Evaluation of an Acute Osmotic Stress in European Sea Bass via Skin Mucus Biomarkers

 , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Material & Methods
2.1. Animals and Experimental Procedures
2.2. Stress Biomarkers
2.3. Total Protein Quantification
2.4. Osmolality and Ion Quantifications of Plasma and Skin Mucus
2.5. Mucus Exudation Values
2.6. Statistical Analysis
3. Results
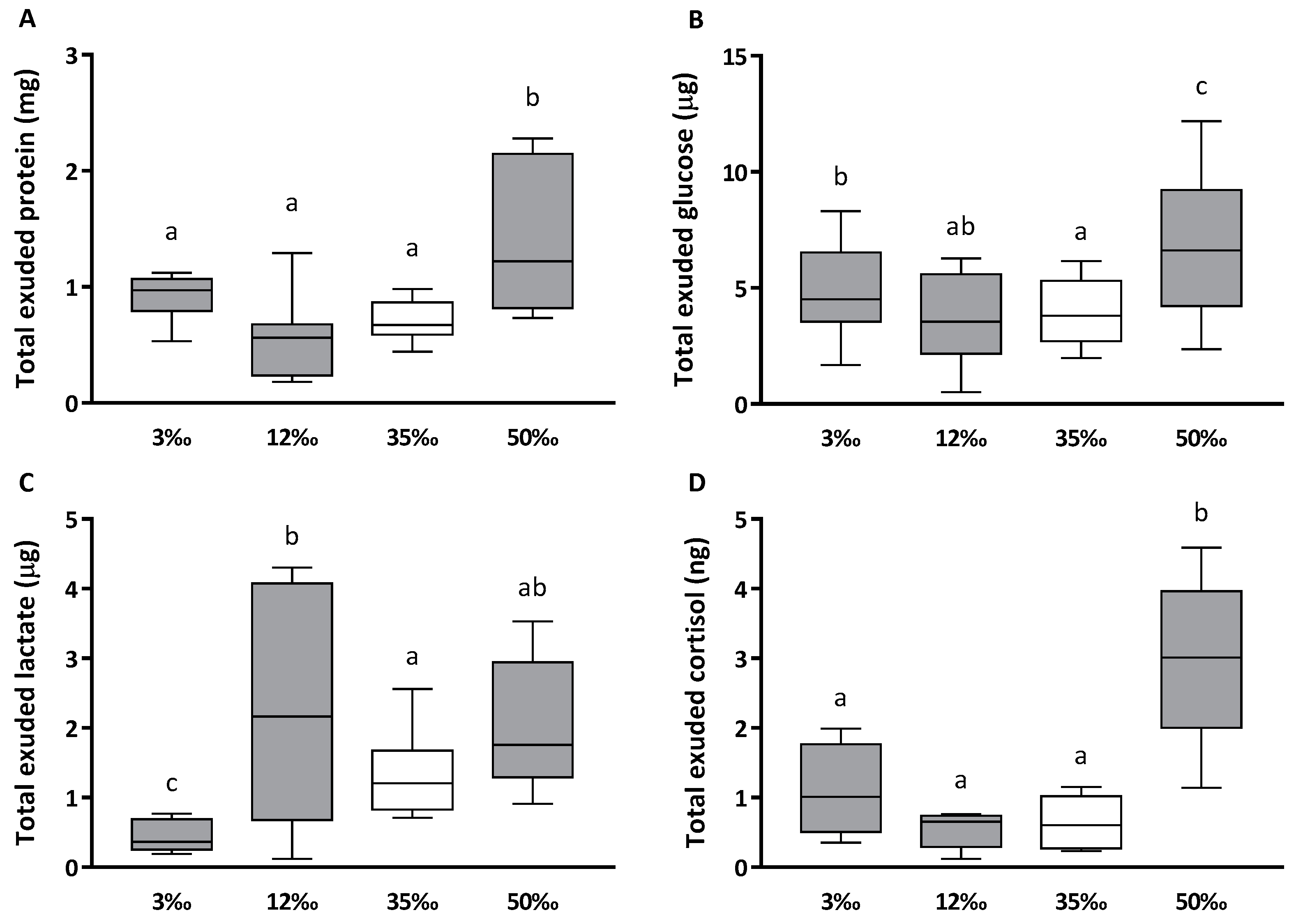
3.1. Mucus and Plasma Biomarkers
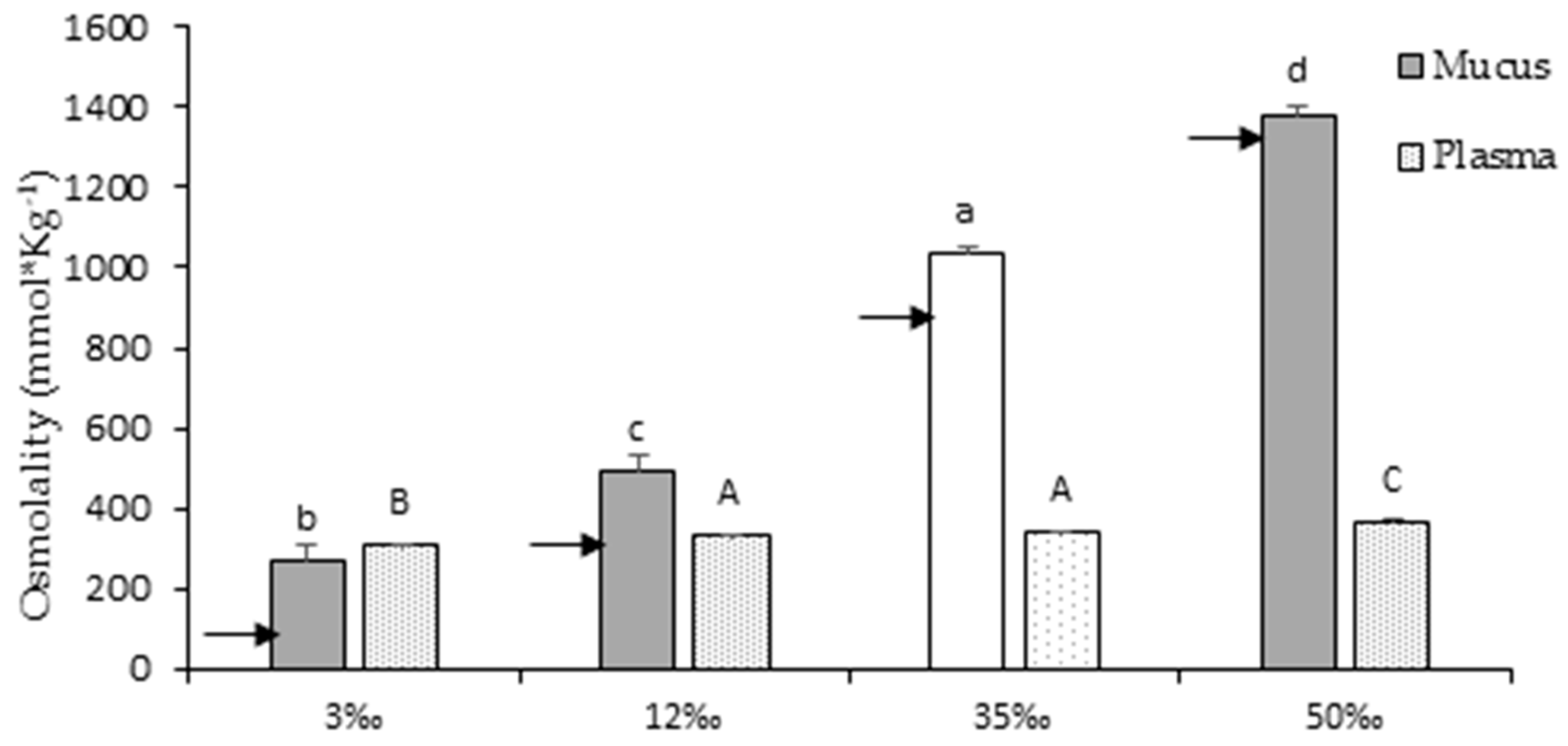
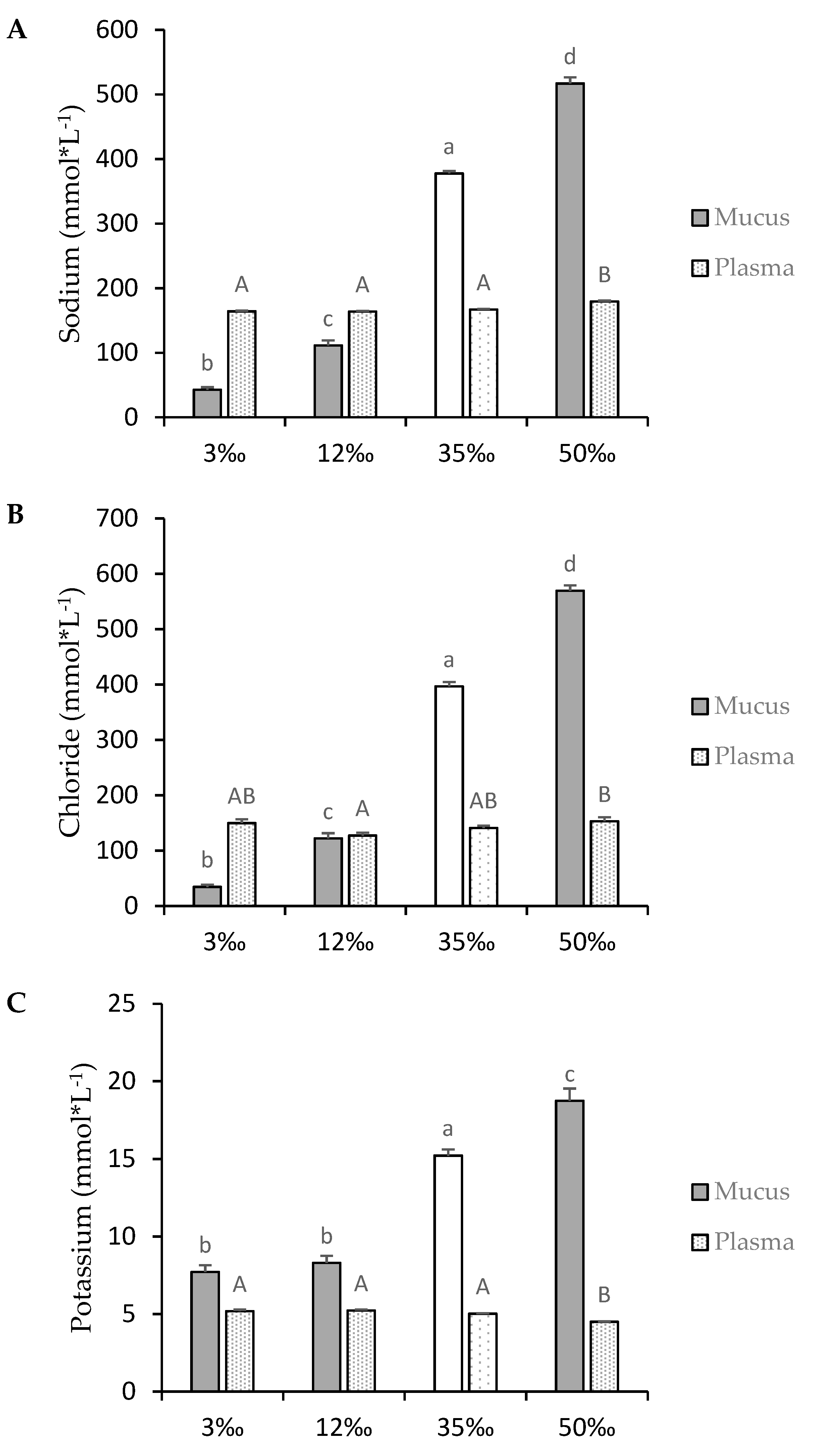
3.2. Osmotic Parameters
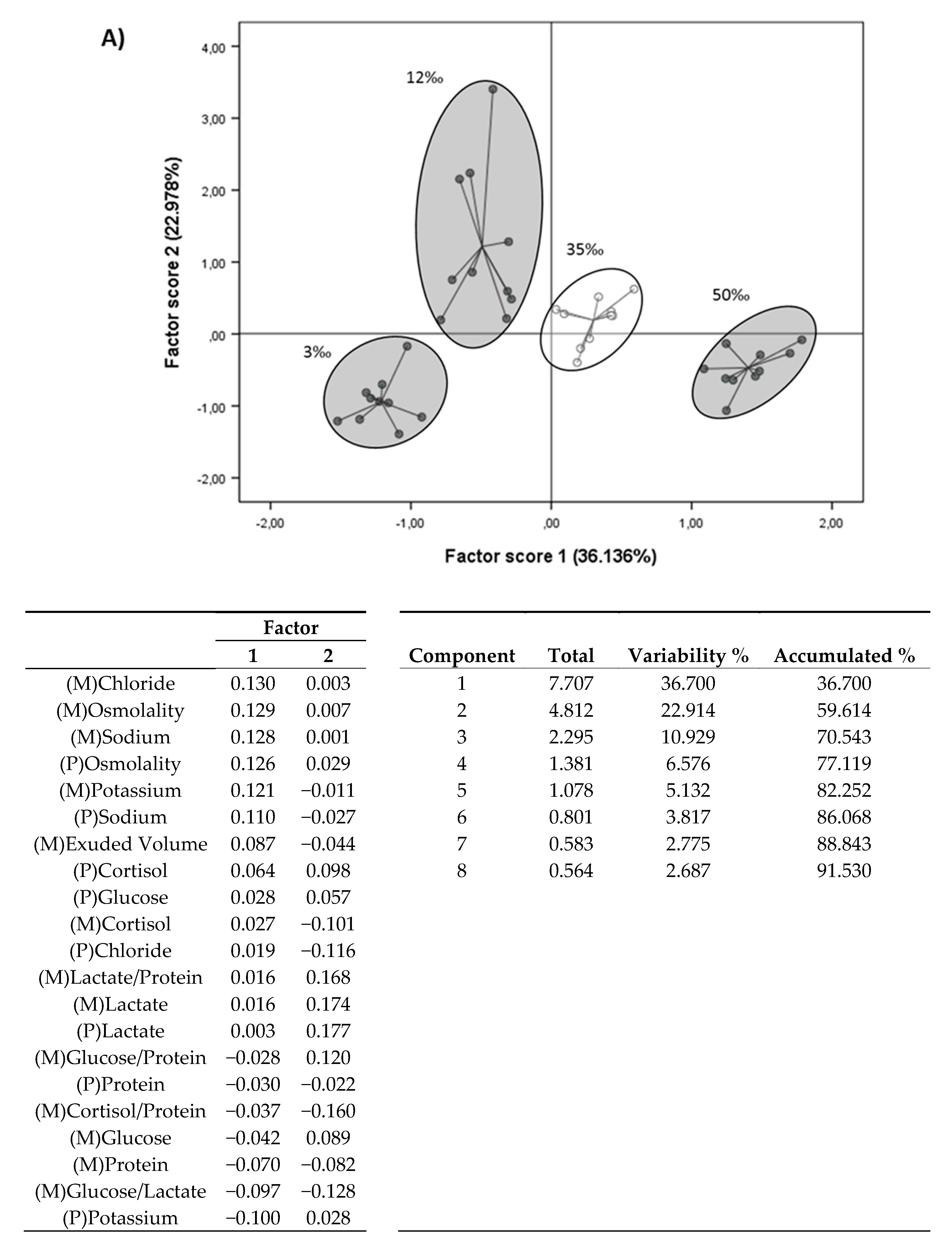
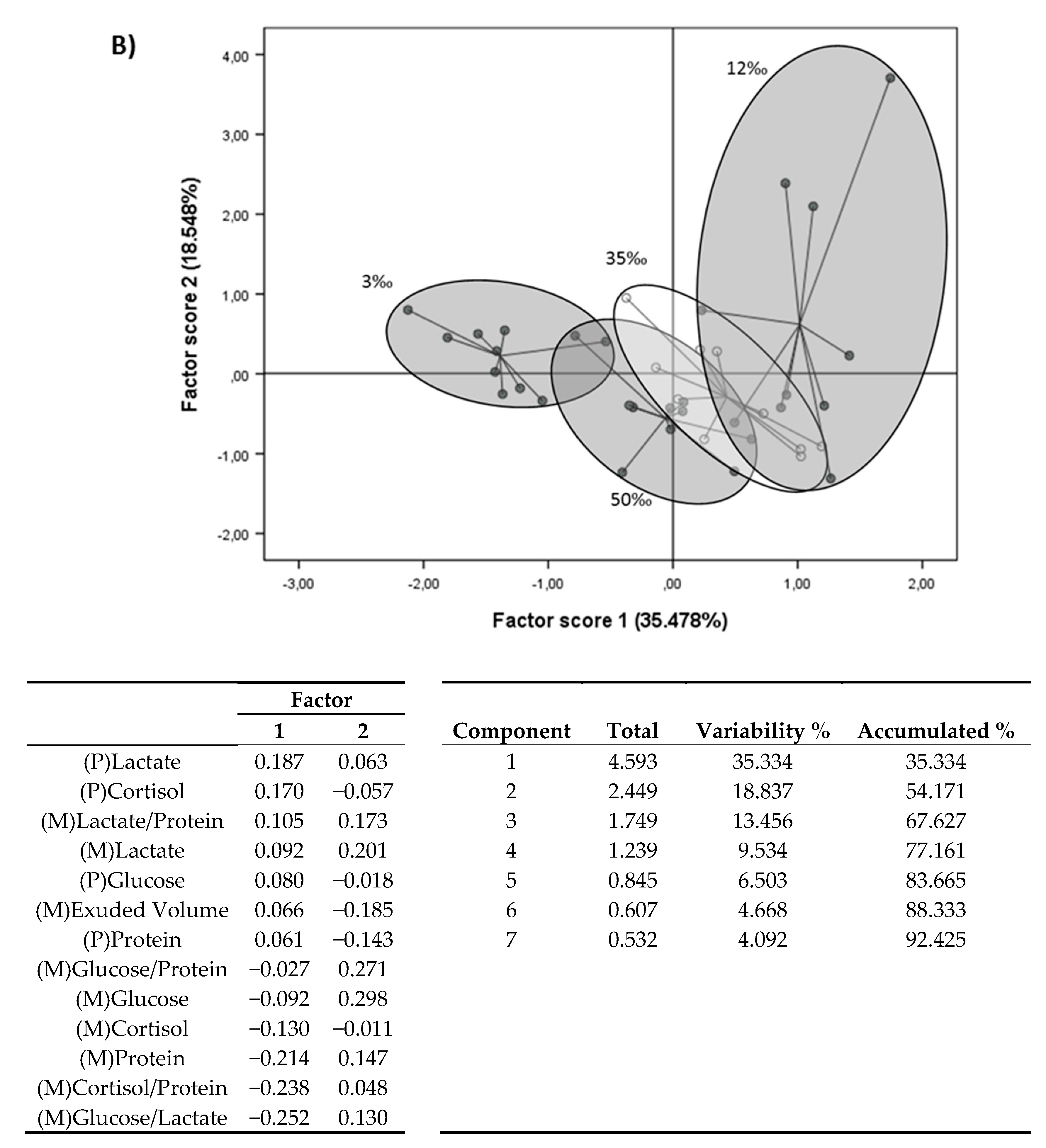
3.3. Principal Component Analysis (PCA)
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Costa, M.J.; Cabral, H.N. Changes in the Tagus nursery function for commercial fish species: Some perspectives for management. Aquat. Ecol. 1999, 33, 287–292. [Google Scholar] [CrossRef]
- Cabral, H.; Costa, M.J. Abundance, feeding ecology and growth of 0-group sea bass, Dicentrarchus labrax, within the nursery areas of the Tagus estuary. J. Mar. Biol. Assoc. UK 2001, 81, 679–682. [Google Scholar] [CrossRef]
- Vasconcelos, R.P.; Reis-Santos, P.; Costa, M.J.; Cabral, H.N. Connectivity between estuaries and marine environment: Integrating metrics to assess estuarine nursery function. Ecol. Indic. 2011, 11, 1123–1133. [Google Scholar] [CrossRef]
- Barnabe, G. Rearing of sea bass larvae (Dicentrarchus labrax (L.); Pisces, Serranidae) using artificial diets. Aquaculture 1976, 9, 237–252. [Google Scholar] [CrossRef]
- Kelley, D.F. The importance of estuaries for sea-bass, Dicentrarchus labrax (L.). J. Fish Biol. 1988, 33, 25–33. [Google Scholar] [CrossRef]
- McDowall, R.M. The evolution of diadromy in fishes (revisited) and its place in phylogenetic analysis. Rev. Fish Biol. Fish. 1997, 7, 443–462. [Google Scholar] [CrossRef]
- Jensen, M.K.; Madsen, S.S.; Kristiansen, R. Osmoregulation and salinity effects on the expression and activity of Na+,K+-ATPase in the gills of European sea bass, Dicentrarchus labrax (L.). J. Exp. Zool. 1998, 282, 290–300. [Google Scholar] [CrossRef]
- Varsamos, S.; Wendelaar Bonga, S.E.; Charmantier, G.; Flik, G. Drinking and Na+/K+ ATPase activity during early development of European sea bass, Dicentrarchus labrax: Ontogeny and short-term regulation following acute salinity changes. J. Exp. Mar. Biol. Ecol. 2004, 311, 189–200. [Google Scholar] [CrossRef]
- Sangiao-Alvarellos, S.; Arjona, F.J.; Martín Del Río, M.P.; Míguez, J.M.; Mancera, J.M.; Soengas, J.L. Time course of osmoregulatory and metabolic changes during osmotic acclimation in Sparus auratus. J. Exp. Biol. 2005, 208, 4291–4304. [Google Scholar] [CrossRef] [Green Version]
- Mylonas, C.C.; Pavlidis, M.; Papandroulakis, N.; Zaiss, M.M.; Tsafarakis, D.; Papadakis, I.E.; Varsamos, S. Growth performance and osmoregulation in the shi drum (Umbrina cirrosa) adapted to different environmental salinities. Aquaculture 2009, 287, 203–210. [Google Scholar] [CrossRef]
- Varsamos, S. Tolerance range and osmoregulation in hypersaline conditions in the European sea bass (Dicentrarchus labrax). J. Mar. Biol. Assoc. UK 2002, 82, 1047–1048. [Google Scholar] [CrossRef]
- Laiz-Carrión, R.; Guerreiro, P.M.; Fuentes, J.; Canario, A.V.M.; Martín Del Río, M.P.; Mancera, J.M. Branchial osmoregulatory response to salinity in the gilthead sea bream. Sparus auratus. J. Exp. Zool. A Comp. Exp. Biol. 2005, 303, 563–576. [Google Scholar] [CrossRef] [PubMed]
- Ellis, T.; Yildiz, H.Y.; López-Olmeda, J.; Spedicato, M.T.; Tort, L.; Øverli, Ø.; Martins, C.I.M. Cortisol and finfish welfare. Fish Physiol. Biochem. 2012, 38, 163–188. [Google Scholar] [CrossRef] [PubMed]
- Balm, P.H.M.; Pepels, P.; Helfrich, S.; Hovens, M.L.M.; Bonga, S.E.W. Adrenocorticotropic hormone in relation to interrenal function during stress in tilapia (Oreochromis mossambicus). Gen. Comp. Endocrinol. 1994, 96, 347–360. [Google Scholar] [CrossRef] [Green Version]
- Barton, B.A. Stress in fishes: A diversity of responses with particular reference to changes in circulating corticosteroids. Integr. Comp. Biol. 2002, 42, 517–525. [Google Scholar] [CrossRef]
- Barton, B.A.; Bollig, H.; Hauskins, B.L.; Jansen, C.R. Juvenile pallid (Scaphirhynchus albus) and hybrid pallidxshovelnose (S. albusxplatorynchus) sturgeons exhibit low physiological responses to acute handling and severe confinement. Comp. Biochem. Physiol. Mol. Integr. Physiol. 2000, 126, 125–134. [Google Scholar] [CrossRef]
- Fernández-Alacid, L.; Sanahuja, I.; Ordóñez-Grande, B.; Sánchez-Nuño, S.; Herrera, M.; Ibarz, A. Skin mucus metabolites and cortisol in meagre fed acute stress-attenuating diets: Correlations between plasma and mucus. Aquaculture 2019, 499, 185–194. [Google Scholar] [CrossRef]
- Fernández-Alacid, L.; Sanahuja, I.; Ordóñez-Grande, B.; Sánchez-Nuño, S.; Herrera, M.; Ibarz, A. Comparison between properties of dorsal and ventral skin mucus in Senegalese sole: Response to an acute stress. Aquaculture 2019, 513, 734410. [Google Scholar] [CrossRef]
- Ruane, N.M.; Wendelaar Bonga, S.E.; Balm, P.H.M. Differences between rainbow trout and brown trout in the regulation of the pituitary-interrenal axis and physiological performance during confinement. Gen. Comp. Endocrinol. 1999, 115, 210–219. [Google Scholar] [CrossRef]
- Schreck, C.B.; Tort, L. The concept of stress in fish. In Fish Physiology; Elsevier Inc.: Amsterdam, The Netherlands, 2016; Volume 35, pp. 1–34. [Google Scholar]
- Wendelaar Bonga, S.E. The stress response in fish. Physiol. Rev. 1997, 77, 591–625. [Google Scholar] [CrossRef]
- Fanouraki, E.; Mylonas, C.C.; Papandroulakis, N.; Pavlidis, M. Species specificity in the magnitude and duration of the acute stress response in Mediterranean marine fish in culture. Gen. Comp. Endocrinol. 2011, 173, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Iwama, G.K.; Vijayan, M.M.; Morgan, J. The stress response in fish in Ichthyology: Recent Research Advances; Saksena, D.N., Ed.; Science Publishers: Enfield, NH, USA, 1999; pp. 47–57. [Google Scholar]
- Mommsen, T.P.; Vijayan, M.M.; Moon, T.W. Cortisol in teleosts: Dynamics, mechanisms of action, and metabolic regulation. Rev. Fish Biol. Fish. 1999, 9, 211–268. [Google Scholar] [CrossRef]
- Laiz-Carrión, R.; Sangiao-Alvarellos, S.; Guzmán, J.M.; Martín Del Río, M.P.; Soengas, J.L.; Mancera, J.M. Growth performance of gilthead sea bream Sparus aurata in different osmotic conditions: Implications for osmoregulation and energy metabolism. Aquaculture 2005, 250, 849–861. [Google Scholar] [CrossRef]
- Sangiao-Alvarellos, S.; Laiz-Carrión, R.; Guzmán, J.M.; Martín del Río, M.P.; Miguez, J.M.; Mancera, J.M.; Soengas, J.L. Acclimation of S. aurata to various salinities alters energy metabolism of osmoregulatory and nonosmoregulatory organs. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2003, 285, 897–907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fanouraki, E.; Papandroulakis, N.; Ellis, T.; Mylonas, C.C.; Scott, A.P.; Pavlidis, M. Water cortisol is a reliable indicator of stress in European sea bass. Dicentrarchus Labrax. Behaviour 2008, 145, 1267–1281. [Google Scholar] [CrossRef]
- Fernández-Alacid, L.; Sanahuja, I.; Ordóñez-Grande, B.; Sánchez-Nuño, S.; Viscor, G.; Gisbert, E.; Herrera, M.; Ibarz, A. Skin mucus metabolites in response to physiological challenges: A valuable non-invasive method to study teleost marine species. Sci. Total Environ. 2018, 644, 1323–1335. [Google Scholar] [CrossRef]
- Sanahuja, I.; Fernández-Alacid, L.; Sánchez-Nuño, S.; Ordóñez-Grande, B.; Ibarz, A. Chronic cold stress alters the skin mucus interactome in a temperate fish model. Front. Physiol. 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Blackstock, N.; Pickering, A.D. Changes in the concentration and histochemistry of epidermal mucous cells during the alevin and fry stages of the Brown trout. Salmo trutta. J. Zool. 1982, 197, 463–471. [Google Scholar] [CrossRef]
- Zaccone, G.; Cascio, P.L.; Fasulo, S.; Licata, A. The effect of an anionic detergent on complex carbohydrates and enzyme activities in the epidermis of the catfish Heteropneustes fossilis (Bloch). Histochem. J. 1985, 17, 453–466. [Google Scholar] [CrossRef]
- Benhamed, S.; Guardiola, F.A.; Mars, M.; Esteban, M.Á. Pathogen bacteria adhesion to skin mucus of fishes. Vet. Microbiol. 2014, 171, 1–12. [Google Scholar] [CrossRef]
- Ibarz, A.; Ordónez-Grande, B.; Sanahuja, I.; Sánchez-Nuno, S.; Fernández-Borras, J.; Blasco, J.; Fernández-Alacid, L. Using stable isotope analysis to study skin mucus exudation and renewal in fish. J. Exp. Biol. 2019, 222. [Google Scholar] [CrossRef] [Green Version]
- Sanahuja, I.; Fernández-Alacid, L.; Ordóñez-Grande, B.; Sánchez-Nuño, S.; Ramos, A.; Araujo, R.M.; Ibarz, A. Comparison of several non-specific skin mucus immune defences in three piscine species of aquaculture interest. Fish Shellfish Immunol. 2019, 89, 428–436. [Google Scholar] [CrossRef]
- Herrera, M.; Fernández-Alacid, L.; Sanahuja, I.; Ibarz, A.; Salamanca, N.; Morales, E.; Giráldez, I. Physiological and metabolic effects of a tryptophan-enriched diet to face up chronic stress in meagre (Argyrosomus regius). Aquaculture 2020, 522, 735102. [Google Scholar] [CrossRef]
- Cordero, H.; Guardiola, F.A.; Tapia-Paniagua, S.T.; Cuesta, A.; Meseguer, J.; Balebona, M.C.; Moriñigo, M.Á.; Esteban, M.Á. Modulation of immunity and gut microbiota after dietary administration of alginate encapsulated Shewanella putrefaciens Pdp11 to gilthead seabream (Sparus aurata L.). Fish Shellfish Immunol. 2015, 45, 608–618. [Google Scholar] [CrossRef]
- Patel, D.M.; Brinchmann, M.F. Skin mucus proteins of lumpsucker (Cyclopterus lumpus). Biochem. Biophys. Rep. 2017, 9, 217–225. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Sánchez, J.; Terova, G.; Simó-Mirabet, P.; Rimoldi, S.; Folkedal, O.; Calduch-Giner, J.A.; Olsen, R.E.; Sitjà-Bobadilla, A. Skin mucus of gilthead sea bream (Sparus aurata L.). protein mapping and regulation in chronically stressed fish. Front. Physiol. 2017, 8, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Rajan, B.; Fernandes, J.M.O.; Caipang, C.M.A.; Kiron, V.; Rombout, J.H.W.M.; Brinchmann, M.F. Proteome reference map of the skin mucus of Atlantic cod (Gadus morhua) revealing immune competent molecules. Fish Shellfish Immunol. 2011, 31, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Sanahuja, I.; Ibarz, A. Skin mucus proteome of gilthead sea bream: A non-invasive method to screen for welfare indicators. Fish Shellfish Immunol. 2015, 46, 426–435. [Google Scholar] [CrossRef]
- Dendrinos, P.; Thorpe, J.P. Effects of reduced salinity on growth and body composition in the European bass Dicentrarchus labrax (L.). Aquaculture 1985, 49, 333–358. [Google Scholar] [CrossRef]
- Varsamos, S.; Connes, R.; Diaz, J.P.; Barnabé, G.; Charmantier, G. Ontogeny of osmoregulation in the European sea bass Dicentrarchus labrax L. Mar. Biol. 2001, 138, 909–915. [Google Scholar] [CrossRef]
- Eroldoǧan, O.T.; Kumlu, M. Growth performance, body traits and fillet composition of the European sea bass (Dicentrarchus labrax) reared in various salinities and fresh water. Turk. J. Vet. Anim. Sci. 2002, 26, 993–1001. [Google Scholar]
- Eroldoǧan, O.T.; Kumlu, M.; Aktaş, M. Optimum feeding rates for European sea bass Dicentrarchus labrax L. reared in seawater and freshwater. Aquaculture 2004, 231, 501–515. [Google Scholar] [CrossRef]
- Sinha, A.K.; Rasoloniriana, R.; Dasan, A.F.; Pipralia, N.; Blust, R.; Boeck, G.D. Interactive effect of high environmental ammonia and nutritional status on ecophysiological performance of European sea bass (Dicentrarchus labrax) acclimated to reduced seawater salinities. Aquat. Toxicol. 2015, 160, 39–56. [Google Scholar] [CrossRef] [PubMed]
- Sinha, A.K.; AbdElgawad, H.; Zinta, G.; Dasan, A.F.; Rasoloniriana, R.; Asard, H.; Blust, R.; De Boeck, G. Nutritional status as the key modulator of antioxidant responses induced by high environmental ammonia and salinity stress in European sea bass (Dicentrarchus labrax). PLoS ONE 2015, 10, e0135091. [Google Scholar] [CrossRef] [Green Version]
- Guardiola, F.A.; Cuesta, A.; Esteban, M.Á. Using skin mucus to evaluate stress in gilthead seabream (Sparus aurata L.). Fish Shellfish Immunol. 2016, 59, 323–330. [Google Scholar] [CrossRef]
- De Mercado, E.; Larrán, A.M.; Pinedo, J.; Tomás-Almenar, C. Skin mucous: A new approach to assess stress in rainbow trout. Aquaculture 2018, 484, 90–97. [Google Scholar] [CrossRef]
- Shephard, K.L. Functions for fish mucus. Rev. Fish Biol. Fish. 1994, 4, 401–429. [Google Scholar] [CrossRef]
- Franklin, C.E. Surface ultrastructural changes in the gills of sockeye salmon (teleostei: Oncorhynchus nerka) during seawater transfer: Comparison of successful and unsuccessful seawater adaptation. J. Morphol. 1990, 206, 13–23. [Google Scholar] [CrossRef]
- Abraham, M.; Iger, Y.; Zhang, L. Fine structure of the skin cells of a stenohaline freshwater fish Cyprinus carpio exposed to diluted seawater. Tissue Cell 2001, 33, 46–54. [Google Scholar] [CrossRef]
- Roberts, S.D.; Powell, M.D. Comparative ionic flux and gill mucous cell histochemistry: Effects of salinity and disease status in Atlantic salmon (Salmo salar L.). Comp. Biochem. Physiol. Mol. Integr. Physiol. 2003, 134, 525–537. [Google Scholar] [CrossRef]
- DiMaggio, M.A.; Ohs, C.L.; Petty, B.D. Salinity tolerance of the Seminole killifish, Fundulus seminolis, a candidate species for marine baitfish aquaculture. Aquaculture 2009, 293, 74–80. [Google Scholar] [CrossRef]
- Fiúza, L.S.; Aragão, N.M.; Ribeiro Junior, H.P.; de Moraes, M.G.; Rocha, Í.R.C.B.; Lustosa Neto, A.D.; de Sousa, R.R.; Madrid, R.M.M.; de Oliveira, E.G.; Costa, F.H.F. Effects of salinity on the growth, survival, haematological parameters and osmoregulation of tambaqui Colossoma macropomum juveniles. Aquac. Res. 2015, 46, 1–9. [Google Scholar] [CrossRef]
- Burden, C.E. The failure of hypophysectomized fundulus heteroclitus to survive in fresh water. Biol. Bull. 1956, 110, 8–28. [Google Scholar] [CrossRef]
- Ahuja, S.K. Chloride-cell and mucus cell response to chloride and sulphate-enriched media in the gills of Gambusia affinis affinis (Barid and Girard) and Catla catla (Hamilton). J. Exp. Zool. 1970, 173, 231–249. [Google Scholar] [CrossRef]
- Wendelaar Bonga, S.E. The role of environmental calcium and magnesium ions in the control of prolactin secretion in the teleost Gasterosteus aculeatus. In Comparative Endocrinology; Gaillard, P.J., Boer, H.H., Eds.; Elseiver/North-Holland Biomedical Press: Amsterdam, The Netherlands, 1978; pp. 259–262. [Google Scholar]
- Scott, A.; Pinillos, M.; Ellis, T. Why measure steroids in fish plasma when you can measure them in water. In Perspectives in Comparative Endocrinology: Unity and Diversity, Proceedings of the 14th International Congress of Comparative Endocrinology, Sorrento, Italy, 26–30 May 2001; Monduzzi Editore: Bologna, Italy, 2001; pp. 1291–1295. [Google Scholar]
- Scott, A.P.; Ellis, T. Measurement of fish steroids in water-a review. Gen. Comp. Endocrinol. 2007, 153, 392–400. [Google Scholar] [CrossRef]
- Ruane, N.M.; Komen, H. Measuring cortisol in the water as an indicator of stress caused by increased loading density in common carp (Cyprinus carpio). Aquaculture 2003, 218, 685–693. [Google Scholar] [CrossRef]
- Bertotto, D.; Poltronieri, C.; Negrato, E.; Majolini, D.; Radaelli, G.; Simontacchi, C. Alternative matrices for cortisol measurement in fish. Aquac. Res. 2010, 41, 1261–1267. [Google Scholar] [CrossRef]
- Gesto, M.; Hernández, J.; López-Patiño, M.A.; Soengas, J.L.; Míguez, J.M. Is gill cortisol concentration a good acute stress indicator in fish? A study in rainbow trout and zebrafish. Comp. Biochem. Physiol. Mol. Integr. Physiol. 2015, 188, 65–69. [Google Scholar] [CrossRef]
- Martínez-Porchas, M.; Martínez-Córdova, L.R.; Ramos-Enriquez, R. Cortisol and glucose: Reliable indicators of fish stress? Pan Am. J. Aquat. Sci. 2009, 4, 158–178. [Google Scholar]
- Pankhurst, N.W. The endocrinology of stress in fish: An environmental perspective. Gen. Comp. Endocrinol. 2011, 170, 265–275. [Google Scholar] [CrossRef] [Green Version]
- Concollato, A.; Olsen, R.E.; Vargas, S.C.; Bonelli, A.; Cullere, M.; Parisi, G. Effects of stunning/slaughtering methods in rainbow trout (Oncorhynchus mykiss) from death until rigor mortis resolution. Aquaculture 2016, 464, 74–79. [Google Scholar] [CrossRef]
- Omlin, T.; Weber, J.M. Hypoxia stimulates lactate disposal in rainbow trout. J. Exp. Biol. 2010, 213, 3802–3809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, P.P.; Sun, C.M.; Wu, S.M. Changes of plasma osmolality, chloride concentration and gill Na-K-ATPase activity in tilapia Oreochromis mossambicus during seawater acclimation. Mar. Biol. 1989, 100, 295–299. [Google Scholar] [CrossRef]
- Walker, R.L.; Wilkes, P.R.H.; Wood, C.M. The effects of hypersaline exposure on oxygen-affinity of the blood of the freshwater teleost Catostomus Commersoni. J. Exp. Biol. 1989, 142, 125–142. [Google Scholar]
- Altinok, I.; Galli, S.M.; Chapman, F.A. Ionic and osmotic regulation capabilities of juvenile Gulf of Mexico sturgeon, Acipenser oxyrinchus de sotoi. Comp. Biochem. Physiol. Mol. Integr. Physiol. 1998, 120, 609–616. [Google Scholar] [CrossRef]
- LeBreton, G.T.O.; Beamish, F.W.H. The influence of salinity on ionic concentrations and osmolarity of blood serum in lake sturgeon, Acipenser fulvescens. Environ. Biol. Fishes 1998, 52, 477–482. [Google Scholar] [CrossRef]
- Evans, D.H.; Piermarini, P.M.; Potts, W.T.W. Ionic transport in the fish gill epithelium. J. Exp. Zool. 1999, 283, 641–652. [Google Scholar] [CrossRef]
- Greenwell, M.G.; Sherrill, J.; Clayton, L.A. Osmoregulation in fish mechanisms and clinical implications. Vet. Clin. North Am. Exot. Anim. Pract. 2003, 6, 169–189. [Google Scholar] [CrossRef]
- Roberts, S.D.; Powell, M.D. The viscosity and glycoprotein biochemistry of salmonid mucus varies with species, salinity and the presence of amoebic gill disease. J. Comp. Physiol. B 2005, 175, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shephard, K.L. The influence of mucus on the diffusion of water across fish epidermis. Physiol. Zool. 1981, 54, 224–229. [Google Scholar] [CrossRef]
- Marshall, W.S. On the involvement of mucous secretion in teleost osmoregulation. Can. J. Zool. 1978, 56, 1088–1091. [Google Scholar] [CrossRef]
- Kirschner, L.B. External charged layer and Na+ regulation. In Osmotic and Volume Regulation, Proceedings of the Alfred Benzon Symposium XI, Copenhagen, Denmark, 5–9 June 1977; Barker Jørgensen, C., Skadhauge, E., Eds.; Munksgaard: Copenhagen, Denmark, 1977. [Google Scholar]
- Verdugo, P. Hydration kinetics of exocytosed mucins in cultured secretory cells of the rabbit trachea: A new model. In Ciba Foundation Symposium 109—Mucus and Mucosa; Nugent, J., O’Connor, M., Eds.; Wiley: Hoboken, NJ, USA, 1984. [Google Scholar]
- Zuchelkowski, E.M.; Pinkstaff, C.A.; Hinton, D.E. Mucosubstance histochemistry in control and acid-stressed epidermis of brown bullhead catfish, lctalurus nebulosus (LeSueur). Anat. Rec. 1985, 212, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Handy, R.D. The ionic composition of rainbow trout body mucus. Comp. Biochem. Physiol. Physiol. 1989, 93, 571–575. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Salinity Challenge | 3‰ | 12‰ | 35‰ | 50‰ | ||||
| Exudation parameters | ||||||||
| Collected Mucus (μL) | 122.22 ± 8.78 | b | 120.00 ± 28.09 | ab | 150.00 ± 1.51 | a | 266.67 ± 33.33 | c |
| Exuded mucus/skin (μL/cm2) | 1.95 ± 0.16 | a | 1.73 ± 0.36 | a | 2.68 ± 0.28 | a | 4.82 ± 0.74 | b |
| Exuded mucus/bw (μL/g) | 0.93 ± 0.08 | b | 0.83 ± 0.17 | ab | 1.28 ± 0.13 | a | 2.31 ± 0.35 | c |
| Salinity Challenge | 3‰ | 12‰ | 35‰ | 50‰ | ||||
| Mucus biomarkers | ||||||||
| Glucose (μg/mL) | 35.31 ± 3.81 | ab | 41.65 ± 7.18 | b | 28.53 ± 4.19 | ab | 25.35 ± 2.58 | a |
| Lactate (μg/mL) | 3.33 ± 0.55 | a | 25.03 ± 7.84 | b | 9.17 ± 0.84 | a | 8.99 ± 1.38 | a |
| Cortisol (ng/mL) | 9.07 ± 2.42 | ab | 4.50 ± 0.98 | a | 4.25 ± 1.16 | a | 11.52 ± 0.54 | b |
| Soluble protein (mg/mL) | 6.96 ± 0.47 | b | 5.04 ± 0.44 | a | 5.14 ± 0.37 | a | 5.08 ± 0.31 | a |
| Glucose/Protein (μg/mg) | 5.29 ± 0.49 | 7.67 ± 1.24 | 5.47 ± 0.63 | 4.67 ± 0.46 | ||||
| Lactate/Protein (μg/mg) | 0.40 ± 0.04 | b | 3.45 ± 0.90 | ab | 1.96 ± 0.19 | a | 1.45 ± 0.11 | a |
| Glucose/Lactate (μg/μg) | 14.36 ± 1.98 | b | 2.09 ± 0.28 | a | 3.04 ± 0.44 | a | 3.05 ± 0.25 | a |
| Cortisol/Protein (ng/mg) | 1.40 ± 0.34 | ab | 0.85 ± 0.09 | a | 0.83 ± 0.21 | a | 2.32 ± 0.38 | b |
| Plasma Biomarkers | Salinity Challenge | Plasma vs. Mucus 1 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 3‰ | 12‰ | 35‰ | 50‰ | R Coefficient | p-Value | |||||
| Glucose (mg/dL) | 173.86 ± 17.64 | 186.22 ± 7.92 | 184.88 ± 11.23 | 188.22 ± 15.33 | 0.07 | >0.05 | ||||
| Lactate (mg/dL) | 35.41 ± 2.45 b | b | 105.51 ± 8.13 | c | 66.39 ± 7.64 | a | 39.86 ± 3.92 | b | 0.69 | <0.01 |
| Cortisol (ng/mL) | 333.77 ± 101.67 | 615.88 ± 102.08 | 453.64 ± 80.99 | 586.66 ± 154.32 | −0.05 | >0.05 | ||||
| Protein (mg/mL) | 20.70 ± 0.75 | 21.18 ± 1.02 | 21.14 ± 0.81 | 18.80 ± 0.95 | 0.07 | >0.05 | ||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ordóñez-Grande, B.; Guerreiro, P.M.; Sanahuja, I.; Fernández-Alacid, L.; Ibarz, A. Evaluation of an Acute Osmotic Stress in European Sea Bass via Skin Mucus Biomarkers. Animals 2020, 10, 1546. https://doi.org/10.3390/ani10091546
Ordóñez-Grande B, Guerreiro PM, Sanahuja I, Fernández-Alacid L, Ibarz A. Evaluation of an Acute Osmotic Stress in European Sea Bass via Skin Mucus Biomarkers. Animals. 2020; 10(9):1546. https://doi.org/10.3390/ani10091546
Chicago/Turabian StyleOrdóñez-Grande, Borja, Pedro M. Guerreiro, Ignasi Sanahuja, Laura Fernández-Alacid, and Antoni Ibarz. 2020. "Evaluation of an Acute Osmotic Stress in European Sea Bass via Skin Mucus Biomarkers" Animals 10, no. 9: 1546. https://doi.org/10.3390/ani10091546