Attention Bias Test Measures Negative But Not Positive Affect in Sheep: A Replication Study


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Animal Details
2.3. Experimental Design
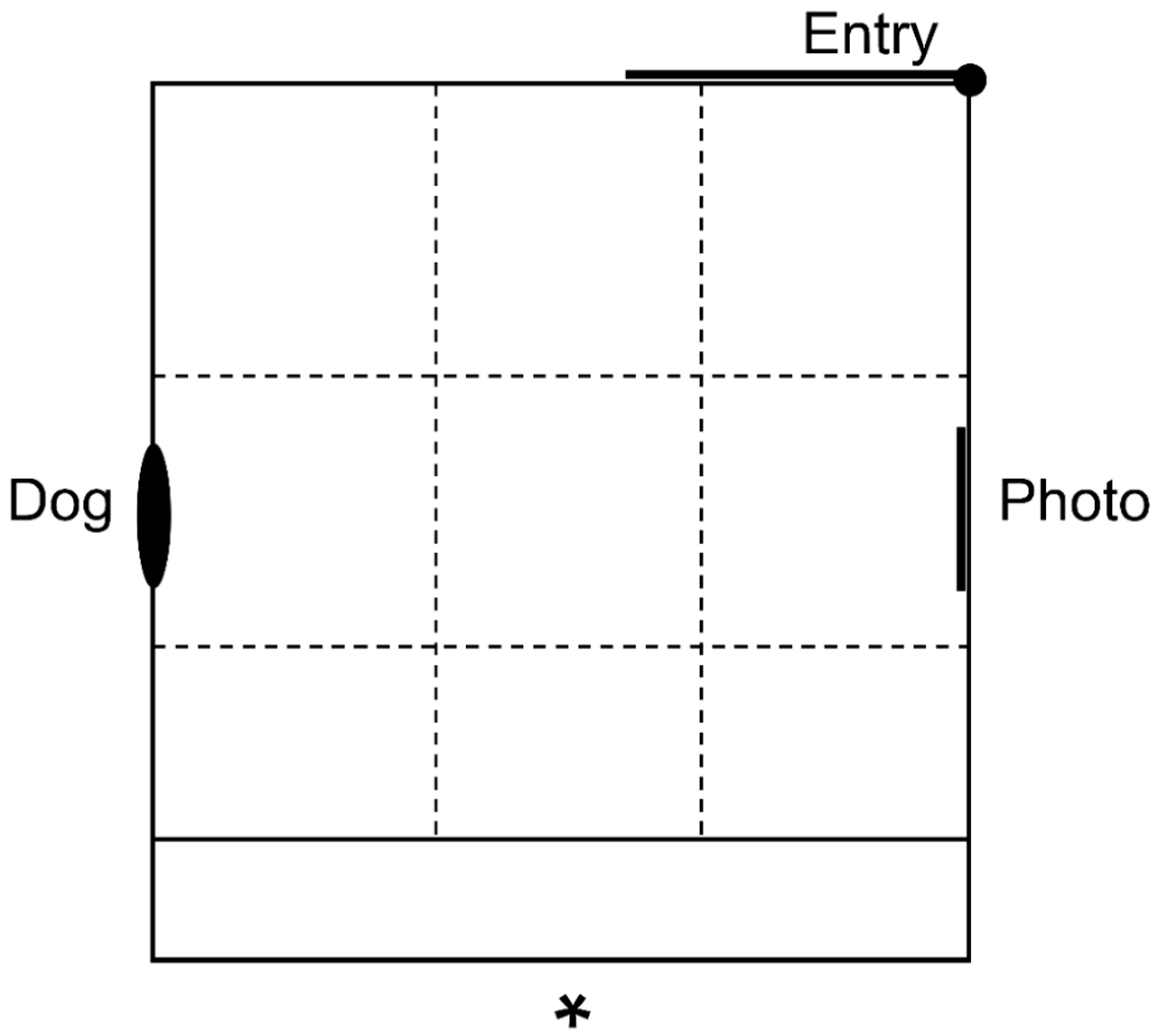
2.4. Attention Bias Test Method
2.5. Body Temperature
2.6. Statistical Methods
3. Results
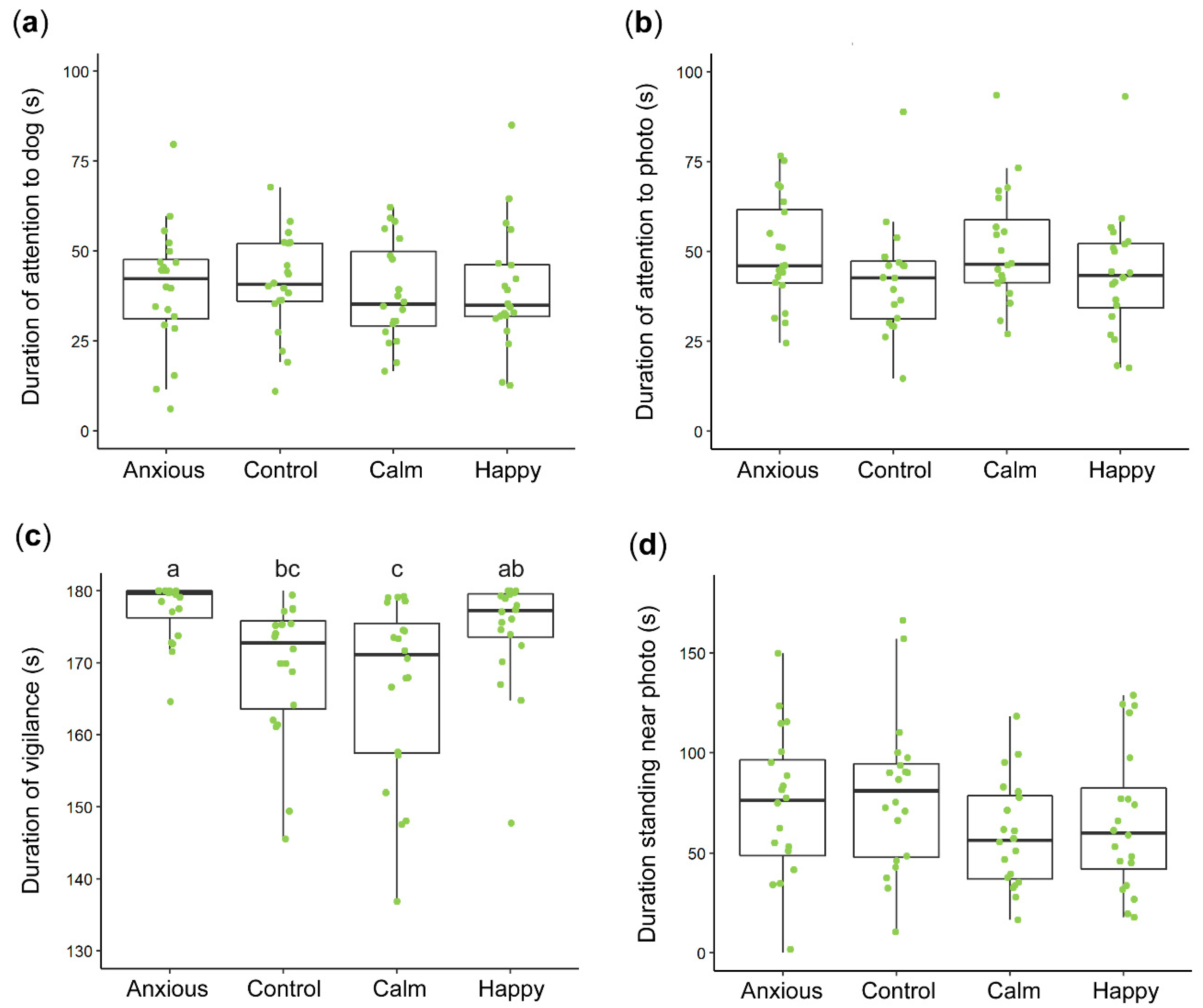
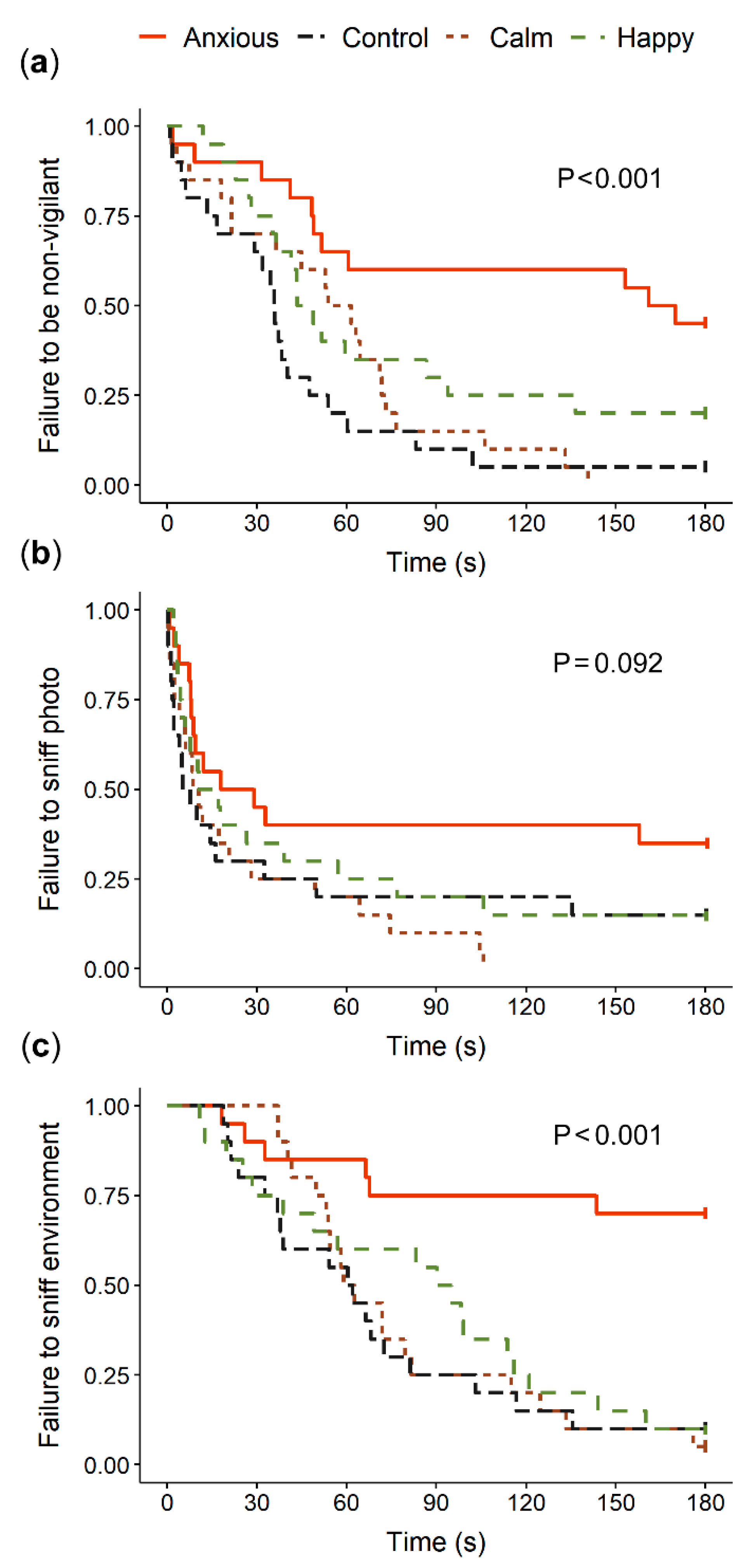
3.1. Vigilance and Attention
3.2. Other Behaviors
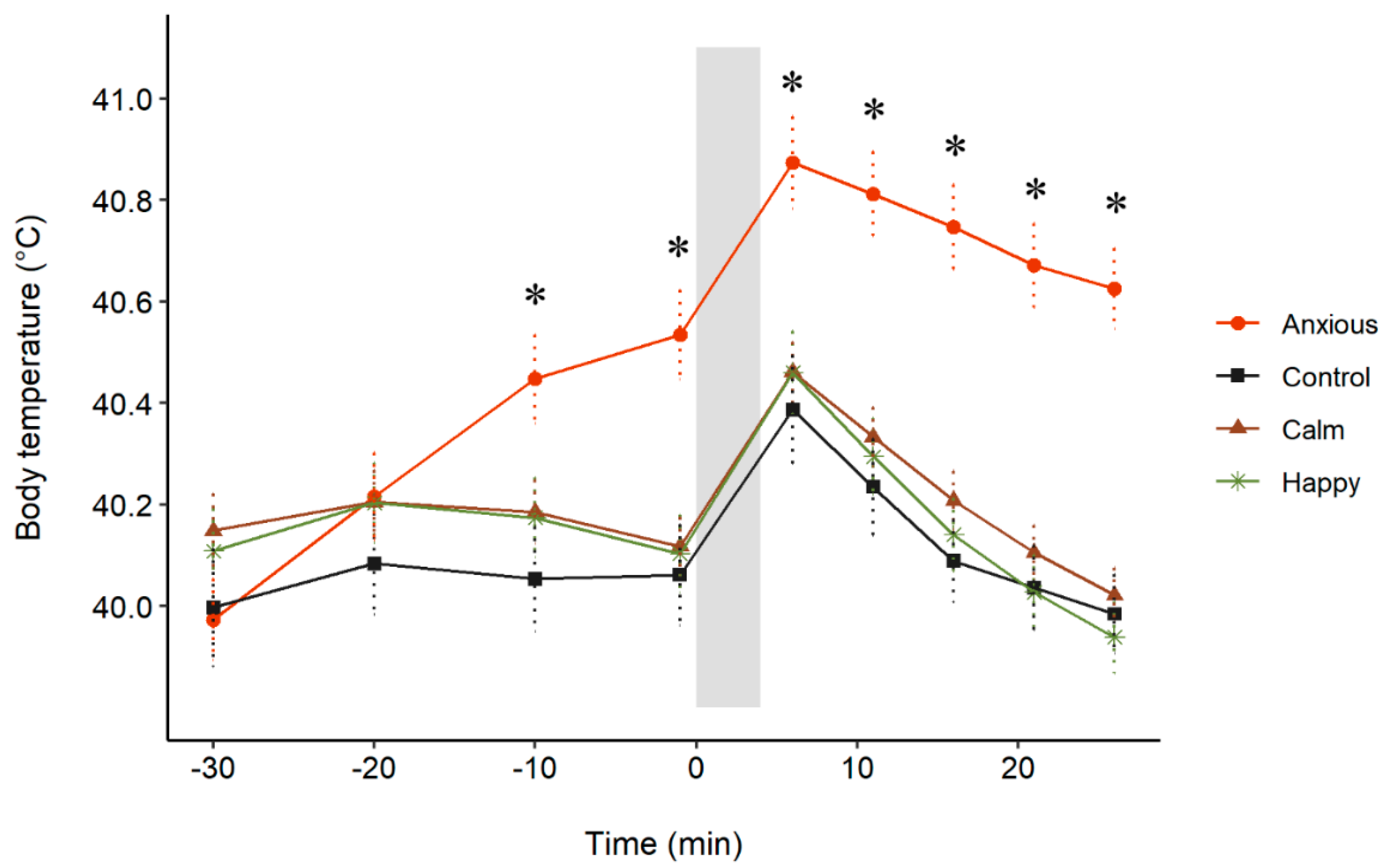
3.3. Body Temperature
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Data Availability
References
- Hemsworth, P.; Mellor, D.; Cronin, G.; Tilbrook, A. Scientific assessment of animal welfare. N. Z. Vet. J. 2015, 63, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Boissy, A.; Manteuffel, G.; Jensen, M.B.; Moe, R.O.; Spruijt, B.; Keeling, L.J.; Winckler, C.; Forkman, B.B.; Dimitrov, I.; Langbein, J.; et al. Assessment of positive emotions in animals to improve their welfare. Physiol. Behav. 2007, 92, 375–397. [Google Scholar] [CrossRef] [PubMed]
- Yeates, J.W.W.; Main, D.C.J. Assessment of positive welfare: A review. Vet. J. 2008, 175, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Mattiello, S.; Battini, M.; De Rosa, G.; Napolitano, F.; Dwyer, C. How Can We Assess Positive Welfare in Ruminants? Animals 2019, 9, 758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paul, E.S.; Harding, E.J.; Mendl, M. Measuring emotional processes in animals: The utility of a cognitive approach. Neurosci. Biobehav. Rev. 2005, 29, 469–491. [Google Scholar] [CrossRef] [PubMed]
- Mendl, M.; Burman, O.H.P.; Paul, E.S. An integrative and functional framework for the study of animal emotion and mood. Proc. R. Soc. B Biol. Sci. 2010, 277, 2895–2904. [Google Scholar] [CrossRef] [Green Version]
- Kremer, L.; Klein Holkenborg, S.E.J.; Reimert, I.; Bolhuis, J.E.; Webb, L.E. The nuts and bolts of animal emotion. Neurosci. Biobehav. Rev. 2020, 113, 273–286. [Google Scholar] [CrossRef]
- Crump, A.; Arnott, G.; Bethell, E. Affect-Driven Attention Biases as Animal Welfare Indicators: Review and Methods. Animals 2018, 8, 136. [Google Scholar] [CrossRef] [Green Version]
- Bar-Haim, Y.; Lamy, D.; Pergamin, L.; Bakermans-Kranenburg, M.J.; van IJzendoorn, M.H. Threat-related attentional bias in anxious and nonanxious individuals: A meta-analytic study. Psychol. Bull. 2007, 133, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Bradley, B.P.; Mogg, K.; Lee, S.C. Attentional biases for negative information in induced and naturally occurring dysphoria. Behav. Res. Ther. 1997, 35, 911–927. [Google Scholar] [CrossRef]
- Bradley, B.P.; Mogg, K.; Millar, N.; White, J. Selective processing of negative information: Effects of clinical anxiety, concurrent depression, and awareness. J. Abnorm. Psychol. 1995, 104, 532–536. [Google Scholar] [CrossRef] [PubMed]
- Cisler, J.M.; Koster, E.H.W. Mechanisms of attentional biases towards threat in anxiety disorders: An integrative review. Clin. Psychol. Rev. 2010, 30, 203–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamir, M.; Robinson, M.D. The Happy Spotlight: Positive Mood and Selective Attention to Rewarding Information. Personal. Soc. Psychol. Bull. 2007, 33, 1124–1136. [Google Scholar] [CrossRef] [Green Version]
- Sanchez, A.; Vazquez, C. Looking at the eyes of happiness: Positive emotions mediate the influence of life satisfaction on attention to happy faces. J. Posit. Psychol. 2014, 9, 435–448. [Google Scholar] [CrossRef]
- Eizenman, M.; Yu, L.H.; Grupp, L.; Eizenman, E.; Ellenbogen, M.; Gemar, M.; Levitan, R.D. A naturalistic visual scanning approach to assess selective attention in major depressive disorder. Psychiatry Res. 2003, 118, 117–128. [Google Scholar] [CrossRef]
- Hermans, D.; Vansteenwegen, D.; Eelen, P. Eye Movement Registration as a Continuous Index of Attention Deployment: Data from a Group of Spider Anxious Students. Cogn. Emot. 1999, 13, 419–434. [Google Scholar] [CrossRef]
- Kellough, J.L.; Beevers, C.G.; Ellis, A.J.; Wells, T.T. Time course of selective attention in clinically depressed young adults: An eye tracking study. Behav. Res. Ther. 2008, 46, 1238–1243. [Google Scholar] [CrossRef] [Green Version]
- Bethell, E.J.; Holmes, A.; MacLarnon, A.; Semple, S. Evidence That Emotion Mediates Social Attention in Rhesus Macaques. PLoS ONE 2012, 7, e44387. [Google Scholar] [CrossRef] [Green Version]
- Winters, S.; Dubuc, C.; Higham, J.P. Perspectives: The Looking Time Experimental Paradigm in Studies of Animal Visual Perception and Cognition. Ethology 2015, 121, 625–640. [Google Scholar] [CrossRef]
- Lee, C.; Verbeek, E.; Doyle, R.; Bateson, M. Attention bias to threat indicates anxiety differences in sheep. Biol. Lett. 2016, 12, 20150977. [Google Scholar] [CrossRef] [Green Version]
- Monk, J.E.; Doyle, R.E.; Colditz, I.G.; Belson, S.; Cronin, G.M.; Lee, C. Towards a more practical attention bias test to assess affective state in sheep. PLoS ONE 2018, 13, e0190404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verbeek, E.; Colditz, I.; Blache, D.; Lee, C. Chronic stress influences attentional and judgement bias and the activity of the HPA axis in sheep. PLoS ONE 2019, 14, e0211363. [Google Scholar] [CrossRef] [PubMed]
- Monk, J.E.; Belson, S.; Lee, C. Pharmacologically-induced stress has minimal impact on judgement and attention biases in sheep. Sci. Rep. 2019, 9, 11446. [Google Scholar] [CrossRef] [PubMed]
- Monk, J.E.; Belson, S.; Colditz, I.G.; Lee, C. Attention Bias Test Differentiates Anxiety and Depression in Sheep. Front. Behav. Neurosci. 2018, 12, 246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynch, J.J.; Hinch, G.N.; Adams, D.B. The Behaviour of Sheep: Biological Principles and Implications for Production; CAB International: Wallingford, UK, 1992; ISBN 0643053298.
- Monk, J.E.; Lee, C.; Belson, S.; Colditz, I.G.; Campbell, D.L.M. The influence of pharmacologically-induced affective states on attention bias in sheep. PeerJ 2019, 7, e7033. [Google Scholar] [CrossRef] [PubMed]
- Bouwknecht, J.A.; Olivier, B.; Paylor, R.E. The stress-induced hyperthermia paradigm as a physiological animal model for anxiety: A review of pharmacological and genetic studies in the mouse. Neurosci. Biobehav. Rev. 2007, 31, 41–59. [Google Scholar] [CrossRef]
- Sherwood, L.; Klandorf, H.; Yancey, P.H. Animal Physiology: From Genes to Organisms; Thomson/Brooks/Cole: Southbank, Australia, 2005; ISBN 0534554040. [Google Scholar]
- Olivier, B.; Zethof, T.; Pattij, T.; van Boogaert, M.; van Oorschot, R.; Leahy, C.; Oosting, R.; Bouwknecht, A.; Veening, J.; van der Gugten, J.; et al. Stress-induced hyperthermia and anxiety: Pharmacological validation. Eur. J. Pharmacol. 2003, 463, 117–132. [Google Scholar] [CrossRef]
- Drake, K.A. The Neurophysiological Regulation of Temperament in Sheep. Ph.D. Thesis, University of New England, Armidale, Australia, 2006. [Google Scholar]
- Doyle, R.E.; Lee, C.; McGill, D.M.; Mendl, M. Evaluating pharmacological models of high and low anxiety in sheep. PeerJ 2015, 3, e1510. [Google Scholar] [CrossRef] [Green Version]
- Destrez, A.; Deiss, V.; Belzung, C.; Lee, C.; Boissy, A. Does reduction of fearfulness tend to reduce pessimistic-like judgment in lambs? Appl. Anim. Behav. Sci. 2012, 139, 233–241. [Google Scholar] [CrossRef]
- Verbeek, E.; Ferguson, D.; Quinquet de Monjour, P.; Lee, C. Generating positive affective states in sheep: The influence of food rewards and opioid administration. Appl. Anim. Behav. Sci. 2014, 154, 39–47. [Google Scholar] [CrossRef]
- Riley, J.L.; Hastie, B.A.; Glover, T.L.; Fillingim, R.B.; Staud, R.; Campbell, C.M. Cognitive—Affective and Somatic Side Effects of Morphine and Pentazocine: Side-Effect Profiles in Healthy Adults. Pain Med. 2010, 11, 195–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lea, J.M.; Niemeyer, D.D.O.; Reed, M.T.; Fisher, A.D.; Ferguson, D.M. Development and validation of a simple technique for logging body temperature in free-ranging cattle. Aust. J. Exp. Agric. 2008, 48, 741. [Google Scholar] [CrossRef]
- R Core Team. R: A language and environment for statistical computing (Software version 3.6.0). R foundation for statistical computing, Vienna, Austria. 2019. Available online: https://www.R-project.org/ (accessed on 29 February 2020).
- Field, A.P.; Miles, J.; Field, Z. Discovering Statistics Using R.; SAGE: London, UK, 2012; ISBN 9781446200469. [Google Scholar]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; R Core Team. Nlme: Linear and Nonlinear Mixed Effects Models. R package version 3.1-139. 2016. Available online: https://CRAN.R-project.org/package=nlme (accessed on 24 May 2020).
- Giraudoux, P. Pgirmess: Data Analysis in Ecology. R package version 1.6.9. 2016. Available online: https://CRAN.R-project.org/package=pgirmess (accessed on 12 March 2020).
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Zeileis, A.; Kleiber, C.; Jackman, S. Regression Models for Count Data in R. J. Stat. Softw. 2008, 27, 1–25. [Google Scholar] [CrossRef] [Green Version]
- Mangiafico, S. Rcompanion: Functions to Support Extension Education Program Evaluation. R package version 2.1.7. 2019. Available online: https://CRAN.R-project.org/package=rcompanion (accessed on 9 February 2020).
- Therneau, T.M.; Grambsch, P.M. Modeling Survival Data: Extending the Cox Model; Statistics for Biology and Health; Springer: New York, NY, USA, 2000; ISBN 978-1-4419-3161-0. [Google Scholar]
- Therneau, T. A Package for Survival Analysis in R. R package version 2.38. 2015. Available online: https://CRAN.R-project.org/package=survival (accessed on 13 June 2020).
- Hinch, G.N. Understanding the natural behaviour of sheep. In Advances in Sheep Welfare; Woodhead Publishing: Duxford, UK, 2017; pp. 3–18. ISBN 9780081007181. [Google Scholar]
- Dwyer, C.M. How has the risk predation shaped the behavioural responses of sheep to fear and distress? Anim. Welf. 2004, 13, 269–281. [Google Scholar]
- Hulet, C.V. A review: Understanding sheep behavior, a key to more efficient and profitable lamb and wool production. Sheep Res. J. 1989, 5, 26–33. [Google Scholar]
- Masterson, F.A.; Crawford, M. The defense motivation system: A theory of avoidance behavior. Behav. Brain Sci. 1982, 5, 661–675. [Google Scholar] [CrossRef]
- Vanlessen, N.; De Raedt, R.; Koster, E.H.W.; Pourtois, G. Happy heart, smiling eyes: A systematic review of positive mood effects on broadening of visuospatial attention. Neurosci. Biobehav. Rev. 2016, 68, 816–837. [Google Scholar] [CrossRef]
- Beausoleil, N.J.; Stafford, K.J.; Mellor, D.J. Sheep show more aversion to a dog than to a human in an arena test. Appl. Anim. Behav. Sci. 2005, 91, 219–232. [Google Scholar] [CrossRef]
- Torres-Hernandez, G.; Hohenboken, W. An attempt to assess traits of emotionality in crossbred ewes. Appl. Anim. Ethol. 1979, 5, 71–83. [Google Scholar] [CrossRef]
- Piggins, D.; Phillips, C.J.C. The eye of the domesticated sheep with implications for vision. Anim. Sci. 1996, 62, 301–308. [Google Scholar] [CrossRef]
- Kendrick, K.M. Sheep Senses, Social Cognition and Capacity for Consciousness. In The Welfare of Sheep; Springer: Dordrecht, The Netherlands, 2008; pp. 135–157. ISBN 978-1-4020-8553-6. [Google Scholar]
- Raoult, C.; Gygax, L. Valence and Intensity of Video Stimuli of Dogs and Conspecifics in Sheep: Approach-Avoidance, Operant Response, and Attention. Animals 2018, 8, 121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vögeli, S.; Wolf, M.; Wechsler, B.; Gygax, L. Housing conditions influence cortical and behavioural reactions of sheep in response to videos showing social interactions of different valence. Behav. Brain Res. 2015, 284, 69–76. [Google Scholar] [CrossRef] [Green Version]
- Bellegarde, L.G.A.; Haskell, M.J.; Duvaux-Ponter, C.; Weiss, A.; Boissy, A.; Erhard, H.W. Face-based perception of emotions in dairy goats. Appl. Anim. Behav. Sci. 2017, 193, 51–59. [Google Scholar] [CrossRef] [Green Version]
- Vögeli, S.; Wechsler, B.; Gygax, L. Welfare by the ear: Comparing relative durations and frequencies of ear postures by using an automated tracking system in sheep. Anim. Welf. 2014, 23, 267–274. [Google Scholar] [CrossRef]
- Raoult, C.M.C.; Gygax, L. Mood induction alters attention toward negative-positive stimulus pairs in sheep. Sci. Rep. 2019, 9, 7759. [Google Scholar] [CrossRef]
- Gaskins, L.A.; Massey, J.G.; Ziccardi, M.H. Effect of oral diazepam on feeding behavior and activity of Hawai’i ’amakihi (Hemignathus virens). Appl. Anim. Behav. Sci. 2008, 112, 384–394. [Google Scholar] [CrossRef]
- Foltin, R.W. Effects of amphetamine, dexfenfluramine, diazepam, and dietary manipulations on responding reinforced by stimuli paired with food in nonhuman primates. Pharmacol. Biochem. Behav. 2004, 77, 471–479. [Google Scholar] [CrossRef]
- Brioni, J.D.; Arolfo, M.P. Diazepam impairs retention of spatial information without affecting retrieval or cue learning. Pharmacol. Biochem. Behav. 1992, 41, 1–5. [Google Scholar] [CrossRef]
- Sasaki-Hamada, S.; Sacai, H.; Sugiyama, A.; Iijima, T.; Saitoh, A.; Inagaki, M.; Yamada, M.; Oka, J.-I. Riluzole Does Not Affect Hippocampal Synaptic Plasticity and Spatial Memory, Which Are Impaired by Diazepam in Rats. J. Pharmacol. Sci. 2013, 122, 232–236. [Google Scholar] [CrossRef] [Green Version]
- Verbeek, E.; Ferguson, D.; Quinquet de Monjour, P.; Lee, C. Opioid control of behaviour in sheep: Effects of morphine and naloxone on food intake, activity and the affective state. Appl. Anim. Behav. Sci. 2012, 142, 18–29. [Google Scholar] [CrossRef]
- Forkman, B.; Boissy, A.; Meunier-Salaün, M.C.; Canali, E.; Jones, R.B. A critical review of fear tests used on cattle, pigs, sheep, poultry and horses. Physiol. Behav. 2007, 92, 340–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dodd, C.L.; Pitchford, W.S.; Hocking Edwards, J.E.; Hazel, S.J. Measures of behavioural reactivity and their relationships with production traits in sheep: A review. Appl. Anim. Behav. Sci. 2012, 140, 1–15. [Google Scholar] [CrossRef]
- Wright, F.L.; Rodgers, R.J. On the behavioural specificity of hypophagia induced in male rats by mCPP, naltrexone, and their combination. Psychopharmacology 2014, 231, 787–800. [Google Scholar] [CrossRef] [PubMed]
- Krebs, H.; Macht, M.; Weyers, P.; Weijers, H.-G.; Janke, W. Effects of Stressful Noise on Eating and Non-eating Behavior in Rats. Appetite 1996, 26, 193–202. [Google Scholar] [CrossRef]
- Martí, O.; Martí, J.; Armario, A. Effects of chronic stress on food intake in rats: Influence of stressor intensity and duration of daily exposure. Physiol. Behav. 1994, 55, 747–753. [Google Scholar] [CrossRef]
- Feuchtl, A.; Bagil, M.; Stephan, R.; Frahnert, C.; Kölsch, H.; Hühn, K.-U.; Rao, M.L.; Bagli, M.; Stephan, R.; Frahnert, C.; et al. Pharmacokinetics of m-Chlorophenylpiperazine after intrevenous and oral administration in healthy male volunteers: Implication for the pharmacodynamic profile. Pharmacopsychiatry 2004, 37, 180–188. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Behavior | Definition |
|---|---|
| Attention to dog | Duration spent looking at the dog window wall with binocular vision (s), where direction of binocular vision was assessed using head orientation, observed from video footage taken at a top-down angle (Video S1) |
| Attention to photo | Duration spent looking at the conspecific photo wall with binocular vision (s) |
| Vigilance | Duration spent with the head at or above shoulder height (s) and latency to become non-vigilant (s) |
| Sniff photo | Frequency of sniffing (n) and latency to sniff the conspecific photo (s) |
| Sniff environment | Frequency of sniffing (n) and latency to sniff the floor or walls of the arena (s) |
| Zones crossed | Number of zones crossed in the arena with two front feet in a new zone or one front foot in the new zone and the other on the line (n) |
| Zones entered | Number of available zones entered (n; ranging from one to nine zones available within the arena, each new zone entered was only counted once during the test) |
| Zone durations | Duration spent standing in the zone closest to the conspecific photo (s) and the number of animals that entered the zone closest to the dog window (n; analysed instead of duration due to low number of entries into zone) |
| Vocalizations | Number of open-and close-mouthed bleats during testing (n) |
| Urinations | Number of urinations (n) |
| Behavior | Anxious | Control | Calm | Happy | Statistical Test | Test Value (df) | p-Value |
|---|---|---|---|---|---|---|---|
| Vigilance duration (s) 1,2 | 5.12 ± 0.01 (177) a | 5.13 ± 0.01 (169) bc | 5.11 ± 0.01 (166) c | 5.16 ± 0.01 (174) ab | LME | F(3,75) = 6.0 | 0.002 |
| Vigilance (mean rank) | 56.5 ± 4.5 (177) a | 30.9 ± 4.4 (170) b | 27.9 ± 4.5 (167) b | 46.8 ± 4.8 (175) ab | Kruskal-Wallis | X2(3) = 20 | <0.001 |
| Looking towards dog (s) | 39.8 ± 3.7 | 40.9 ± 3.7 | 39.5 ± 3.7 | 39.3 ± 3.7 | LME | F(3,75) = 0.1 | 0.967 |
| Looking towards photo (s) 1 | 3.9 ± 0.1 (47.5) | 3.7 ± 0.1 (39.5) | 3.9 ± 0.1 (48.9) | 3.7 ± 0.1 (40.7) | LME | F(3,75) = 2.0 | 0.141 |
| Sniff photo (n) 1 | 0.4 ± 0.2 (1.5) a | 1.3 ± 0.1 (3.5) b | 1.4 ± 0.1 (4.2) b | 1.0 ± 0.1 (2.8) b | GLME_P | X2(3) = 25.9 | <0.001 |
| Sniff environment (n) 1 | −0.4 ± 0.3 (0.7) a | 1.4 ± 0.2 (4.0) b | 1.3 ± 0.2 (3.8) b | 1.1 ± 0.2 (3.1) b | GLME_NB | X2(3) = 26.7 | <0.001 |
| Sniff closed window (n) 3 | 0 | 1 | 2 | 1 | FET | n/A | 0.900 |
| Standing near photo (s) | 72.0 ± 8.0 | 79.2 ± 8.0 | 59.1 ± 8.0 | 66.5 ± 8.0 | LME | F(3,75) = 1.2 | 0.336 |
| Zones crossed (n) 1 | 2.81 ± 0.1 (16.6) a | 2.98 ± 0.1 (19.8) ab | 3.35 ± 0.1 (28.4) b | 3.25 ± 0.1 (25.9) ab | GLME_NB | X2(3) = 10.6 | 0.014 |
| Zones entered (n) 1 | 1.73 ± 0.1 (5.65) | 1.82 ± 0.1 (6.15) | 1.97 ± 0.1 (7.20) | 1.82 ± 0.1 (6.20) | GLME_P | X2(3) = 4 | 0.260 |
| Enter zone close to dog (n) 3 | 5 | 7 | 13 | 9 | FET | n/A | 0.070 |
| Open-mouthed bleats (n) | 9.0 ± 2.3 | 11.1 ± 2.0 | 12.2 ± 2.3 | 25.9 ± 3.6 | GLME_NB_H | X2(3) = 5.5 | 0.140 |
| Close-mouthed bleats (n) | 6.7 ± 0.5 | 9.9 ± 0.7 | 9.5 ± 0.7 | 9.6 ± 0.7 | GLME_NB_H | X2(3) = 1.9 | 0.580 |
| Urinations (n) 3 | 7 a | 1 ab | 2 ab | 0 b | FET | n/A | 0.008 |
| Head shaking (n) 3 | 5 | 1 | 1 | 2 | FET | n/A | 0.09 |
| Tail shaking (n) 3 | 9 a | 0 b | 0 b | 1 a | FET | n/A | <0.001 |
| Latency to | Group | Mean (s) 1 | Censored (n) 2 | Coefficient 3 | SE (Coeff) | Hazard Ratio (95% CI) | Wald (z) | p-Value | Likelihood Ratio | df | p-Value |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Non-vigilance | Control | 42.7 a | 1 | Reference | 18.2 | 3 | <0.001 | ||||
| Anxious | 119.9 b | 9 | −1.533 | 0.392 | 0.22 (0.10–0.47) | −3.91 | <0.001 | ||||
| Calm | 56.2 ac | 0 | −0.345 | 0.324 | 0.71 (0.38–1.34) | −1.06 | 0.288 | ||||
| Happy | 75.3 c | 4 | −0.716 | 0.343 | 0.49 (0.25–0.96) | −2.08 | 0.04 | ||||
| Anxious | Reference | ||||||||||
| Calm | 1.188 | 0.390 | 3.28 (1.53–7.04) | 3.05 | 0.002 | ||||||
| Happy | 0.817 | 0.395 | 2.26 (1.04–4.91) | 2.07 | 0.039 | ||||||
| Calm | Reference | ||||||||||
| Happy | −0.371 | 0.34 | 0.69 (0.35–1.35) | −1.09 | 0.277 | ||||||
| Sniff photo | Control | 41.5 | 3 | Reference | 6.7 | 3 | 0.08 | ||||
| Anxious | 77.9 | 7 | −0.713 | 0.37 | 0.49 (0.23–1.01) | −1.93 | 0.053 | ||||
| Calm | 26.4 | 0 | 0.144 | 0.33 | 1.16 (0.60–2.20) | 0.43 | 0.665 | ||||
| Happy | 46.8 | 3 | −0.228 | 0.33 | 0.80 (0.41–1.56) | −0.66 | 0.507 | ||||
| Anxious | Reference | ||||||||||
| Calm | 0.850 | 0.36 | 2.36 (1.16–4.79) | 2.37 | 0.018 | ||||||
| Happy | 0.490 | 0.37 | 1.63 (0.79–3.36) | 1.31 | 0.190 | ||||||
| Calm | Reference | ||||||||||
| Happy | −0.372 | 0.33 | 0.69 (0.36–1.32) | −1.12 | 0.262 | ||||||
| Sniff environment | Control | 70.5 a | 2 | Reference | 25.8 | 3 | <0.001 | ||||
| Anxious | 143.7 b | 14 | −1.905 | 0.48 | 0.15 (0.06–0.38) | –3.97 | <0.001 | ||||
| Calm | 79.1 a | 1 | −0.073 | 0.33 | 0.93 (0.49–1.77) | –0.22 | 0.820 | ||||
| Happy | 86.1 a | 2 | −0.262 | 0.34 | 0.77 (0.40–1.48) | –0.78 | 0.430 | ||||
| Anxious | Reference | ||||||||||
| Calm | 1.832 | 0.48 | 6.25 (2.45–15.9) | 3.83 | <0.001 | ||||||
| Happy | 1.643 | 0.48 | 5.17 (2.03–13.2) | 3.44 | <0.001 | ||||||
| Calm | Reference | ||||||||||
| Happy | −0.189 | 0.33 | 0.83 (0.43–1.59) | −0.57 | 0.568 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Monk, J.E.; Lee, C.; Dickson, E.; Campbell, D.L.M. Attention Bias Test Measures Negative But Not Positive Affect in Sheep: A Replication Study. Animals 2020, 10, 1314. https://doi.org/10.3390/ani10081314
Monk JE, Lee C, Dickson E, Campbell DLM. Attention Bias Test Measures Negative But Not Positive Affect in Sheep: A Replication Study. Animals. 2020; 10(8):1314. https://doi.org/10.3390/ani10081314
Chicago/Turabian StyleMonk, Jessica E., Caroline Lee, Emily Dickson, and Dana L. M. Campbell. 2020. "Attention Bias Test Measures Negative But Not Positive Affect in Sheep: A Replication Study" Animals 10, no. 8: 1314. https://doi.org/10.3390/ani10081314