltrate with fibrin. |
| Ulcers | Sunken fossae with rough and reddened exposed surface, crater-like, thinning and detachment of mucosa, shiny appearance. Presence of re-epithelialized ulcers on the serous surface. |
| Hemorrhage | Limited in the margin of the lesions or in the intestinal lumen. |
| Thin and flaccid intestinal wall | Detachment of large areas of mucosa that accumulate in the intestinal lumen, loss of smooth muscle tone, presence of discoloration produced by thick dark green bile at the duodenum and proximal jejunum. Smelly gas build-up. |
| Number | Lesions |
|---|---|
| 0 | No apparent injuries. |
| 1 | Thin or brittle wall. Congested intestinal mucosa. |
| 2 | Focal necrosis or ulceration (1 to 5 lesions). |
| 3 | Coalescent multifocal areas of necrosis (6 to 15 lesion targets). |
| 4 | Extent of severe necrosis (more than 16 lesion targets. |
| 5 | Necrosis patches 2–3 cm long (variable amount). |
| 6 | Extensive diffuse necrosis (variable amount). |
| Genus | Strain | Results | Reference |
|---|---|---|---|
| Bacillus | B. amyloliquefaciens H57 | Improvement of feed conversion. Lower score in intestinal lesions caused by C. perfringens. Structural protection of villi at the mucosal level (improves intestinal integrity). | [96] |
| B. coagulans | Improvement of intestinal morphology and cecum and liver damage decreases (P ˂ 0.01). Expression increase of fowlicidin-2, an antimicrobial peptide described in chickens. Increased levels of sIgA and alkaline-phosphatase activity in jejunum. Increase in the expression levels of lysozyme in the jejunum. Inhibition of growth, colonization, and invasion by C. perfringens. | [97] | |
| B. licheniformis H2 | Normalization of disorders in the microbiota caused by infection with C. perfringens. | [98] | |
| Significant suppression of the negative effects on weight gain, decrease in feed consumption, and feed conversion rate (P ˂ 0.05). Increase in the villis height: depth ratio of the crypts in the ileum (P ˂ 0.05). Increase in the activity of antioxidant enzymes and intestinal capacity in ileum, serum, and liver (P ˂ 0.05). Increase in the concentration of Bcl-2 protein in the liver. | [99] | ||
| B. subtilis DSM32315 | Meta-analysis carried out in three different countries and five independent trials. Significantly improves weight gain and feed conversion. Decreases mortality. Lower injury score. | [91] | |
| B. subtilis PB6 | Lower score of intestinal lesions. Increase in the concentration of propionic acid in cecum. | [100] | |
| A mix of 6-probiotic strains, 4 Bacillus subtilis (CPB 011, CPB 029, HP 1.6, and D 014) and 2 Bacillus velezensis (CBP 020 and CPB 035) | Significantly improves feed conversion. Increase in villus height (P ˂ 0.0001) and in the ratio of villi height: crypt depth (P ˂ 0.0004) in duodenum and jejunum. | [101] | |
| Lactobacillus | L. johnsonii BS15 | Significant increase in the levels of IgG (or IgY) and IgA in serum after 21 days. Beneficial effects on subpopulations of T lymphocytes in peripheral blood. | [102] |
| L. plantarum 1.2567 | Significant decrease in the loss of epithelial cells and lymphocyte infiltration, showing an effect of attenuation of the inflammatory response. Significant reduction in intestinal injury scores. Improvement in weight gain. Improvement in the structure of microvilli. | [103] | |
| L. fermentum 1.2029 | Attenuation of the inflammatory damage causing distortion in the crypt architecture, infiltration of granulocytes in the lamina propria and subepithelial and hyperplasia in the lamina propria. Modulation in the expression levels of interferon γ, interleukin IL-10 and the Toll-like receptor 2 receptor. Decrease in the percentage of injury incidence, intestinal injury score and injury severity. | [104] | |
| L. acidophilus CGMCC 1.1878 and L. fermentum CGMCC 1.2029 | In in vitro assays, both strains degraded C. perfringens α-toxin at 2 and 4 h of incubation. The pretreatment of C. perfringens with L. acidophilus significantly decreased (P ˂ 0.05) the percentage of adhesion of the pathogen to chicken intestinal epithelial cells. The relative expression levels of interleukins 6, 8 and 1β, inducible nitric oxide synthase and tumor necrosis factor α (TNF-α) were under-expressed in cells treated with Lactobacillus strains. | [105] | |
| L. plantarum R1.0320 | Increase in the villus height: crypt depth ratio. Greater expression of MUC2 and a decrease in the expression of TNF-α in the mucosa of the ileum. Significant increase in the levels of IgA and IgG (or IgY) (on the 3rd day of administration) and IgM (on the 10th day of administration). | [106] | |
| Enterococcus | E. faecium NCIMB 11181 | Significant improvement in weight gain. Lower rate of intestinal lesions, histopathological inflammation, and apoptosis in intestinal cells. Overexpression of the gene encoding Claudin-1 that promotes epithelial cell attachment. Promote a balance in the intestinal immune response by modulating the expression of pro and anti-inflammatory cytokines, growth factors, heat shock proteins, and negative regulators of signaling mediated by Toll-like receptors. Modulation of the intestinal microbiota. | [107] |
| Probiotic | Concentration on the Diet | Time | Results | Reference |
|---|---|---|---|---|
| Bacillus subtilisCGMCC 1.921 | 1 × 107 and 1 × 108 CFU/g | 1 to 24 weeks | Significantly lower enumeration of C. perfringens in ceca digesta (p ˂ 0.05). | [108] |
| Bacillus subtilisPB6 | 5 × 1011 CFU/kg | 35 days | Reduced the intestinal C. perfringens enumeration significantly (p ˂ 0.05) and improved villi length by 10.88 and 30.46% (p ˂ 0.05) compared with uninfected and infected control groups. | [109] |
| L. acidophilusD2/CSL CECT 4529 and B. subtilis PB6 ATCC-PTA 6737 | 0.1% of L. acidophilus (Lactomalt D2 Bio®); 0.05% of B. subtilis (Clostat® brand dry—740210) | 5 and 7 months | Significantly decrease in E. coli, Clostridia, and Staphylococci in cecum and ileum digesta (p ˂ 0.001). The two probiotic-supplemented diets, increased Lactobacillus spp. and Bifidobacterium spp. enumeration compared with the control diet. | [110] |
| A mix of Bacillus subtilis DSM17299, Clostridium butyricum, and Lactobacillus acidophilus | 2 × 102 CFU/g and 4 × 102 CFU/g | 35 days | Significantly lower enumeration of C. perfringens and Escherichia coli in caecum and increase the enumeration of Lactobacillus and Bifidobacterium. | [111] |
| Enterococcus faeciumNCIMB 11181 | 1 × 106 CFU/kg | 26 days | Microbial community composition among the different groups, indicating significant variability in their microbial profiles. Highest relative abundance of Lactobacillus and Butyricicoccus in the cecum compared to the negative control and the C. perfringens-infected group without the administration of probiotic. | [107] |
| L. johnsonii | Feed and water delivery > 106 CFU/g or mL; oral and litter delivery > 108 CFU/mL of PBS | 21 days | Establishment of the probiotic L. johnsonii in the intestinal tract. No statistically significant differences between delivery methods on the gut microbiota. Significantly decrease enterobacteria and C. perfringens in the ileum. | [112] |
| Synbiotic Composition | Dose | Time | Results | Reference |
|---|---|---|---|---|
| Enterococcus faecium+ FOS + phycophytic substances | 1 kg/ton of feed | 3 weeks | Decrease in mortality rate. Significant improvement (p ˂ 0.05) in the intestinal lesion score. Absence and reduction of histopathological alterations. Significant decrease (p ˂ 0.05) in the counts of C. perfringens in intestine and cecum, from day 3 to day 21, all this between the control group and the infected group fed with the synbiotic. | [138] |
| Saccharomyces cerevisiae, Enterococcus faecium, andBacillusspp. (Avi-Lution®) | 1 and 2 g/Kg of feed | 42 days | Significant increase in weight gain. Decrease in the percentage of mortality and in cumulative mortality at day 28 and 42 (both levels of synbiotic supplementation). No effect on intestinal lesions was observed. | [139] |
| Synbiotic mix Kurago Biotek, 1 mL contains (7 log UFC/g of Lactobacillus rhamnosus HN001, Pediococcus acidilactici MA18/5M and 4.5% Agave tequilana fructans) | 50 µL/day | 39 and 42 days | Increase in lactic acid bacteria enumerations in the duodenum. Improvement in intestinal morphology (higher villi and shallow crypts) in the duodenal mucosa. | [75] |
| L. reuteri, E. faecium, B. animalis,and P. acidilactici con FOS. | 0.05% | 21 and 42 days | Significant difference in the height of the jejunal villi (p ˂ 0.05) on day 28 and 42. Significant weight gain (p ˂ 0.01) (at 21 and 42 days of the experiment). Significant decrease in C. perfringens enumerations from day 28 to day 42. Increasing the number of specific antibodies (IgA) at the level of the ceca mucosa. | [140] |
| Product | Species | Results | Reference |
|---|---|---|---|
| Anise essential oil | Pimpinella anisum | Promotes intestinal development (longer villi and shallow crypts). Decreased intensity of intestinal lesions associated with necrotic enteritis. | [144] |
| Benzophenanthridine (alkaloids) | Chelidonium majus | Improves productive efficiency parameters. Reduces intestinal lesions and mortality associated with necrotic enteritis. | [145] |
| Oregano essential oil | Origanum vulgare | Increase in the body weight and breast weight at 42 d and promotes the cell proliferation in duodenum (P = 0.001) and jejunum (P = 0.012). Significantly decrease in the Clostridium counts. Decrease of gut lesions caused by C. perfringens and improved villus height to crypt depth, improvement of feed conversion efficiency. Increase of serum antibody titers and tendency to elevate occludin mRNA expression at the same time that linearly inhibited the mRNA expression of TLR-2 and tumor necrotic factor-α in the ileum. | [146,147,148] |
| Carvacrol | Origanum vulgare | Improved health (longer villi and shallow crypts) and function of the intestinal barrier. They promote intestinal colonization by Bifidobacterium. Antimicrobial activity against C. perfringens and reduction of intestinal lesions associated with necrotic enteritis. | [141,142,149,150] |
| Curcumin | Curcuma Longa | Decreases C. perfringens enumerations in intestinal contents. | [143] |
| Piperine | Piper nigrum | Decreases C. perfringens enumerations in intestinal contents. | [143] |
| Protopine (alkaloids) | Eschscholzia californica Fumaria officinalis | Improves productive efficiency parameters. Reduces intestinal lesions and mortality associated with necrotic enteritis. | [145] |
| Tannins | Castanea sativa | Inhibits the growth of C. perfringens in vitro and in vivo, without affecting food consumption and weight gain. | [151] |
| Thymol | Thymus vulgaris | Improved health (longer villi and shallow crypts) and function of the intestinal barrier. Promotes intestinal colonization by Bifidobacterium. Antimicrobial activity against C. perfringens. | [142,149,150] |
| Sanguinarin | Chelidonium majus | Improves productive efficiency parameters. Reduces intestinal lesions and mortality associated with necrotic enteritis. | [145] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mora, Z.V.-d.l.; Macías-Rodríguez, M.E.; Arratia-Quijada, J.; Gonzalez-Torres, Y.S.; Nuño, K.; Villarruel-López, A. Clostridium perfringens as Foodborne Pathogen in Broiler Production: Pathophysiology and Potential Strategies for Controlling Necrotic Enteritis. Animals 2020, 10, 1718. https://doi.org/10.3390/ani10091718
Mora ZV-dl, Macías-Rodríguez ME, Arratia-Quijada J, Gonzalez-Torres YS, Nuño K, Villarruel-López A. Clostridium perfringens as Foodborne Pathogen in Broiler Production: Pathophysiology and Potential Strategies for Controlling Necrotic Enteritis. Animals. 2020; 10(9):1718. https://doi.org/10.3390/ani10091718
Chicago/Turabian StyleMora, Zuamí Villagrán-de la, María Esther Macías-Rodríguez, Jenny Arratia-Quijada, Yesica Sughey Gonzalez-Torres, Karla Nuño, and Angélica Villarruel-López. 2020. "Clostridium perfringens as Foodborne Pathogen in Broiler Production: Pathophysiology and Potential Strategies for Controlling Necrotic Enteritis" Animals 10, no. 9: 1718. https://doi.org/10.3390/ani10091718