Effects of Bacillus amyloliquefaciens and Bacillus pumilus on Rumen and Intestine Morphology and Microbiota in Weanling Jintang Black Goat



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Experimental Design
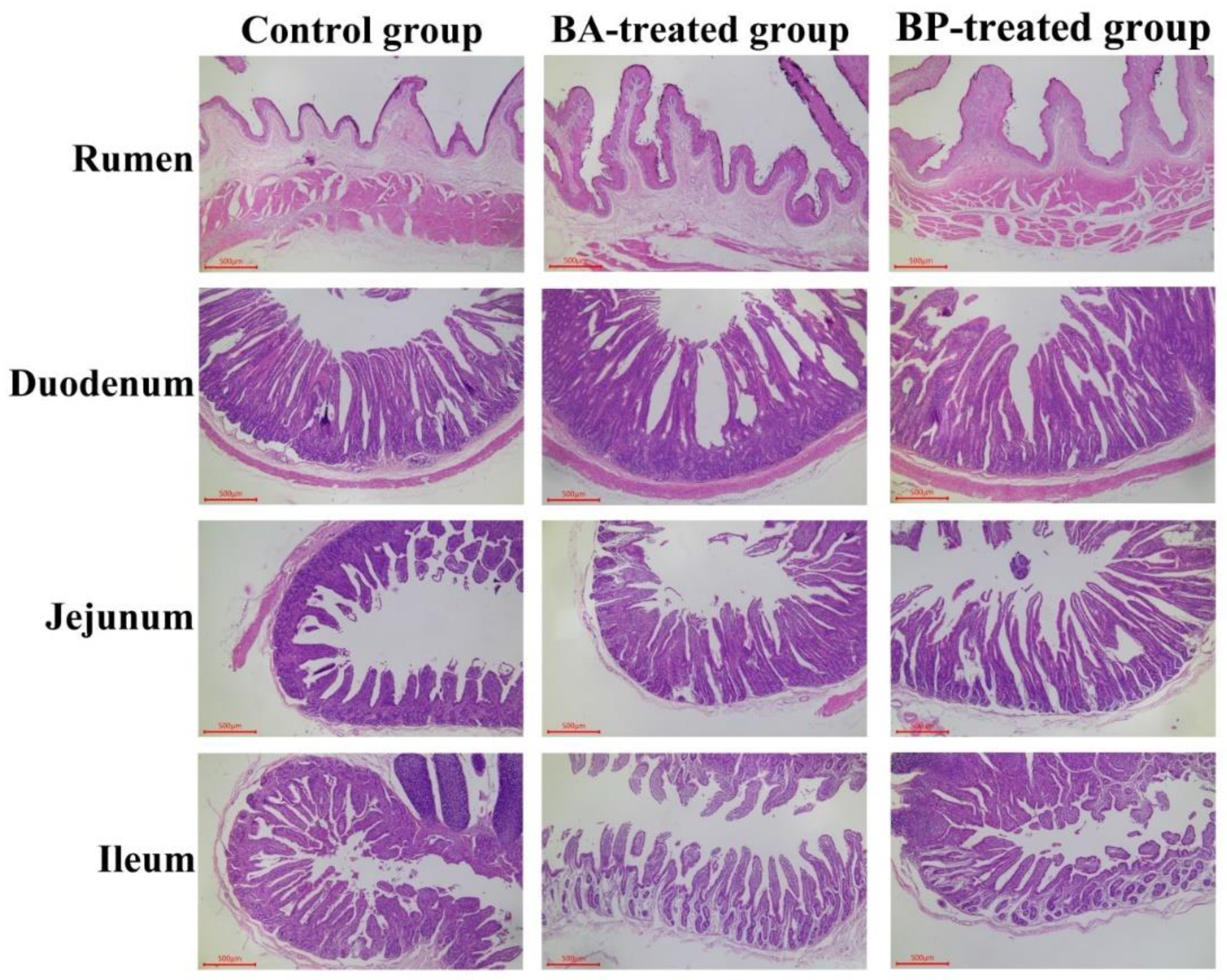
2.2. Effect of Bacillus amyloliquefaciens and Bacillus pumilus on Rumen and Small Intestine Morphology
2.3. Gut Microbiota Analyses
2.4. Statistical Analysis
3. Results
3.1. Effect of Bacillus amyloliquefaciens and Bacillus pumilus on Rumen and Small Intestine Morphology
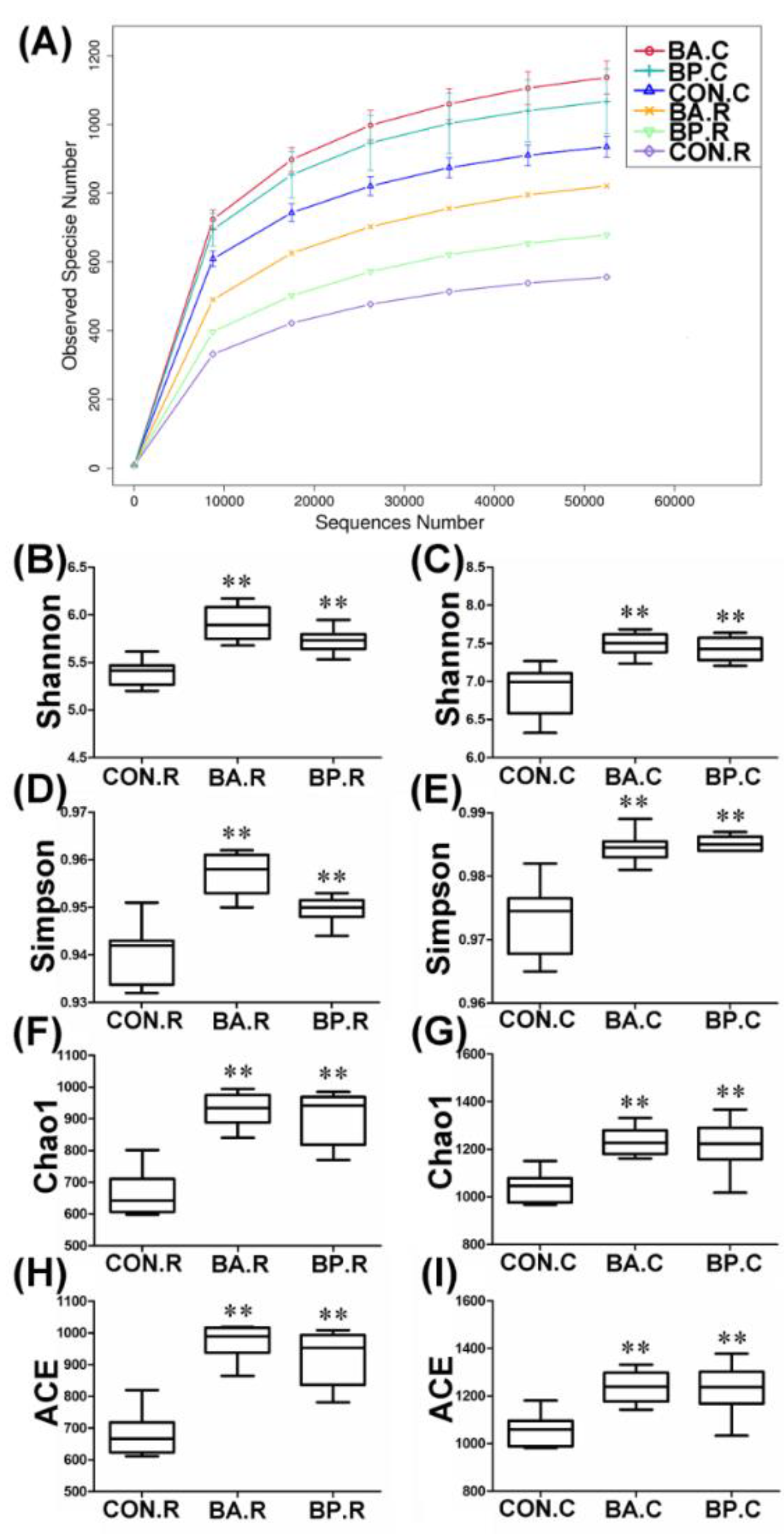
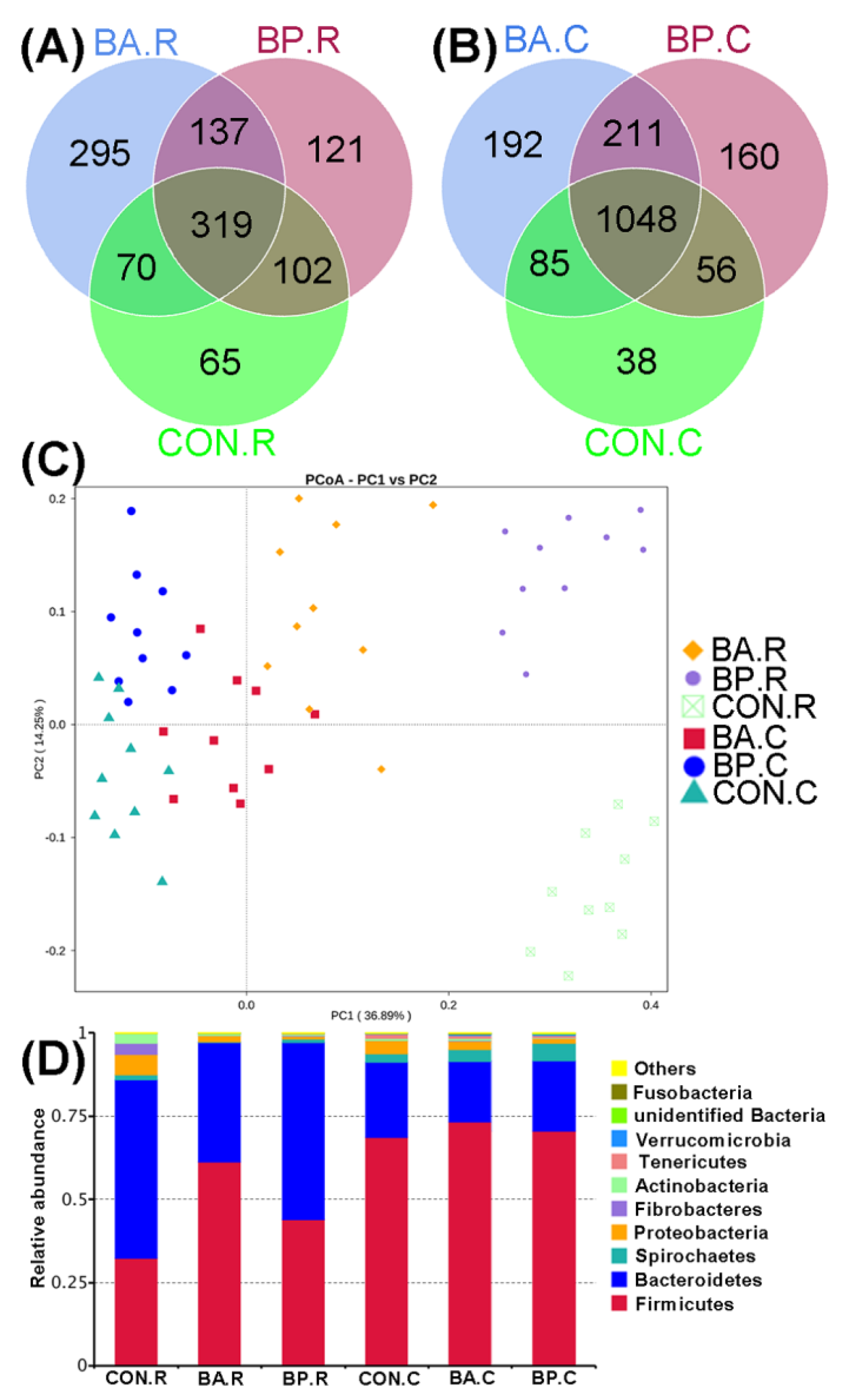
3.2. Analyzing the Bacterial Community Structure by 16S rRNA Sequencing
3.3. Diversity and Compositions of Microbiota
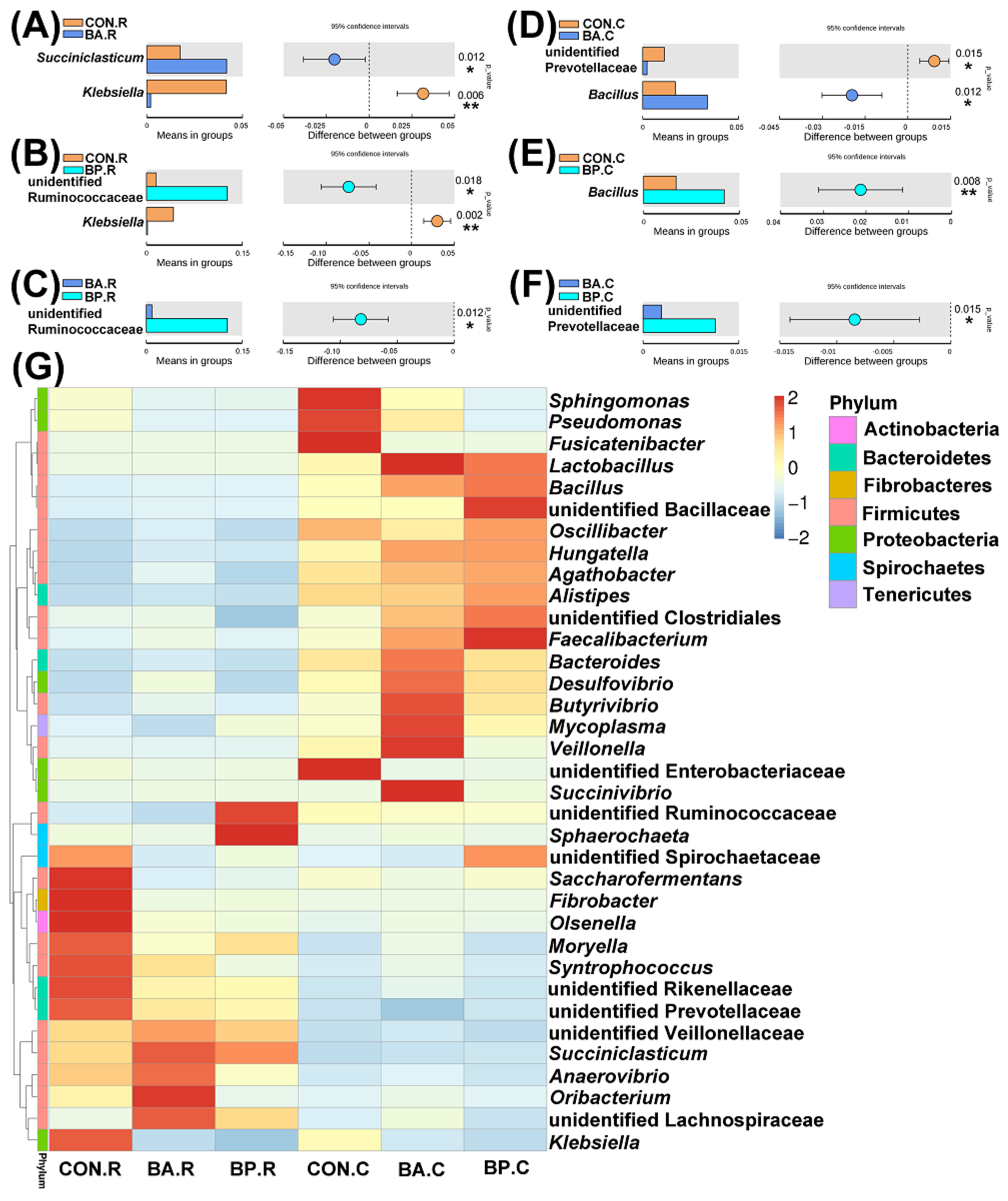
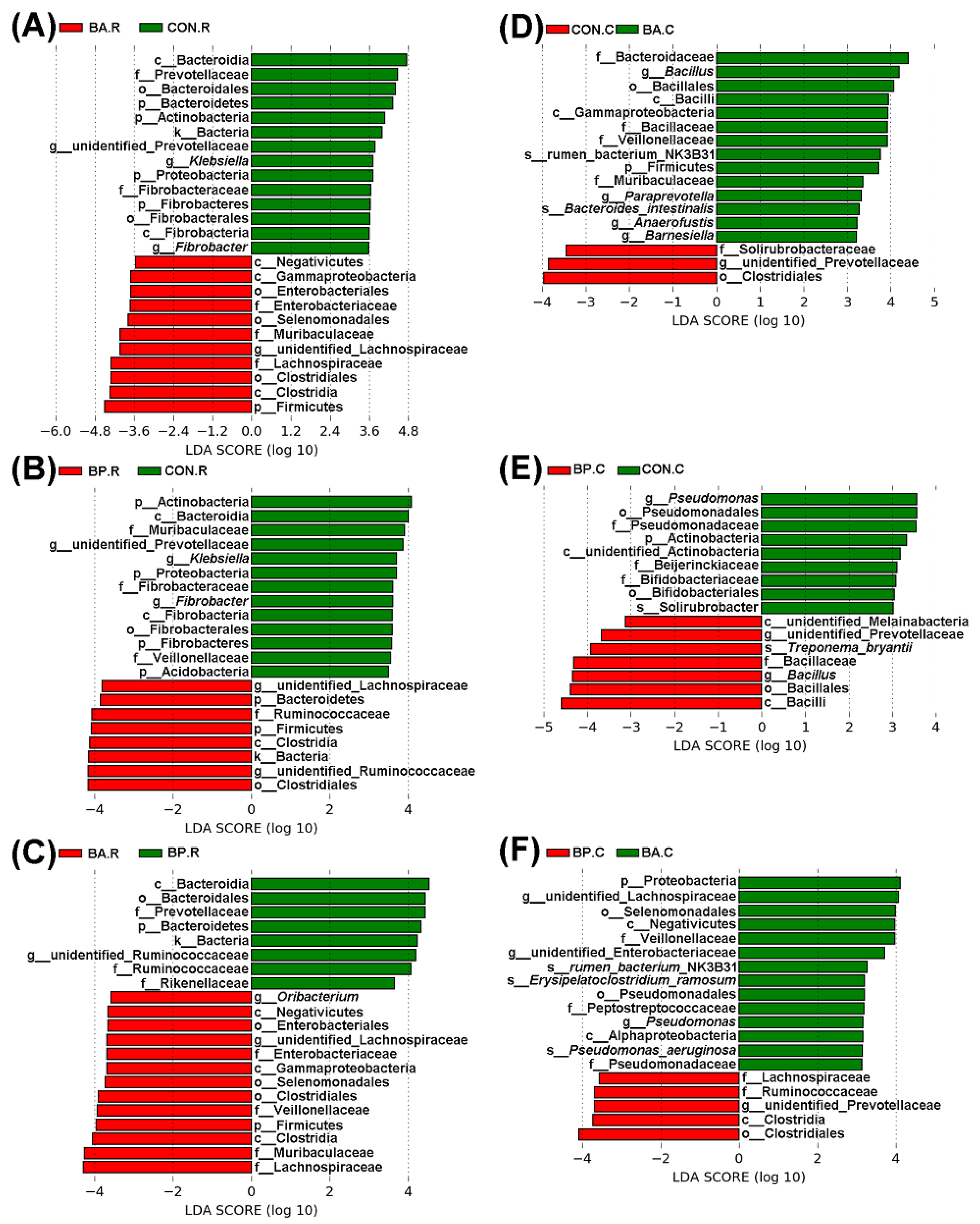
3.4. Differences in Microbial Communities
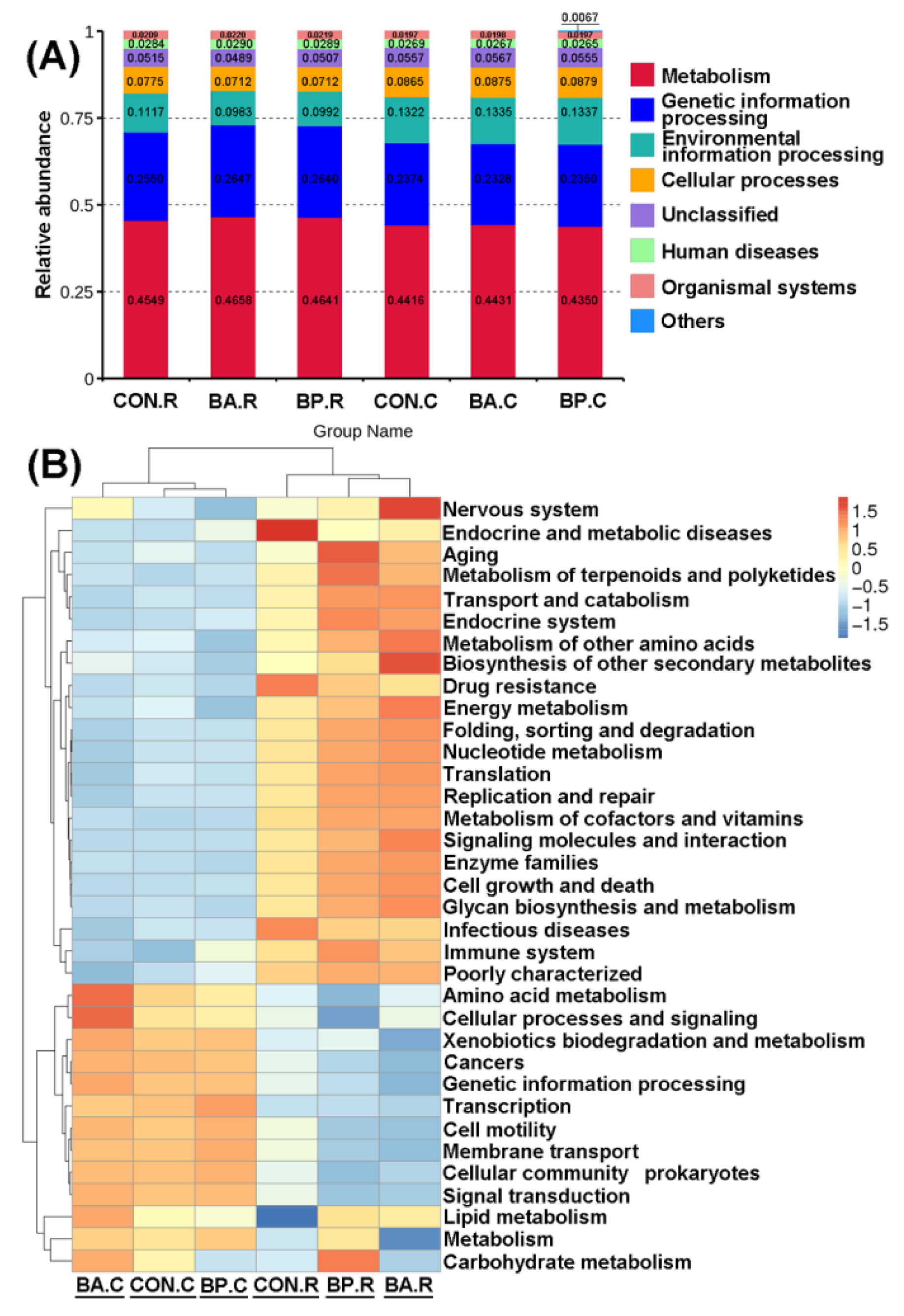
3.5. Estimated Functional Profiles of Microbial Communities
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Ethics statement
Data Availability
References
- Olmos, J.; Acosta, M.; Mendoza, G.; Pitones, V. Bacillus subtilis, an ideal probiotic bacterium to shrimp and fish aquaculture that increase feed digestibility, prevent microbial diseases, and avoid water pollution. Arch. Microbiol. 2020, 202, 427–435. [Google Scholar] [CrossRef]
- Grant, A.; Gay, C.G.; Lillehoj, H.S. Bacillus spp. as direct-fed microbial antibiotic alternatives to enhance growth, immunity, and gut health in poultry. Avian Pathol. 2018, 47, 339–351. [Google Scholar] [CrossRef]
- Harwood, C.R.; Mouillon, J.M.; Pohl, S.; Arnau, J. Secondary metabolite production and the safety of industrially important members of the Bacillus subtilis group. FEMS Microbiol. Rev. 2018, 42, 721–738. [Google Scholar] [CrossRef] [PubMed]
- Blavi, L.; Jørgensen, J.N.; Stein, H.H. Effects of Bacillus amyloliquefaciens and Bacillus subtilis on ileal digestibility of AA and total tract digestibility of CP and gross energy in diets fed to growing pigs. J. Anim. Sci. 2019, 97, 727–734. [Google Scholar] [CrossRef]
- Lan, R.; Kim, I.H. Effects of Bacillus licheniformis and Bacillus subtilis complex on growth performance and faecal noxious gas emissions in growing-finishing pigs. J. Sci. Food Agric. 2019, 99, 1554–1560. [Google Scholar] [CrossRef] [PubMed]
- Luan, S.; Duersteler, M.; Galbraith, E.A.; Cardoso, F.C. Effects of direct-fed Bacillus pumilus 8G-134 on feed intake, milk yield, milk composition, feed conversion, and health condition of pre- and postpartum Holstein cows. J. Dairy Sci. 2015, 98, 6423–6432. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Wu, Y.; Wang, B.; Xu, H.; Mei, X.; Xu, X.; Zhang, X.; Ni, J.; Li, W. Bacillus amyloliquefaciens SC06 protects mice against high-fat diet-induced obesity and liver injury via regulating host metabolism and gut microbiota. Front. Microbiol. 2019, 10, 1161. [Google Scholar] [CrossRef]
- Truong Thy, H.T.; Tri, N.N.; Quy, O.M.; Fotedar, R.; Kannika, K.; Unajak, S.; Areechon, N. Effects of the dietary supplementation of mixed probiotic spores of Bacillus amyloliquefaciens 54A, and Bacillus pumilus 47B on growth, innate immunity and stress responses of striped catfish (Pangasianodon hypophthalmus). Fish Shellfish Immunol. 2017, 60, 391–399. [Google Scholar] [CrossRef]
- Prieto, M.L.; O’Sullivan, L.; Tan, S.P.; McLoughlin, P.; Hughes, H.; Gutierrez, M.; Lane, J.A.; Hickey, R.M.; Lawlor, P.G.; Gardiner, G.E. In vitro assessment of marine Bacillus for use as livestock probiotics. Mar. Drugs 2014, 12, 2422–2445. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.L.; Sun, Y.Z.; Hu, X.; Ye, J.D.; Lu, K.L.; Hu, L.H.; Zhang, J.J. Bacillus pumilus SE5 originated PG and LTA tuned the intestinal TLRs/MyD88 signaling and microbiota in grouper (Epinephelus coioides). Fish Shellfish Immunol. 2019, 88, 266–271. [Google Scholar] [CrossRef]
- Long, S.F.; Xu, Y.T.; Pan, L.; Wang, Q.Q.; Wang, C.L.; Wu, J.Y.; Wu, Y.Y.; Han, Y.M.; Yun, C.H.; Piao, X.S. Mixed organic acids as antibiotic substitutes improve performance, serum immunity, intestinal morphology and microbiota for weaned piglets. Anim. Feed Sci. Technol. 2018, 235, 23–32. [Google Scholar] [CrossRef]
- Cromwell, G.L. Why and how antibiotics are used in swine production. Anim. Biotechnol. 2002, 13, 7–27. [Google Scholar] [CrossRef]
- Iji, P.A.; Saki, A.; Tivey, D.R. Body and intestinal growth of broiler chicks on a commercial starter diet. 3. Development and characteristics of tryptophan transport. Br. Poult. Sci. 2001, 42, 523–529. [Google Scholar] [CrossRef] [PubMed]
- Suo, C.; Yin, Y.; Wang, X.; Lou, X.; Song, D.; Wang, X.; Gu, Q. Effects of Lactobacillus plantarum ZJ316 on pig growth and pork quality. BMC Vet. Res. 2012, 8, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomsen, L.E.; Knudsen, K.E.; Hedemann, M.S.; Roepstorff, A. The effect of dietary carbohydrates and Trichuris suis infection on pig large intestine tissue structure, epithelial cell proliferation and mucin characteristics. Vet. Parasitol. 2006, 142, 112–122. [Google Scholar] [CrossRef] [PubMed]
- Du, R.; Jiao, S.; Dai, Y.; An, J.; Lv, J.; Yan, X.; Wang, J.; Han, B. Probiotic Bacillus amyloliquefaciens C-1 improves growth performance, stimulates GH/IGF-1, and regulates the gut microbiota of growth-retarded beef calves. Front. Microbiol. 2018, 9, 2006. [Google Scholar] [CrossRef] [Green Version]
- Tong, J.; Zhang, H.; Yang, D.; Zhang, Y.; Xiong, B.; Jiang, L. Illumina sequencing analysis of the ruminal microbiota in high-yield and low-yield lactating dairy cows. PLoS ONE 2018, 13, e0198225. [Google Scholar] [CrossRef] [Green Version]
- Do, T.H.; Le, N.G.; Dao, T.K.; Nguyen, T.; Le, T.L.; Luu, H.L.; Nguyen, K.; Nguyen, V.L.; Le, L.A.; Phung, T.N.; et al. Metagenomic insights into lignocellulose-degrading genes through Illumina-based de novo sequencing of the microbiome in Vietnamese native goats’ rumen. J. Gen. Appl. Microbiol. 2018, 64, 108–116. [Google Scholar] [CrossRef] [Green Version]
- Zeineldin, M.; Megahed, A.; Burton, B.; Blair, B.; Aldridge, B.; Lowe, J.F. Effect of single dose of antimicrobial administration at birth on fecal microbiota development and prevalence of antimicrobial resistance genes in piglets. Front. Microbiol. 2019, 10, 1414. [Google Scholar] [CrossRef]
- Saggese, A.; Culurciello, R.; Casillo, A.; Corsaro, M.M.; Ricca, E.; Baccigalupi, L. A marine isolate of Bacillus pumilus secretes a pumilacidin active against Staphylococcus aureus. Mar. Drugs 2018, 16, 180. [Google Scholar] [CrossRef] [Green Version]
- Prieto, M.L.; O’Sullivan, L.; Tan, S.P.; McLoughlin, P.; Hughes, H.; O’Donovan, O.; Rea, M.C.; Kent, R.M.; Cassidy, J.P.; Gardiner, G.E.; et al. Evaluation of the efficacy and safety of a marine-derived Bacillus strain for use as an in-feed probiotic for newly weaned pigs. PLoS ONE 2014, 9, e88599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saputra, F.; Shiu, Y.L.; Chen, Y.C.; Puspitasari, A.W.; Danata, R.H.; Liu, C.H.; Hu, S.Y. Dietary supplementation with xylanase-expressing B. amyloliquefaciens R8 improves growth performance and enhances immunity against Aeromonas hydrophila in Nile tilapia (Oreochromis niloticus). Fish Shellfish Immunol. 2016, 58, 397–405. [Google Scholar] [PubMed]
- Bäumler, A.J.; Sperandio, V. Interactions between the microbiota and pathogenic bacteria in the gut. Nature 2016, 535, 85–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lettat, A.; Nozière, P.; Silberberg, M.; Morgavi, D.P.; Berger, C.; Martin, C. Rumen microbial and fermentation characteristics are affected differently by bacterial probiotic supplementation during induced lactic and subacute acidosis in sheep. BMC Microbiol. 2012, 12, 142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, D.; Chang, S.Y.; Bogere, P.; Won, K.; Choi, J.Y.; Choi, Y.J.; Lee, H.K.; Hur, J.; Park, B.Y.; Kim, Y.; et al. Beneficial roles of probiotics on the modulation of gut microbiota and immune response in pigs. PLoS ONE 2019, 14, e0220843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Z.Z.; Cheng, Y.Y.; Wang, S.Q.; Ge, J.Z.; Shi, H.P.; Kou, J.C. Positive effects of dietary supplementation of three probiotics on milk yield, milk composition and intestinal flora in Sannan dairy goats varied in kind of probiotics. J. Anim. Physiol. Anim. Nutr. 2020, 104, 44–55. [Google Scholar] [CrossRef]
- Zhang, K.; Li, B.; Guo, M.; Liu, G.; Yang, Y.; Wang, X.; Chen, Y.; Zhang, E. Maturation of the goat rumen microbiota involves three stages of microbial colonization. Animals 2019, 9, 1028. [Google Scholar] [CrossRef] [Green Version]
- Hong, Y.; Cheng, Y.; Li, Y.; Li, X.; Zhou, Z.; Shi, D.; Li, Z.; Xiao, Y. Preliminary study on the effect of Bacillus amyloliquefaciens TL on cecal bacterial community structure of broiler chickens. BioMed Res. Int. 2019, 2009, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Ferreira-Halder, C.V.; Faria, A.; Andrade, S.S. Action and function of Faecalibacterium prausnitzii in health and disease. Best Pract. Res. Clin. Gastroenterol. 2017, 31, 643–648. [Google Scholar] [CrossRef]
- Savin, K.W.; Zawadzki, J.; Auldist, M.J.; Wang, J.; Ram, D.; Rochfort, S.; Cocks, B.G. Faecalibacterium diversity in dairy cow milk. PLoS ONE 2019, 14, e0221055. [Google Scholar] [CrossRef] [Green Version]
- Quan, Y.; Song, K.; Zhang, Y.; Zhu, C.; Shen, Z.; Wu, S.; Luo, W.; Tan, B.; Yang, Z.; Wang, X. Roseburia intestinalis-derived flagellin is a negative regulator of intestinal inflammation. Biochem. Biophys. Res. Commun. 2018, 501, 791–799. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Diet Level |
|---|---|
| Ingredients | % |
| Corn | 47.20 |
| Soybean meal | 19.00 |
| Chinese wildrye grass hay | 15.00 |
| Wheat bran | 7.60 |
| Corn germ meal | 7.50 |
| Limestone | 1.40 |
| Calcium hydrogen phosphate | 0.45 |
| NaCl | 0.45 |
| L-Lysine | 0.40 |
| Premix * | 1.00 |
| Analyzed composition (% of dry matter) | |
| Metabolizable energy (MJ/kg) | 10.37 |
| Crude protein | 14.63 |
| Ether extract | 3.53 |
| Crude ash | 6.45 |
| Neutral detergent fiber | 23.69 |
| Acid detergent fiber | 8.23 |
| Lysine | 0.50 |
| Calcium | 1.04 |
| Phosphorus | 0.45 |
| Items | Groups | SEM | p-Value | ||
|---|---|---|---|---|---|
| CON | BA | BP | |||
| Rumen | |||||
| Papilla length (mm) | 2.68 ± 0.29 C | 3.48 ± 0.43 B | 4.65 ± 0.33 A | 0.163 | <0.001 |
| Papilla width (mm) | 1.07 ± 0.09 Bb | 1.25 ± 0.12 Ba | 1.78 ± 0.23 A | 0.626 | <0.001 |
| Epithelium thickness (μm) | 89.32 ± 5.99Ab | 92.03 ± 2.49 Aab | 96.08 ± 7.34 Aa | 1.121 | 0.040 |
| Duodenum | |||||
| Villus height (μm) | 915.38 ± 56.83 B | 1041.17 ± 32.05 A | 1078.12 ± 26.92 A | 14.794 | <0.001 |
| Crypt depth (μm) | 414.94 ± 9.06 A | 326.01 ± 7.44 C | 360.09 ± 6.01 B | 6.933 | <0.001 |
| V/C | 2.21 ± 0.17 C | 3.19 ± 0.12 A | 2.99 ± 0.08 B | 0.082 | <0.001 |
| Villus width (μm) | 92.17 ± 6.85 | 91.11 ± 9.66 | 92.56 ± 14.75 | 1.927 | 0.954 |
| Jejunum | |||||
| Villus height (μm) | 424.23 ± 37.77 C | 562.80 ± 35.28 B | 818.90 ± 43.14 A | 31.122 | <0.001 |
| Crypt depth (μm) | 214.22 ± 8.11 C | 226.14 ± 7.21 B | 301.44 ± 6.61 A | 7.286 | <0.001 |
| V/C | 1.98 ± 0.16 C | 2.49 ± 0.17 B | 2.72 ± 0.13 A | 0.064 | <0.001 |
| Villus width (μm) | 97.61 ± 17.17 Bb | 115.72 ± 20.17 ABb | 143.31 ± 28.03 Aa | 5.250 | <0.001 |
| Ileum | |||||
| Villus height (μm) | 517.30 ± 44.73 | 558.07 ± 68.07 | 540.69 ± 36.97 | 9.612 | 0.227 |
| Crypt depth (μm) | 299.81 ± 10.46 A | 248.31 ± 10.77 B | 218.98 ± 7.09 C | 6.430 | <0.001 |
| V/C | 1.73 ± 0.17 Bc | 2.25 ± 0.26 Ab | 2.47 ± 0.19 Aa | 0.069 | <0.001 |
| Villus width (μm) | 102.64 ± 13.31 Bb | 132.31 ± 23.99 Aa | 141.10 ± 26.07 Aa | 4.915 | 0.001 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, N.; Wang, L.; Wei, Y. Effects of Bacillus amyloliquefaciens and Bacillus pumilus on Rumen and Intestine Morphology and Microbiota in Weanling Jintang Black Goat. Animals 2020, 10, 1604. https://doi.org/10.3390/ani10091604
Zhang N, Wang L, Wei Y. Effects of Bacillus amyloliquefaciens and Bacillus pumilus on Rumen and Intestine Morphology and Microbiota in Weanling Jintang Black Goat. Animals. 2020; 10(9):1604. https://doi.org/10.3390/ani10091604
Chicago/Turabian StyleZhang, Nanchi, Li Wang, and Yong Wei. 2020. "Effects of Bacillus amyloliquefaciens and Bacillus pumilus on Rumen and Intestine Morphology and Microbiota in Weanling Jintang Black Goat" Animals 10, no. 9: 1604. https://doi.org/10.3390/ani10091604