A Novel Nanobody Directed against Ovine Myostatin to Enhance Muscle Growth in Mouse

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. SDS-PAGE Analysis
2.2. ELISA
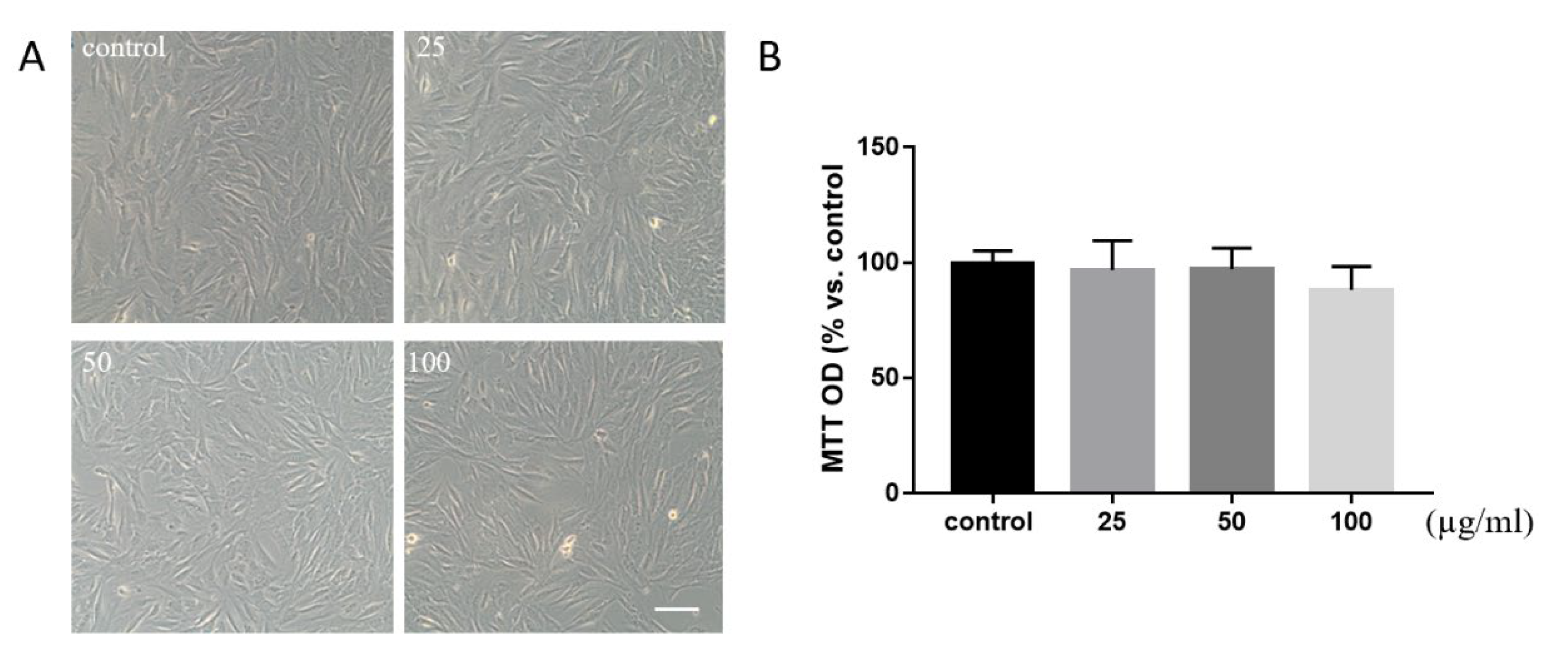
2.3. Cell Culture and Viability
2.4. Animal Experiments
2.5. Immunofluorescence and Hematoxylin–Eosin (H&E) Staining
2.6. Statistics
3. Results
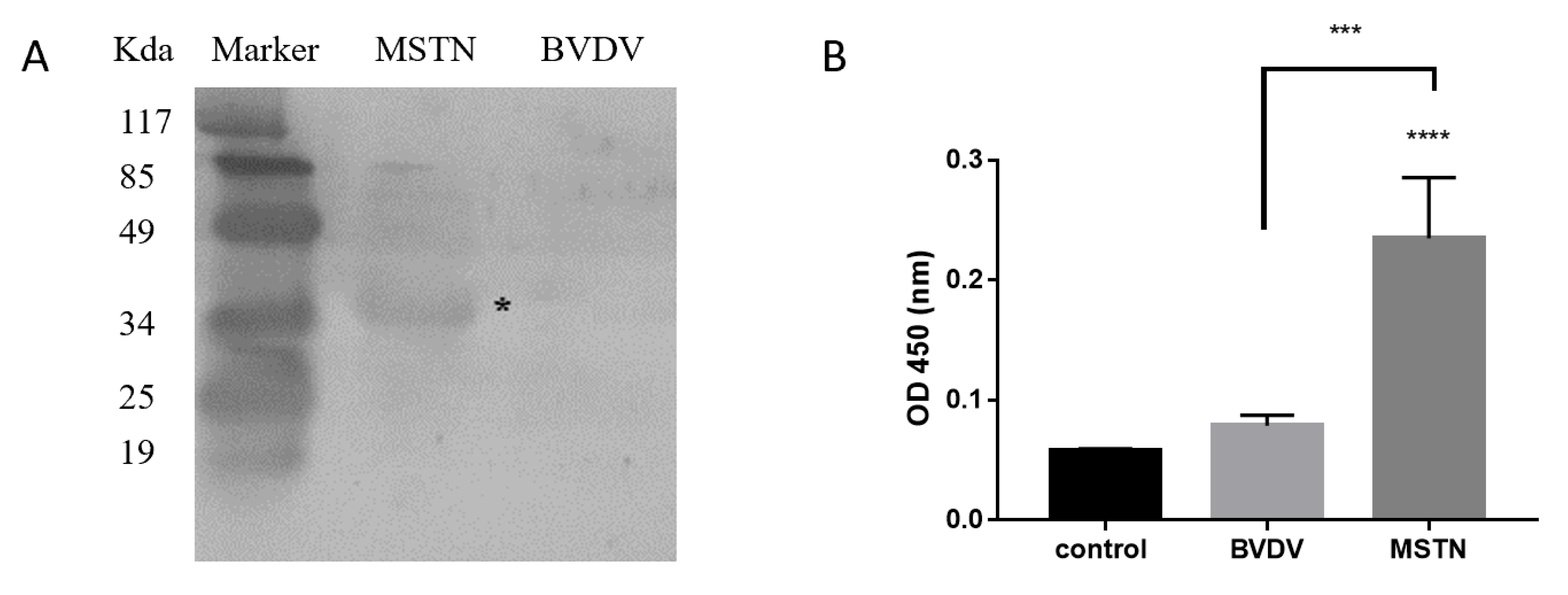
3.1. Specific Active Detection of RMN
3.2. Evaluation of RMN
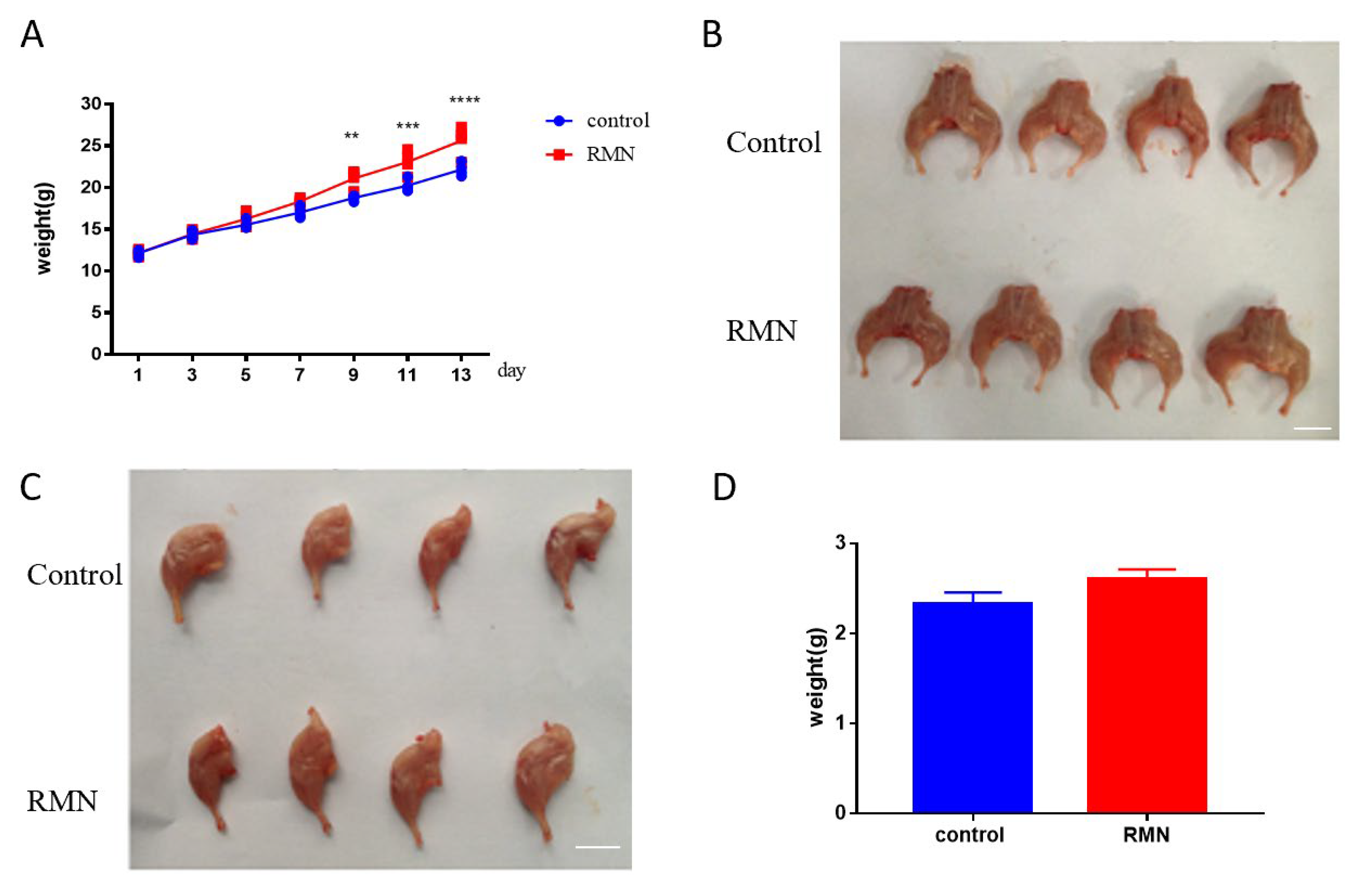
3.3. Effect of RMN Injection on Body and Hind Leg Weight in Mice
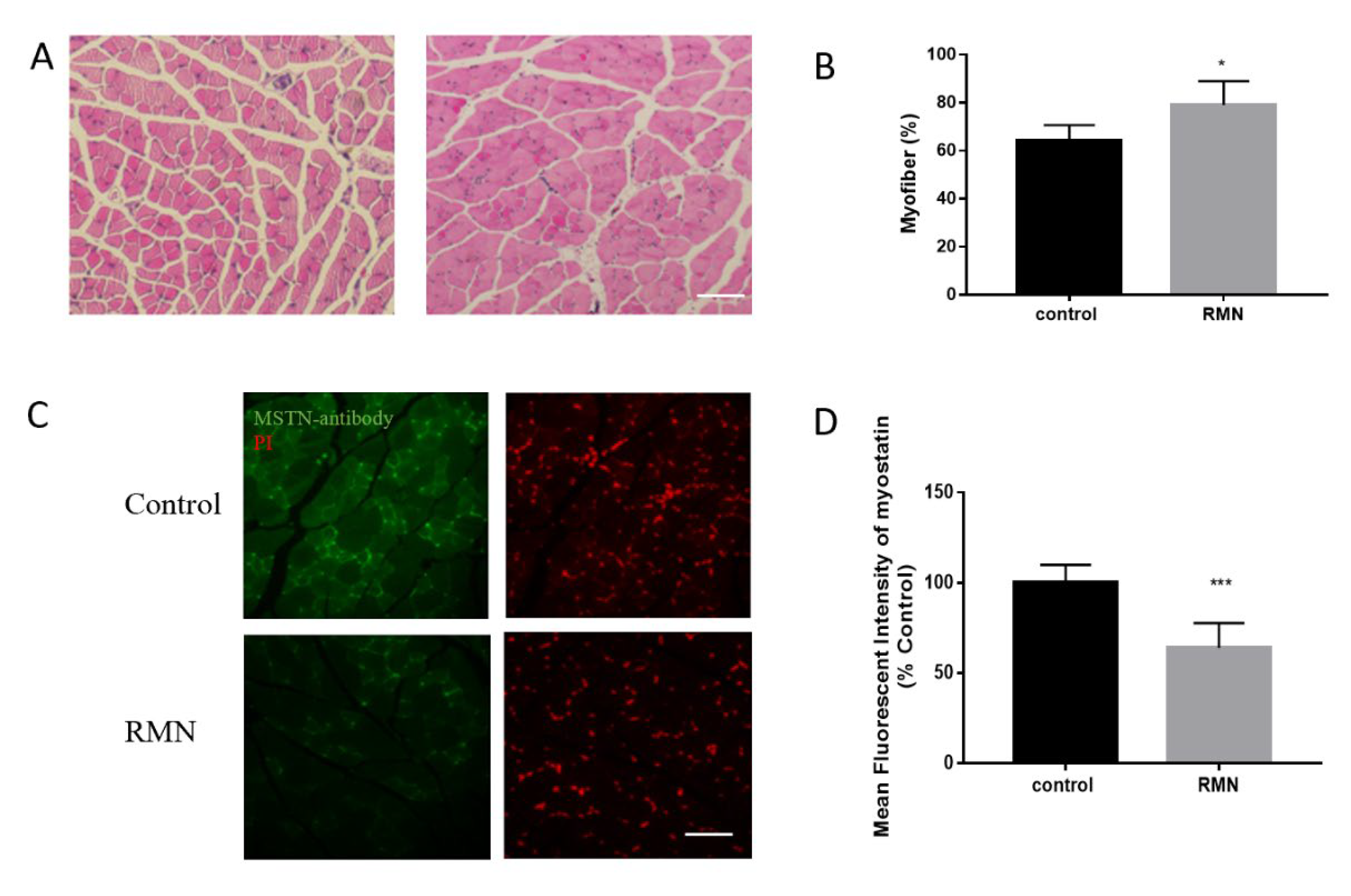
3.4. RMN Increases Muscle Fiber and Inhibits MSTN Protein
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Matsakas, A.; Diel, P. The growth factor myostatin, a key regulator in skeletal muscle growth and homeostasis. Int. J. Sports Med. 2005, 26, 83–89. [Google Scholar] [CrossRef] [PubMed]
- McPherron, A.C.; Lee, S.J. Double muscling in cattle due to mutations in the myostatin gene. Proc. Natl. Acad. Sci. USA 1997, 94, 12457–12461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clop, A.; Marcq, F.; Takeda, H.; Pirottin, D.; Tordoir, X.; Bibé, B.; Bouix, J.; Caiment, F.; Elsen, J.M.; Eychenne, F.; et al. A mutation creating a potential illegitimate microRNA target site in the myostatin gene affects muscularity in sheep. Nat. Genet. 2006, 38, 813–818. [Google Scholar] [CrossRef] [PubMed]
- Mosher, D.S.; Quignon, P.; Bustamante, C.D.; Sutter, N.B.; Mellersh, C.S.; Parker, H.G.; Ostrander, E.A. A mutation in the myostatin gene increases muscle mass and enhances racing performance in heterozygote dogs. PLoS Genet. 2007, 3, e79. [Google Scholar] [CrossRef]
- Williams, M.S. Myostatin mutation associated with gross muscle hypertrophy in a child. N. Engl. J. Med. 2004, 351, 1030–1031. [Google Scholar]
- Zhu, X.; Hadhazy, M.; Wehling, M.; Tidball, J.G.; McNally, E.M. Dominant negative myostatin produces hypertrophy without hyperplasia in muscle. FEBS Lett. 2000, 474, 71–75. [Google Scholar] [CrossRef]
- Chen, P.R.; Lee, K. INVITED REVIEW: Inhibitors of myostatin as methods of enhancing muscle growth and development. J. Anim. Sci. 2016, 94, 3125–3134. [Google Scholar] [CrossRef] [Green Version]
- Joulia-Ekaza, D.; Cabello, G. Myostatin regulation of muscle development: Molecular basis, natural mutations, physiopathological aspects. Exp. Cell Res. 2006, 312, 2401–2414. [Google Scholar] [CrossRef]
- Aiello, D.; Patel, K.; Lasagna, E. The myostatin gene: An overview of mechanisms of action and its relevance to livestock animals. Anim. Genet. 2018, 49, 505–519. [Google Scholar] [CrossRef] [Green Version]
- McPherron, A.C.; Lawler, A.M.; Lee, S.J. Regulation of skeletal muscle mass in mice by a new TGF-beta superfamily member. Nature 1997, 387, 83–90. [Google Scholar] [CrossRef]
- Hu, S.; Ni, W.; Sai, W.; Zi, H.; Qiao, J.; Wang, P.; Sheng, J.; Chen, C. Knockdown of myostatin expression by RNAi enhances muscle growth in transgenic sheep. PLoS ONE 2013, 8, e58521. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Chen, C.; Sheng, J.; Sun, Y.; Cao, X.; Qiao, J. Enhanced muscle growth by plasmid-mediated delivery of myostatin propeptide. J. Biomed. Biotechnol. 2010, 2010, 862591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saeed, A.F.; Wang, R.; Ling, S.; Wang, S. Antibody Engineering for Pursuing a Healthier Future. Front. Microbiol. 2017, 8, 495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruigrok, V.J.; Levisson, M.; Eppink, M.H.; Smidt, H.; van der Oost, J. Alternative affinity tools: More attractive than antibodies? Biochem. J. 2011, 436, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Hamers-Casterman, C.; Atarhouch, T.; Muyldermans, S.; Robinson, G.; Hamers, C.; Songa, E.B.; Bendahman, N.; Hamers, R. Naturally occurring antibodies devoid of light chains. Nature 1993, 363, 446–448. [Google Scholar] [CrossRef]
- Salvador, J.P.; Vilaplana, L.; Marco, M.P. Nanobody: Outstanding features for diagnostic and therapeutic applications. Anal. Bioanal. Chem. 2019, 411, 1703–1713. [Google Scholar] [CrossRef]
- Siontorou, C.G. Nanobodies as novel agents for disease diagnosis and therapy. Int. J. Nanomed. 2013, 8, 4215–4227. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Fan, Z.; Shao, L.; Kong, X.; Hou, X.; Tian, D.; Sun, Y.; Xiao, Y.; Yu, L. Nanobody-derived nanobiotechnology tool kits for diverse biomedical and biotechnology applications. Int. J. Nanomed. 2016, 11, 3287–3303. [Google Scholar] [CrossRef] [Green Version]
- Manglik, A.; Kobilka, B.K.; Steyaert, J. Nanobodies to Study G Protein-Coupled Receptor Structure and Function. Annu. Rev. Pharmacol. Toxicol. 2017, 57, 19–37. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.J.; Zhang, G.Q.; Guo, J.X.; Chen, X.K.; Dou, X.X.; Chen, C.F.; Sheng, J.L. Expression of Ovine Myostatin Gene and Construction and Identification of Nanobody Library Against Recombinant MSTN. Chin. Biotechnol. 2014, 34, 87–93. [Google Scholar] [CrossRef]
- Zhang, G.Q.; Li, Y.J.; Xiao, H.R.; Huang, M.L.; Yang, X.; Chen, C.F.; Sheng, J.L. Construction and Characterization of a Nanobody Library against BVDV. Acta Agricu. Boreali-Occident. Sin. 2015, 24, 20–25. [Google Scholar] [CrossRef]
- Li, Z.; Zhao, B.; Kim, Y.S.; Hu, C.Y.; Yang, J. Administration of a mutated myostatin propeptide to neonatal mice significantly enhances skeletal muscle growth. Mol. Reprod. Dev. 2010, 77, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Ma, D.; Jiang, S.; Gao, P.; Qian, L.; Wang, Q.; Cai, C.; Xiao, G.; Yang, J.; Cui, W. Functional verification of a porcine myostatin propeptide mutant. Transgenic Res. 2015, 24, 837–845. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Li, C.; Liu, Z.; Ni, W.; Yao, R.; Xu, Y.; Quan, R.; Zhang, M.; Li, H.; Liu, L.; et al. Circular RNA circ-FoxO3 Inhibits Myoblast Cells Differentiation. Cells 2019, 8, 616. [Google Scholar] [CrossRef] [Green Version]
- Milazzotto, M.P.; Goissis, M.D.; Feitosa, W.B.; Martins, L.F.; Strauss, B.E.; Bajgelman, M.C.; Assumpção, M.E.; Visintin, J.A. Myostatin gene knockdown through lentiviral-mediated delivery of shRNA for in vitro production of transgenic bovine embryos. Zygote 2010, 18, 339–344. [Google Scholar] [CrossRef]
- Rodino-Klapac, L.R.; Haidet, A.M.; Kota, J.; Handy, C.; Kaspar, B.K.; Mendell, J.R. Inhibition of myostatin with emphasis on follistatin as a therapy for muscle disease. Muscle Nerve Off. J. Am. Assoc. Electrodiagn. Med. 2009, 39, 283–296. [Google Scholar] [CrossRef] [Green Version]
- Hu, S.; Ni, W.; Sai, W.; Zhang, H.; Cao, X.; Qiao, J.; Sheng, J.; Guo, F.; Chen, C. Sleeping Beauty-mediated knockdown of sheep myostatin by RNA interference. Biotechnol. Lett. 2011, 33, 1949–1953. [Google Scholar] [CrossRef]
- Whittemore, L.A.; Song, K.; Li, X.; Aghajanian, J.; Davies, M.; Girgenrath, S.; Hill, J.J.; Jalenak, M.; Kelley, P.; Knight, A.; et al. Inhibition of myostatin in adult mice increases skeletal muscle mass and strength. Biochem. Biophys. Res. Commun. 2003, 300, 965–971. [Google Scholar] [CrossRef]
- Huang, L.; Muyldermans, S.; Saerens, D. Nanobodies®: Proficient tools in diagnostics. Expert Rev. Mol. Diagn. 2010, 10, 777–785. [Google Scholar] [CrossRef]
- Muyldermans, S.; Baral, T.N.; Retamozzo, V.C.; De Baetselier, P.; De Genst, E.; Kinne, J.; Leonhardt, H.; Magez, S.; Nguyen, V.K.; Revets, H.; et al. Camelid immunoglobulins and nanobody technology. Vet. Immunol. Immunopathol. 2009, 128, 178–183. [Google Scholar] [CrossRef] [Green Version]
- De Meyer, T.; Muyldermans, S.; Depicker, A. Nanobody-based products as research and diagnostic tools. Trends Biotechnol. 2014, 32, 263–270. [Google Scholar] [CrossRef] [PubMed]
- De Groeve, K.; Deschacht, N.; De Koninck, C.; Caveliers, V.; Lahoutte, T.; Devoogdt, N.; Muyldermans, S.; De Baetselier, P.; Raes, G. Nanobodies as tools for in vivo imaging of specific immune cell types. J. Nucl. Med. Off. Publ. Soc. Nucl. Med. 2010, 51, 782–789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Mukhtar, H.; Ma, L.; Pang, Q.; Wang, X. VHH Antibodies: Reagents for Mycotoxin Detection in Food Products. Sensors 2018, 18, 485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, W.; Song, H.; Chen, Q.; Yu, J.; Xian, M.; Nian, R.; Feng, D. Recent advances in the selection and identification of antigen-specific nanobodies. Mol. Immunol. 2018, 96, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Crivianu-Gaita, V.; Thompson, M. Aptamers, antibody scFv, and antibody Fab’ fragments: An overview and comparison of three of the most versatile biosensor biorecognition elements. Biosens. Bioelectron. 2016, 85, 32–45. [Google Scholar] [CrossRef] [PubMed]
- Ingram, J.R.; Schmidt, F.I.; Ploegh, H.L. Exploiting Nanobodies’ Singular Traits. Annu. Rev. Immunol. 2018, 36, 695–715. [Google Scholar] [CrossRef]
- Iezzi, M.E.; Policastro, L.; Werbajh, S.; Podhajcer, O.; Canziani, G.A. Single-Domain Antibodies and the Promise of Modular Targeting in Cancer Imaging and Treatment. Front. Immunol. 2018, 9, 273. [Google Scholar] [CrossRef] [Green Version]
- Steeland, S.; Vandenbroucke, R.E.; Libert, C. Nanobodies as therapeutics: Big opportunities for small antibodies. Drug Discov. Today 2016, 21, 1076–1113. [Google Scholar] [CrossRef]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ou, K.; Li, Y.; Wu, P.; Guo, J.; Hao, X.; Sheng, J.; Chen, C. A Novel Nanobody Directed against Ovine Myostatin to Enhance Muscle Growth in Mouse. Animals 2020, 10, 1398. https://doi.org/10.3390/ani10081398
Ou K, Li Y, Wu P, Guo J, Hao X, Sheng J, Chen C. A Novel Nanobody Directed against Ovine Myostatin to Enhance Muscle Growth in Mouse. Animals. 2020; 10(8):1398. https://doi.org/10.3390/ani10081398
Chicago/Turabian StyleOu, Kepeng, Youjian Li, Peng Wu, Jixing Guo, Xiujing Hao, Jinliang Sheng, and Chuangfu Chen. 2020. "A Novel Nanobody Directed against Ovine Myostatin to Enhance Muscle Growth in Mouse" Animals 10, no. 8: 1398. https://doi.org/10.3390/ani10081398