In the present study, functional analysis of DEGs revealed a set of biological processes, canonical pathways and networks, potentially related with tenderness. In the functional enrichment analyses using FatiGO, there were an overrepresentation of processes related with
Proteolysis such as Positive regulation of proteolysis involved in cellular protein catabolic process (GO:1903052), Cytosolic calcium ion transport (GO:0060401), Regulation of calcium ion transport (GO:0051924) or Actin-myosin filament sliding (GO:0033275);
Skeletal muscle tissue development and growth: Skeletal muscle tissue development GO:0007519), Muscle cell development (GO:0055001), Skeletal muscle cell differentiation (GO:0035914), or Regulation of muscle system process (GO:0090257);
Lipid metabolism: Lipid homeostasis (GO:0055088), Lipid storage (GO:0019915) or Positive regulation of lipid storage (GO:0010884) and
Collagen metabolic process: Collagen metabolic process (GO:0032963), Collagen fibril organization (GO:0030199) or Collagen biosynthetic process (GO:0032964) (
Table 2 and
Table S5).
Next, the most relevant DEGs and their potential implications in the aforementioned biological pathways and processes will be detailed.
4.1. Proteolysis Process
It is well-known that the proteolytic system has a key role in meat tenderization [
34], which is related with the degree of post-mortem alteration of proteins and muscle structure [
35]. Several proteases such as calpains, calpastatins, cathepsins, caspases and kinases are involved in the meat tenderization process [
34]. During the conversion of muscle to meat, cathepsins degrade actomyosin binding [
34] and the weakening of the strong actomyosin interaction imply the widening of sarcomeres. Then, calpains are more able to hydrolyze associated proteins, allowing proteolysis and influencing the maturation of muscle [
36]. In our study, a higher expression of the
Cathepsine C gene (
CTSC) in the Tender group was observed (
Table 1). The functional analyses revealed that this gene is associated with the positive regulation of proteolysis (GO:1903052,
Table 2). In a study comparing muscle expression in the Casertana pig breed with two commercial breeds, an overexpression of
CTSC in Casertana muscle was also observed [
37]. Like the Iberian breed, Casertana is an autochthonous breed characterized for having better meat quality than commercial ones. These results support that higher expression of
CTSC is associated with a higher activation of the proteolysis process favoring the meat tenderization. Besides, in a variant calling analyses based on RNA-seq data of two Polish pig breeds divergent for meat tenderness, variants with different genotype distribution between breeds on
CTSC gene were detected [
38]; however, any association analyses between the genetic variants identified and tenderness have been carried out so far.
Furthermore,
Ras Homolog Family Member D (RHOD) is overexpressed in the Tender group and codifies for a protein involved in reorganization of the actin cytoskeleton. Our functional analysis showed that
RHOD was involved on actin filament organization GO
BP (GO: 0061572) (
Table 2).
RHOD gene maps on the porcine chromosome 2 (5.39 Mb) within a quantitative trait loci (QTL) for shear force detected in the F2 of a Duroc x Pietrain crossbred [
39]. In vitro studies have revealed that the interference of RHOD protein produces a higher cell attachment and diminishes cell migration [
40]. Therefore, higher expression of the
RHOD gene could ease the degradation of actin cytoskeleton during proteolysis.
Two of the most overexpressed genes in the Tough group were
Myosin Heavy Chain 8 (MYH8) and
Actin Alpha 1, Skeletal Muscle (ACTC1), both enclosed in functional network #6 (
Table 3), and functional analysis revealed that
MYH8 and
ACTC1 play a relevant role in GO
BP as actin-myosin filament sliding, structure organization and contraction (
Table 2) and
ACTC1 is also involved in muscle cell development (GO: 0055001). MYH8 protein is related with functions as skeletal muscle contraction, ATPase activity ([
41] and actin filament binding [
42]. A higher expression of this gene was related with muscle hypertrophy in a transcriptome analysis on Canadian double-muscled Large White pigs, which are characterized by having a notable muscle mass [
43].
ACTC1 encodes for a protein involved in skeletal muscle development [
44] and contributes to the structural integrity of cytoskeleton [
45]. Expression differences of
ACTC1 associated to tenderness have been uneven. In a study comparing the transcriptome of
Longissimus dorsi between Shaziling pig, an autochthonous Chinese pig breed with a high-quality meat than Yorkshire, an overexpression of
ACTC1 was observed in Shaziling pig [
46]. However, the study was carried out in 25-day-old pigs and the results could be different in older animals. On the other hand, in a study comparing the
Longissimus dorsi transcriptome of male and female Qinchuan cattle individuals, in which females have tenderer meats, a down-regulation of
ACTC1 gene was observed. In our study, the overexpression of
ACTC1 is apparently associated with tougher meat.
GADD45B gene was overexpressed in the Tough group. This gene encodes for Growth Arrest and DNA Damage Inducible Beta protein, which plays a crucial role in cellular growth arrest and apoptosis, associated with stress signals [
47]. The authors of [
48] observed a higher expression of
GADD45B in cattle
Longissimus thoracis muscle with high ultimate pH values. Alteration of pH implies changes in the regulation of calcium transport pathways into the cellular sarcoplasm. When pH muscle is at isoelectric point (5.2 to 5.5) an increase in calcium concentration in the cell is produced, causing a rise of calpain activity [
49,
50], which degrades myofibrillar and cytoskeletal proteins, promoting meat tenderization [
51]. A disparity of results regarding the relationship between pH and tenderness has been reported by other authors. While [
6] did not observe a phenotypic relation between these traits, [
52] determined that the relationship between pH and tenderness depends on the breed. In this study, we do not have pH values and we cannot conclude that the differential expression of
GADD45B gene among groups could be explained by the pH.
ACTC1 and
RHOD codify for proteins involved on ILK and Integrin signaling pathways (
Table 4). ILK Signaling is related with cell survival and apoptosis [
53] and Integrin Signaling is linked with cell apoptosis and regulation of actin cytoskeleton [
54]. Interestingly, [
15] reported pathways involved in cellular apoptosis (survival) and stress response as important factors of tenderization. Moreover, apoptosis is considered one of the first steps in development of meat tenderization, inducing biochemical and structural muscle changes [
55]. In the same direction that
GADD45B, ACTC1 and
RHOD have been associated with cellular apoptosis.
Other important DEGs that could be involved in proteolysis, with higher expression in the Tender group, were
Calsequestrin 2 (
CASQ2),
Cholinergic Receptor Nicotinic Alpha 9 Subunit (
CHRNA9) and
Ankyrin Repeat Domain 1 protein (
ANKRD1).
CASQ2 codifies for a protein involved in calcium store in the sarcoplasmic reticulum and also modulate calcium homeostasis, calcium release and muscle contraction [
56]. In the functional analyses, there was an enrichment of the
CASQ2 gene in GO annotations related with calcium transport and muscle contraction (
Table 2). Differential expression of
CASQ2 was also observed in several studies contrasting the transcriptome of breeds divergent for several meat quality parameters including tenderness in some cases. These studies compared the muscle transcriptome of Basque vs. Large White [
13], Iberian vs. Duroc × Iberian crossbred [
57] and Wannanhua vs. Yorkshire breeds [
58]. However, in these studies, the highest expression level of
CASQ2 was observed in the breed with the tougher meats. This disagreement could be due to the fact that the expression differences observed in these studies are between breeds divergent for different quality traits and that, in our study, we analyzed the expression differences between Iberian pigs divergent for meat tenderness.
ANKRD1 gene was proposed as candidate gene for meat quality by [
59] since they observed in their study that it could be a transcriptional regulator of myogenesis and of myofibril assembly in porcine LD muscle of Duroc x Pietrain. In the present study, an enrichment of this gene was observed in GO
BP related with cellular assembly involved in morphogenesis and myofibril assembly, actomyosin structure organization and sarcomere organization (
Table S5). In addition,
ANKRD1 is involved in biological processes related with muscular growth as skeletal muscle tissue development and muscle cell differentiation linked to myogenesis (
Table 2). The authors of [
13] also observed a higher expression of
ANKRD1 in Large White than in Basque pigs and proposed that ANKRD1 interacts with CASQ2 protein, which regulates calcium homeostasis in skeletal muscle as it was observed in cardiac muscle [
56]. The overexpression of both genes in tenderer meat group observed in our study would support this hypothesis.
Cholinergic Receptor Nicotinic Alpha 9 Subunit (
CHRNA9) was enriched in a biological process related to the regulation of cytosolic calcium concentration (
Table S5), and the canonical pathway analysis interpreted that
CHRNA9 is involved in Calcium signaling pathway, together with
ACTC1,
CASQ2 and
MYH8 genes (
Table S7). High expression of
CHRNA9 was associated with tenderer meats in F2 animals from Duroc × Pietrain cross [
60]. One more time, a regulation of the calcium releasing to the cytoplasm would have related with proteolytic enzymatic activity and have an influence on meat tenderness.
4.2. Skeletal Muscle Tissue Development and Growth
As we mentioned above, the genetic selection of most common European breeds has usually been focused on improving the efficiency of lean tissue growth. The increase in growth rate and lean meat percentage could alter other meat characteristics such as myofiber composition [
61] which would have an impact on meat tenderness. Nevertheless, it should be noted that Iberian pigs have not been previously selected for this or other related traits.
Our transcriptome analysis revealed some pivotal DEGs related with cellular and muscle development such as
MSTN,
DMD, and
FOS, were overexpressed in the Tough group.
Myostatin (
MSTN) encodes for a protein that inhibits myogenesis. This process consists of the growth and differentiation of muscle. The inhibition or loss of function of this gene produces an increase in muscle and reduced fat mass that have been reported in several animal species as cattle [
62] or sheep [
63]. In pigs,
MSTN null mutations generated in Meishan individuals reproduced the double muscle phenotype and meat from pigs homozygous for the mutation was tenderer than the wild-type ones [
64]. This study agrees with our results, and both seem to be contradictory since we would expect that animals with higher muscle mass have tougher meat. However, the role paper of MSTN on adipogenesis has to be considered too. Despite this general lower fat mass content, an inhibition of adipogenesis in intramuscular preadipocytes isolated from porcine
Longissimus dorsi muscles has been observed [
65]. In the current study, the intramuscular fat content in the animals with tougher meat was lower (%IMF = 3.38 ± 0.73) than that measured in the group with tenderer meat (%IMF = 7.81 ± 2.29). Therefore, this higher expression in the Tough group could inhibit the adipogenesis in intramuscular fat of these animals.
DMD encodes for dystrophin protein, which has a relevant role in structural function stabilizing the sarcolemma and anchoring the extracellular matrix to the cytoskeleton via F-actin [
66]. The authors of [
67] suggested that a decrease in the activity of this essential protein may result in progressive porcine
Biceps femoris muscle degeneration and wasting. Network #6 (
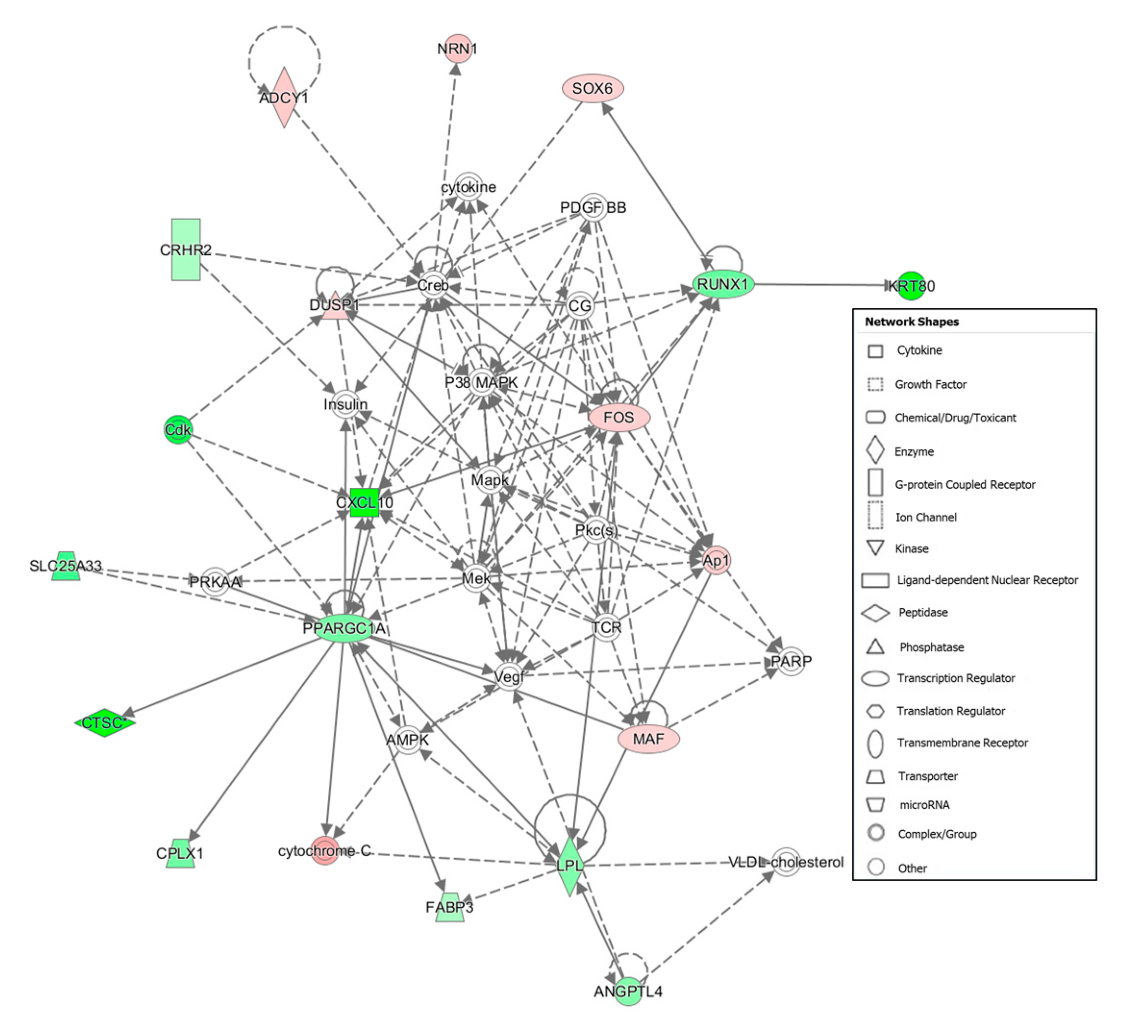
Figure 2) shows that
DMD could activate
MYH8. Therefore, higher expression of
DMD seems to result in a better assembly of actin filament binding, which could be more resistant to degradation by proteases hindering the meat tenderization.
Fos proto-oncogene (
FOS) belongs to the immediate early gene family of transcription factors.
FOS is involved in the maintenance of cytoskeleton, cell-grown regulation, proliferation and differentiation [
68].
FOS gene maps in a QTL for skeletal muscle fiber detected in a Meishan x Pietrain F2 [
69] and codifies for a transcription factor involved that has been previously identified as regulating myogenesis [
70]. Differential expression of this gene on muscle has been observed between different breeds divergent for growth and meat quality at different age stages [
13,
71,
72]. In the current study, functional analyses related this gene with skeletal muscle tissue development and cell differentiation (
Table 2) as well as connective tissue development (
Figure 1). Moreover, IPA analysis showed that
FOS participates on the IGF-1 signaling pathway (
Table S7), which is involved in the activation of receptor tyrosine kinase activity, thereby initiating cell proliferation, cell differentiation and cell survival [
73,
74] also is an important regulator of cellular growth and metabolism [
73].
4.3. Lipid Metabolism
It is well known that the intramuscular fat (IMF) content is a main determinant of tenderness in pig. The positive relation between IMF and tenderness could be due to fat cell expansion that may open the muscle structure favoring the muscle separation [
75]. However, this relationship is controversial, and it is very influenced by the pig breed [
52]. Both IMF and tenderness are heritable traits and the positive genetic correlation among them suggests a common genetic background between IMF and tenderness has also been reported by several authors [
76,
77]. In addition, some transcriptome studies have shown that genes encoding proteins implicated in IMF accretion are overexpressed in tender pork [
78] and it was also proposed that higher IMF content could ease the tenderization associated with the cooking process [
78].
Interestingly, our results showed some DEGs involved in biological process related with lipid metabolism that are overexpressed in animals with tenderer meat such as
Fatty Acid Binding Protein 3 (FABP3) and
Peroxisome Proliferator-Activated Receptor Gamma Coactivator 1-Alpha (PPARGC1A).
FABP3 encodes for a member of the fatty acid-binding protein family that comprises a group of small cytosolic proteins, which specifically bind and transport intracellular fatty acids. There are several studies that find associations between polymorphisms in
FABP3 gene and IMF in different pig breeds [
79,
80,
81]. Besides, [
80] reported associations between polymorphisms mapped in the
FABP3 gene and tenderness, and a positive correlation between the expression of this gene and IMF in muscle of a Korean x Yorkshire F2.
PPARGC1A codifies for a transcription factor which regulates hormone receptors and transcription factors involved in adipogenesis and adipocyte differentiation [
82] also promotes the fiber conversion to oxidative-type ones [
83]. Therefore, this protein could be related with tenderness not only favoring the adipogenesis and IMF content but also for its influence in muscle fiber composition. Actually, there are several studies that report association between polymorphisms located in this gene and tenderness in a commercial hybrid pig population [
84]. In the current study,
Figure 1 shows as
PPARGC1A activates
FABP3 and
CTSC that could suggest favoring the adipogenesis and proteolysis in the group with tenderer meat. Furthermore,
PPARGC1A is involved in PPARα/RXRα Activation and White Adipose Tissue Browning pathways, which presented a trend for activation in the Tender group and are related with lipid metabolism. Peroxisome proliferator-activated receptor-α (PPARα) heterodimerizes with retinoid x receptor (RXR) and play a role in the transcription of regulator genes of adipocyte differentiation and fatty acid oxidation [
85].
It is well known that there is a moderate antagonism between muscular development and intramuscular fat content (IMF) in pigs [
86]. In the previous study carried out in the same pig population [
24], an overexpression of genes related with myogenesis and skeletal muscle development on animals with low IMF content such as
ACTC1,
DMD and
FOS was observed. As we previously mentioned, our findings indicate that
ACTC1,
DMD and
FOS are upregulated in the Tough group, supporting the hypothesis that IMF is related to the tenderization process in Iberian pigs.
4.4. Collagen Metabolic Process
Collagen protein determines the structural support and strength of the extracellular matrix in the connective tissue [
87]. Collagen content depends on animal species and age. For instance, collagen crosslinks in older animals is considered related with tougher meat [
88] and meat tenderness usually decreases when animals are older as well. Therefore, collagen content seems to contribute to meat toughness. High correlation between collagen content and shear force values measured with Warner–Bratzler method on cattle were found [
89]. However, other studies have observed lower correlations in different cattle breeds and ages [
90,
91]. In a theoretical study, [
92] revealed that meat can be ranked in terms of tenderness using the number of collagen crosslink per volume of cooked meat.
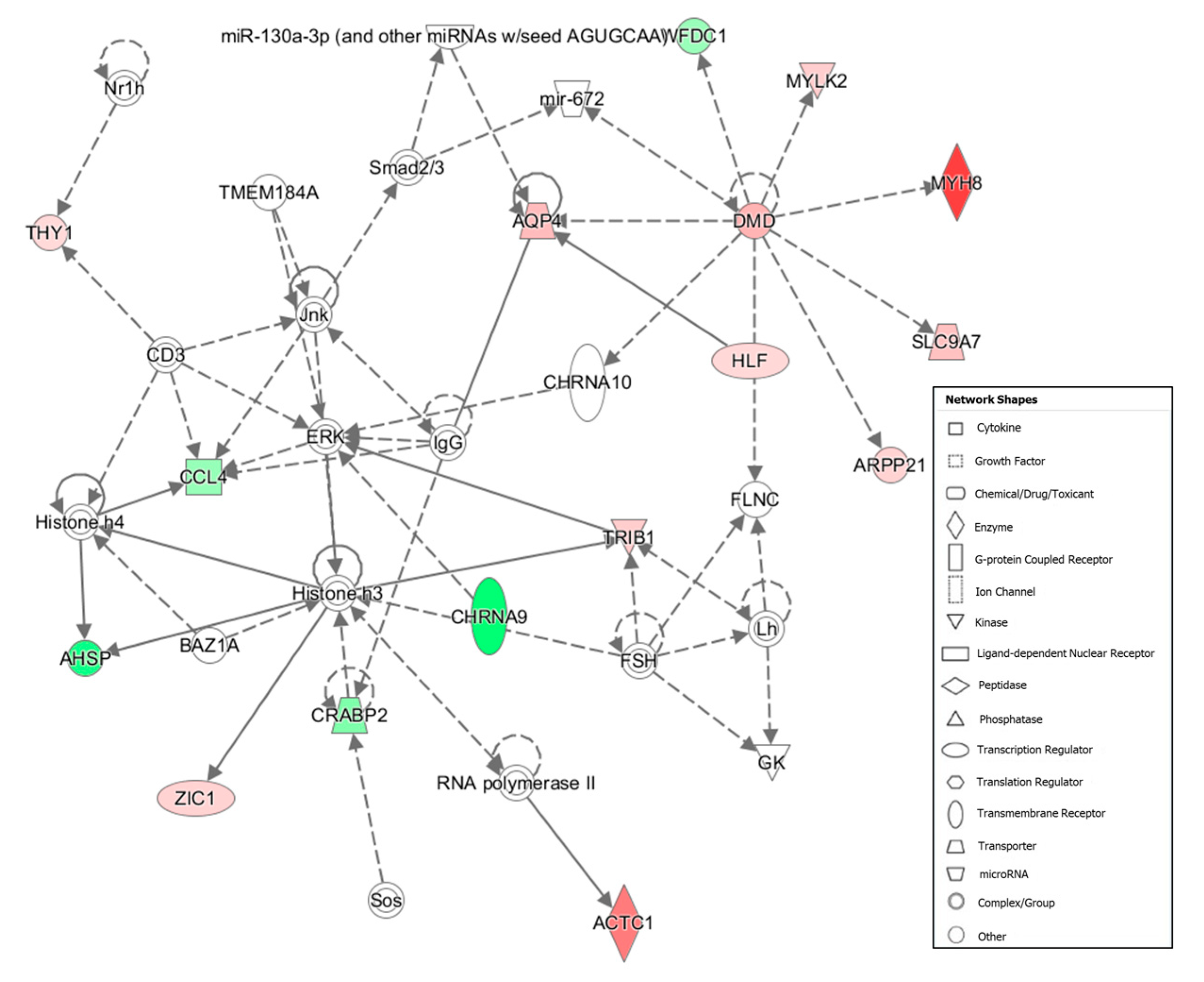
In our study, we detected higher expression of several collagen-encoding genes in tougher meat samples compared with tenderer samples, suggesting a differentiation in collagen constituents between divergent samples for shear force. Our gene ontology analysis revealed that cluster 3 (
Figure S2) contained DEGs from the collagen family (
COL1A1,
COL1A2,
COL12A1 and
COL14A1), upregulated in the Tough group. Among these DEGs cited previously,
Collagen Type XIV Alpha 1 Chain (
COL14A1) encodes for a protein that plays a key role in the extracellular matrix structure organization, cell-cell adhesion and collagen fibril organization [
93]. Other authors have also reported differential expression of
COL14A1 between pigs that, a priori, can be divergent for meat tenderness. In the same sense, here, [
57] showed that
COL14A1 was upregulated in the transcriptome of Duroc x Iberian pigs compared with Iberian purebred pigs, which are expected to have tenderer meat. In addition, higher expression of this gene was observed in Yorkshire pigs than in Wannanhua [
58] and Wei [
94] pig breeds with better meat quality properties.
In summary, the use of two different bioinformatics software for functional analysis showed that some of the most significant differential expressed genes encode proteins that have been involved in similar relevant biological functions, networks and pathways. Genes encoding for proteins involved in proteolysis and activators of the conversion of muscle to meat in post-mortem process are overexpressed in tenderer meat. Otherwise, those genes codifying for proteins that activate myogenesis, stimulate the muscle development and constitute the extracellular matrix of connective tissue are overexpressed in tougher meat.
The results here are very relevant and support that part of the tenderness variability can be explained by genetics. However, tenderness is a complex trait that can be affected by pre-slaughter conditions as stress situations and other post-mortem factors as temperature [
1]. These factors should also be always controlled to avoid undesirable meat textures.
4.5. Transcription Regulatory Factors
A study of the potential regulatory factors explaining the observed expression differences between groups was also carried out. It is not necessary that the regulatory factors are differentially expressed since they can join to DNA sequences adjacent to DEGs with more or less affinity due to potential mutations located in these DNA motifs or in coding sequences of the regulatory factors that could alter the final protein structure.
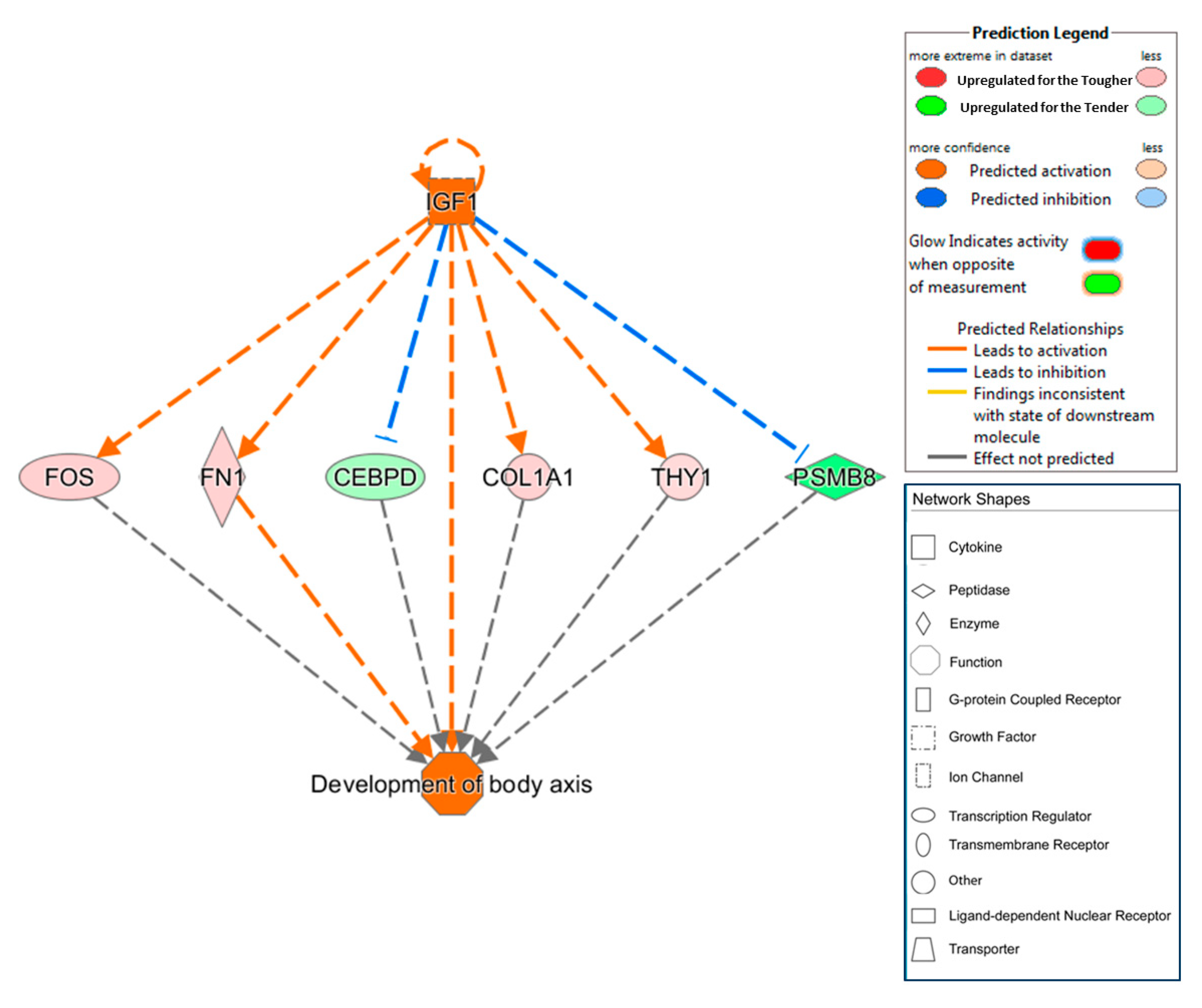
The IPA analyses predicted a regulator effect network that could explain the expression of some DEGs.
Figure 3 represented causal hypotheses to interpret the regulatory potential mechanism of the upstream regulator IGF1 on
FOS, FN1, COL1A1 and
THY1. Apparently, IGF1 activates the expression of
FOS, FN1, COL1A1 and
THY1 that are overexpressed in Iberian pigs with tougher meats and inhibits the expression of CCAAT/enhancer binding protein delta (
CEBPD) and proteasome subunit beta 8 (
PSMB8) which are repressed in this type of pigs. As was explained above,
FOS is involved in muscle growth and development and
COL1A1 in the extracellular matrix constitution; therefore, a higher activation of this process seems to make tougher meat. On the other hand,
CEBPD plays an essential role during the earliest phases of the adipocyte differentiation [
95] and
PSMB8 maps in a genomic region explaining part of the IMF phenotypical variance observed in Iberian pigs [
96]. Therefore, IGF1 would activate the muscle growth and inhibit adipogenesis explaining the antagonism relationship between these traits.
One of the most significant regulatory factors is vestigial-like family member 3 (VGLL3) (
Table 5), which was identified as a transcriptional co-factor associated with myogenesis, skeletal muscle development and muscle hypertrophy [
97]. VGLL3 was predicted to be activated in tougher samples inducing the expression of
COL12A1, COL1A1,
COL1A2 and
GADD45B genes. Some of them are involved in collagen metabolic process (
COL12A1, COL1A1, and
COL1A2), cellular growth and apoptosis (
GADD45B). Therefore, a higher activity of this transcription factor would hinder the tenderization process.
Peroxisome proliferator-activated receptor gamma (PPARG) is a ligand-dependent nuclear receptor known as the “master regulator of adipogenesis”, being related with lipid metabolism processes as adipose differentiation [
98] and it has been identified as a potential candidate genes for improving IMF content [
99]. Moreover, a higher expression of PPARG gene have been observed in Iberian piglets [
100] and foetuses [
72] than in Duroc x Iberian piglets and Large-White foetuses, respectively, which have less IMF content than Iberian ones. In this study, although the expression differences of this gene were not observed, the upstream analysis identified this gene as an activator of
FABP3 and
PPARGC1A, which, as has been pointed out above, are overexpressed in the Tender group and promote adipogenesis and increase IMF content.
4.6. Candidate Genes for an Iberian Pig-Breeding Program
The ultimate objective of this study was to propose several candidate genes for searching polymorphisms and design a genotyping panel for improving tenderness in Iberian pigs. In further steps, polymorphisms with divergent allelic frequencies should be identified in the regulatory regions of the proposed candidate genes and association analyses between their genotypes and shear force should be carried out in the same Iberian pig population.
In summary, the most promising candidate genes to be selected are involved in proteolysis processes (ACTC1, ANKRD1, CHRNA9, CTSC and RHOD), skeletal muscle tissue development and growth (DMD and FOS), lipid metabolism (FABP3 and PPARGC1A) and collagen metabolic process (COL14A1). Although the MSTN gene is clearly involved in muscle growth, the results observed here are controversial since this protein inhibits myogenesis and a higher expression in the Tough group was observed; therefore, more cautions should be taken before to be included as a selection marker.
It is also interesting to consider genes encoding regulator factors such as IGF1, PPARG and VGLL3 since they modulate the expression of some of the genes mentioned before.
It is worth mentioning that some genes, such as ACTC1, DMD and FOS, were also overexpressed in Iberian pigs with low IMF content (Muñoz et al., 2018); therefore, they could be used for improving both IMF content as shear force (tenderness).

 ,
, 



{kind=link}
{kind=link}
{kind=link}