Antimicrobial Effects of Black Soldier Fly and Yellow Mealworm Fats and Their Impact on Gut Microbiota of Growing Rabbits

 , ,
, ,  ,
,  , , , , , , and
, , , , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Ethics Statement
2.2. In Vitro Analyses for Antimicrobial Activity of Insect Fats
2.3. Inclusion of Hermetia illucens and Tenebrio molitor Fats in the Diet of Growing Rabbits
2.3.1. Experimental Design
2.3.2. Fatty Acid Profiles of Insect Lipids and Experimental Diets
2.3.3. Cecal Fermentation Traits
2.3.4. DNA Extraction and 16S rRNA High-Throughput Amplicon Target Sequencing
2.3.5. Bioinformatics and Statistical Analysis
3. Results
3.1. In Vitro Antimicrobial Activities of Insect Fats
3.2. Cecal Fermentation Traits
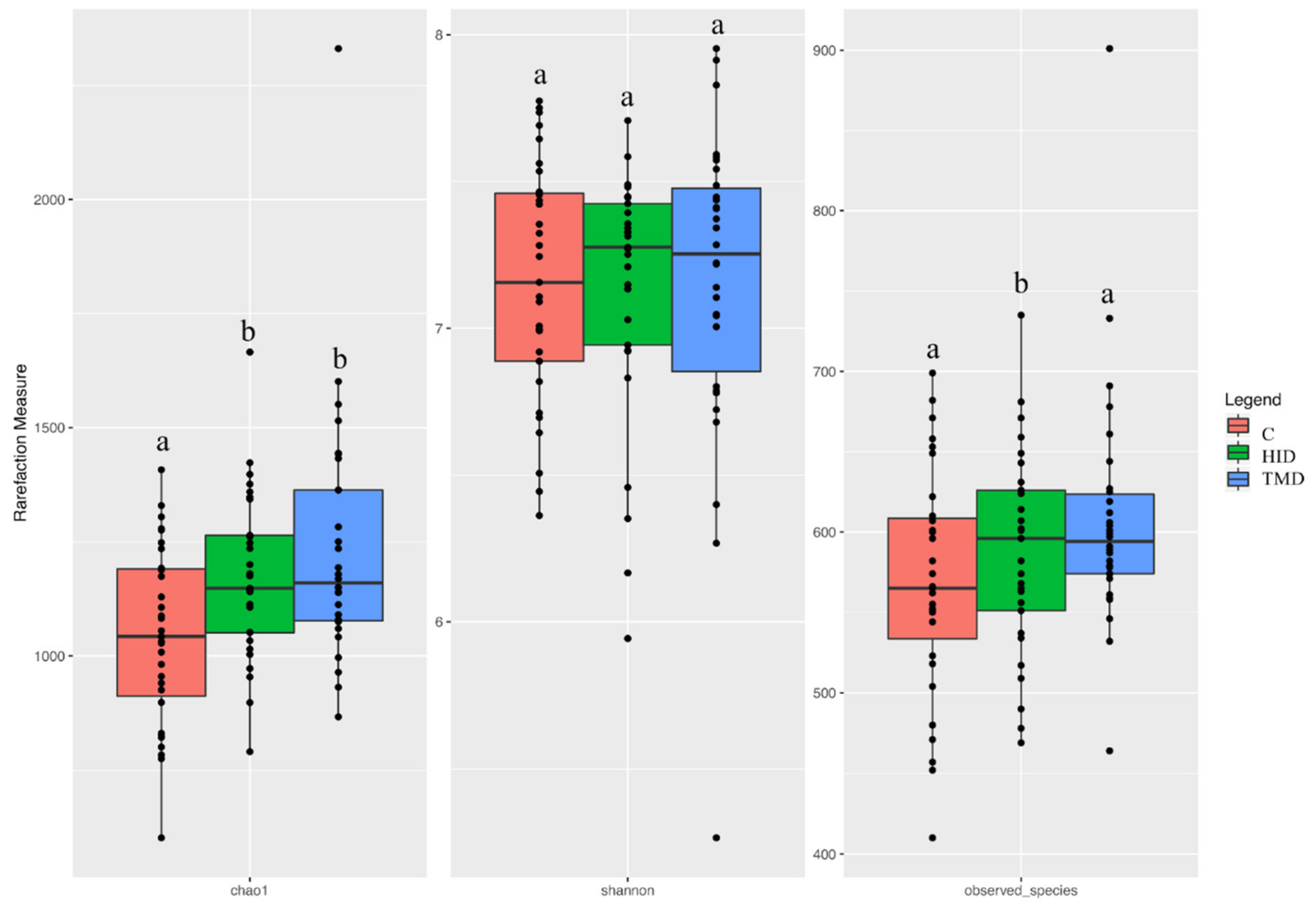
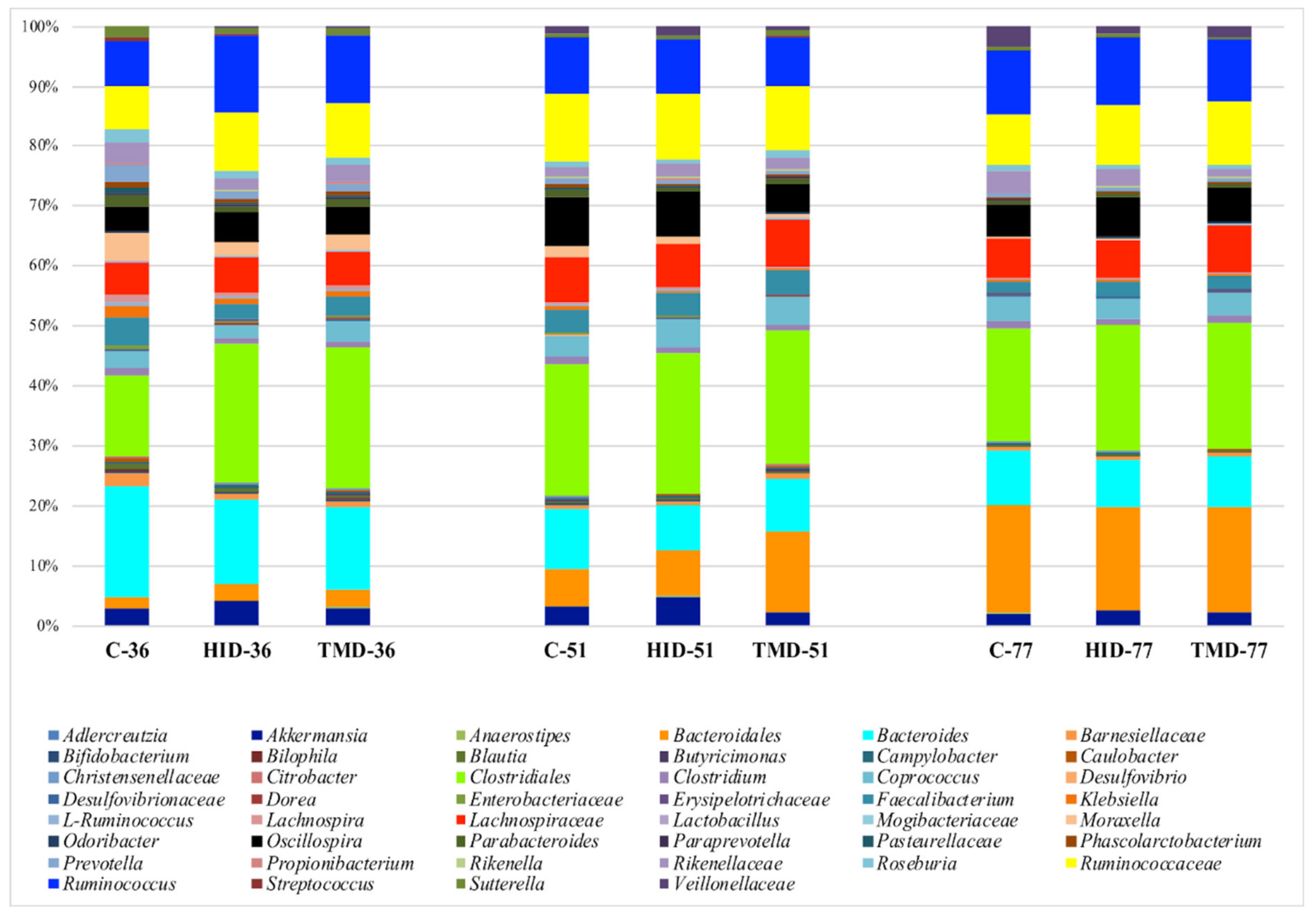
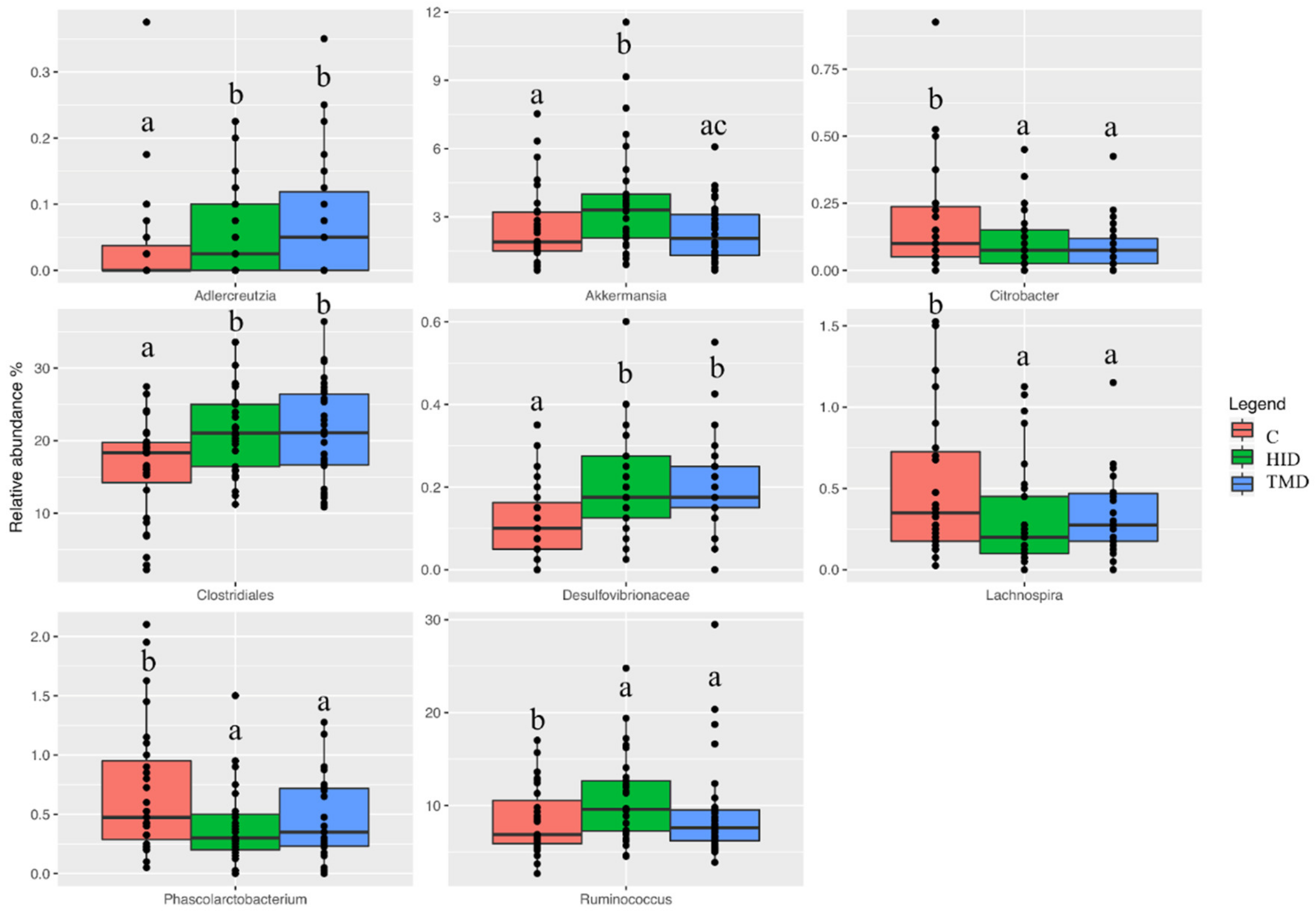
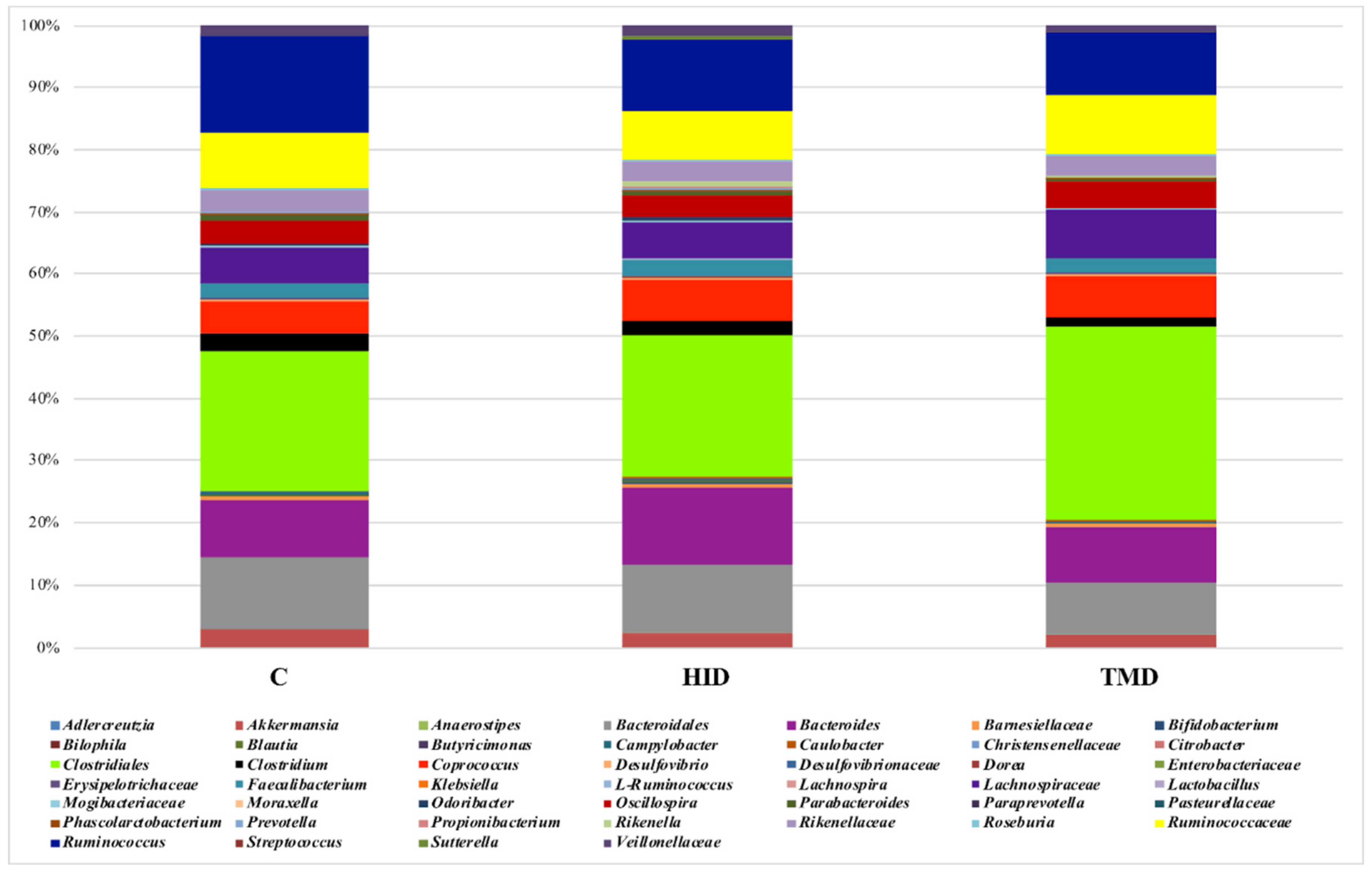
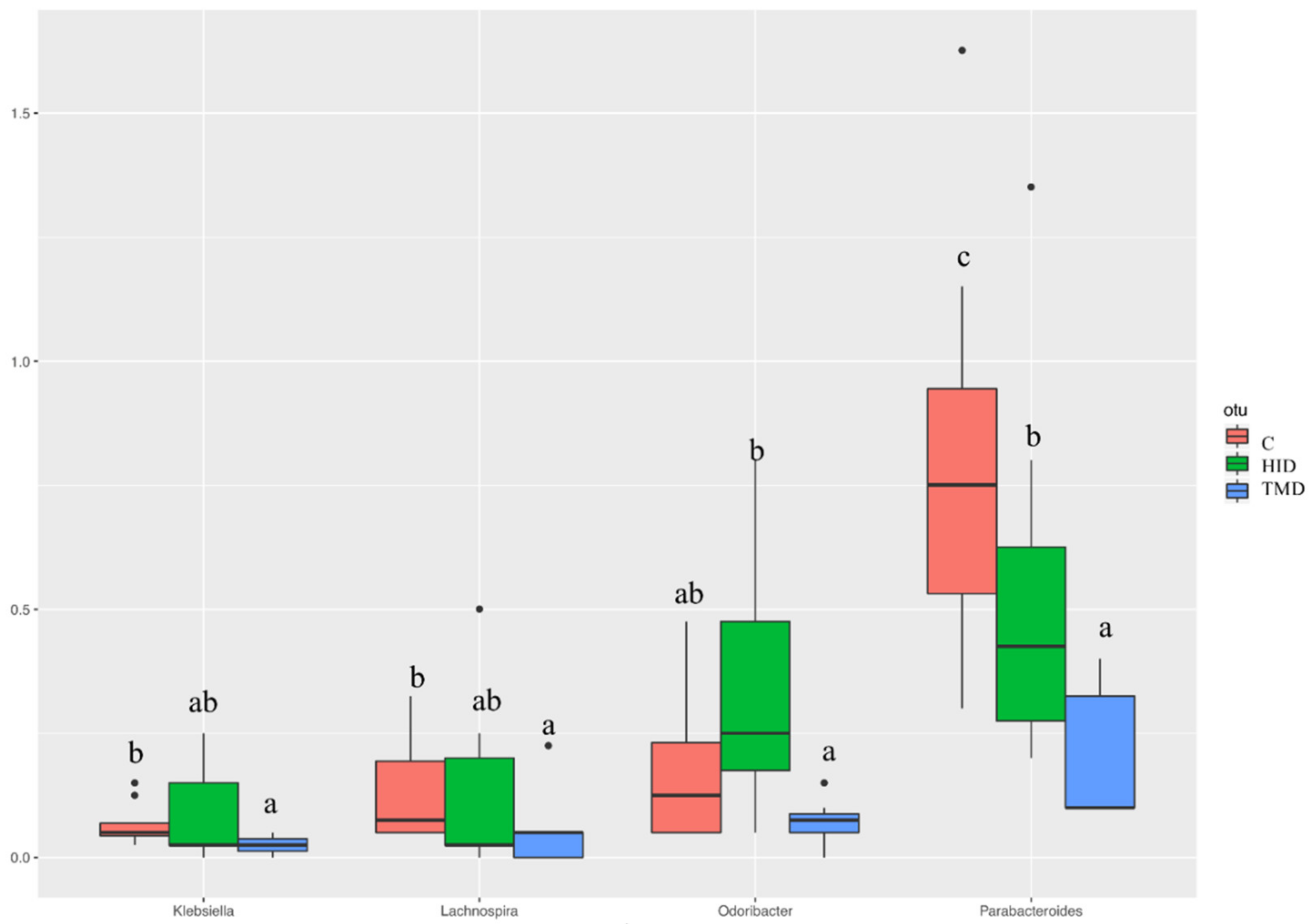
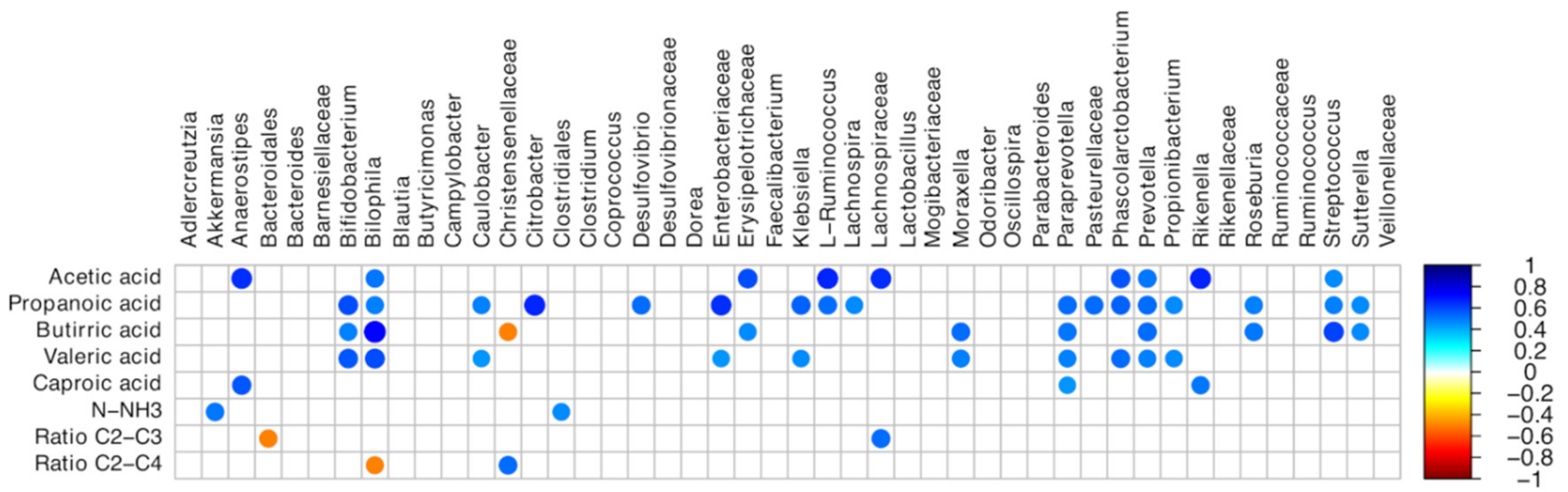
3.3. Cecal and Fecal Microbiota Characterization
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Licois, D.; Wyers, M.; Coudert, P. Epizootic Rabbit Enteropathy: Experimental transmission and clinical characterization. Vet. Res. 2005, 36, 601–613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Combes, S.; Fortun-Lamothe, L.; Cauquil, L.; Gidenne, T. Engineering the rabbit digestive ecosystem to improve digestive health and efficacy. Animal 2013, 7, 1429–1439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vántus, V.B.; Kovács, M.; Zsolnai, A. The rabbit cecal microbiota: Development, composition and its role in the prevention of digestive diseases—A review on recent literature in the light of molecular genetic methods. Acta Agrar. Kaposváriensis 2014, 18, 55–65. [Google Scholar]
- Fortun-Lamothe, L.; Theau-Clément, M.; Combes, S.; Allain, D.; Lebas, F.; Le Normand, B.; Gidenne, T. Physiologie. In Le Lapin de la Biologie à L’élevage, 1st ed.; Gidenne, T., Ed.; Quae: Versailles, France, 2015; pp. 48–49. ISBN 9782759224166. [Google Scholar]
- Fortun-Lamothe, L.; Boullier, S. A review on the interactions between gut microflora and digestive mucosal immunity. Possible ways to improve the health of rabbits. Livest. Sci. 2007, 107, 1–18. [Google Scholar] [CrossRef]
- Kim, S.J.; Kim, S.E.; Kim, A.R.; Kang, S.; Park, M.Y.; Sung, M.K. Dietary fat intake and age modulate the composition of the gut microbiota and colonic inflammation in C57BL/6J mice. BMC Microbiol. 2019, 19, 193. [Google Scholar] [CrossRef]
- Yang, Q.; Liang, Q.; Balakrishnan, B.; Belobrajdic, D.P.; Feng, Q.J.; Zhang, W. Role of dietary nutrients in the modulation of gut microbiota: A narrative review. Nutrients 2020, 12, 381. [Google Scholar] [CrossRef] [Green Version]
- Yoon, B.K.; Jackman, J.A.; Valle-González, E.R.; Cho, N.J. Antibacterial free fatty acids and monoglycerides: Biological activities, experimental testing, and therapeutic applications. Int. J. Mol. Sci. 2018, 19, 1114. [Google Scholar] [CrossRef] [Green Version]
- Xiccato, G. Fat Digestion. In Nutrition of the Rabbit, 3rd ed.; De Blas, C., Wiseman, J., Eds.; CAB International: Wallingford, UK, 2020; pp. 58–68. ISBN 9781789241273. [Google Scholar] [CrossRef]
- Gidenne, T.; Lebas, F.; Licois, D.; García, J. Nutrition and Feeding Strategy: Impact on Health Status. In Nutrition of the Rabbit, 3rd ed.; De Blas, C., Wiseman, J., Eds.; CAB International: Wallingford, UK, 2020; pp. 193–221. ISBN 9781789241273. [Google Scholar] [CrossRef]
- Christaki, E.; Marcou, M.; Tofarides, A. Antimicrobial resistance in Bacteria: Mechanisms, evolution, and persistence: Review. J. Mol. Evol. 2020, 88, 26–40. [Google Scholar] [CrossRef]
- Girard, M.; Bee, G. Invited review: Tannins as a potential alternative to antibiotics to prevent coliform diarrhea in weaned pigs. Animal 2020, 14, 95–107. [Google Scholar] [CrossRef] [Green Version]
- Gasco, L.; Biasato, I.; Dabbou, S.; Schiavone, A.; Gai, F. Animals fed insect-based diets: State-of-the-art on digestibility, performance and product quality. Animals 2019, 9, 170. [Google Scholar] [CrossRef] [Green Version]
- Gasco, L.; Dabbou, S.; Trocino, A.; Xiccato, G.; Capucchio, M.T.; Biasato, I.; Dezzutto, D.; Birolo, M.; Meneguz, M.; Schiavone, A.; et al. Effect of dietary supplementation with insect fats on growth performance, digestive efficiency and health of rabbits. J. Anim. Sci. Biotechnol. 2019, 10, 4. [Google Scholar] [CrossRef] [PubMed]
- Sypniewski, J.; Kierończyk, B.; Benzertiha, A.; Mikołajczak, Z.; Pruszyńska Oszmałek, E.; Kołodziejski, P.; Sassek, M.; Rawski, M.; Czekała, W.; Józefiak, D. Replacement of soybean oil by Hermetia illucens fat in turkey nutrition: Effect on performance, digestibility, microbial community, immune and physiological status and final product quality. Br. Poult. Sci. 2020, 61, 294–302. [Google Scholar] [CrossRef]
- Borrelli, L.; Coretti, L.; Dipineto, L.; Bovera, F.; Menna, F.; Chiariotti, L.; Nizza, A.; Lembo, F.; Fioretti, A. Insect based diet, a promising nutritional source, modulates gut microbiota composition and SCFAs production in laying hens. Sci. Rep. 2017, 7, 16269. [Google Scholar] [CrossRef] [Green Version]
- Biasato, I.; Ferrocino, I.; Biasibetti, E.; Grego, E.; Dabbou, S.; Sereno, A.; Gai, F.; Gasco, L.; Schiavone, A.; Cocolin, L.; et al. Modulation of intestinal microbiota, morphology and mucin composition by dietary insect meal inclusion in free-range chickens. BMC Vet. Res. 2018, 14, 383. [Google Scholar] [CrossRef] [Green Version]
- Biasato, I.; Ferrocino, I.; Grego, E.; Dabbou, S.; Gai, F.; Gasco, L.; Cocolin, L.; Capucchio, M.T.; Schiavone, A. Gut microbiota and mucin composition in female broiler chickens fed diets including yellow mealworm (Tenebrio molitor, L.). Animals, 2019; 9, 213. [Google Scholar] [CrossRef] [Green Version]
- Biasato, I.; Ferrocino, I.; Dabbou, S.; Evangelista, R.; Gai, F.; Gasco, L. Black soldier fly and gut health in broiler chickens: Insights into the relationship between caecal microbiota and intestinal mucin composition. J. Anim. Sci. Biotechnol. 2020, 11, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berezina, N. Insects: Novel source of lipids for a fan of applications. Oil Seeds Fats Crop. Lipids. 2017, 24, D402. [Google Scholar] [CrossRef]
- Schiavone, A.; Cullere, M.; De Marco, M.; Meneguz, M.; Biasato, I.; Bergagna, S.; Dezzutto, D.; Gai, F.; Dabbou, S.; Gasco, L.; et al. Partial or total replacement of soybean oil by black soldier larvae (Hermetia illucens L.) fat in broiler diets: Effect on growth performances, feed-choice, blood traits, carcass characteristics and meat quality. Ital. J. Anim. Sci. 2017, 16, 93–100. [Google Scholar] [CrossRef] [Green Version]
- Schiavone, A.; Dabbou, S.; De Marco, M.; Cullere, M.; Biasato, I.; Biasibetti, E.; Capucchio, M.T.; Bergagna, S.; Dezzutto, D.; Meneguz, M.; et al. Black soldier fly (Hermetia illucens L.) larva fat inclusion in finisher broiler chicken diet as an alternative fat source. Animal 2018, 12, 2032–2039. [Google Scholar] [CrossRef]
- Dalle Zotte, A.; Cullere, M.; Martins, C.; Alves, S.P.; Freire, J.P.B.; Falcão-e-Cunha, L.; Bessa, R.J.B. Incorporation of Black Soldier Fly (Hermetia illucens L.) larvae fat or extruded linseed in diets of growing rabbits and their effects on meat quality traits including detailed fatty acid composition. Meat Sci. 2018, 146, 50–56. [Google Scholar] [CrossRef]
- Kierónczyk, B.; Rawski, M.; Józefiak, A.; Mazurkiewicz, J.; Świątkiewicz, S.; Siwek, M.; Bednarczyk, M.; Szumacher-Strabel, M.; Cieślak, A.; Benzertiha, A.; et al. Effects of replacing soybean oil with selected insect fats on broilers. Anim. Feed Sci. Technol. 2018, 240, 170–183. [Google Scholar] [CrossRef]
- Benzertiha, A.; Kierónczyk, B.; Rawski, M.; Kołodziejski, P.; Bryszak, M.; Józefiak, D. Insect oil as an alternative to palm oil and poultry fat in broiler chicken nutrition. Animals 2019, 9, 116. [Google Scholar] [CrossRef] [Green Version]
- Gasco, L.; Dabbou, S.; Gai, F.; Brugiapaglia, A.; Schiavone, A.; Birolo, M.; Xiccato, G.; Trocino, A. Quality and consumer acceptance of meat from rabbits fed diets in which soybean oil is replaced with black soldier fly and yellow mealworm fats. Animals 2019, 9, 629. [Google Scholar] [CrossRef] [Green Version]
- Matsue, M.; Mori, Y.; Nagase, S.; Sugiyama, Y.; Hirano, R.; Ogai, K.; Ogura, K.; Kurihara, S.; Okamoto, S. Measuring the antimicrobial activity of lauric acid against various bacteria in human gut microbiota using a new method. Cell Transplant. 2019, 28, 1528–1541. [Google Scholar] [CrossRef]
- Skřivanová, E.; Marounek, M.; Benda, V.; Brezina, P. Susceptibility of Escherichia coli, Salmonella s.p. and Clostridium perfringens to organic acids and monolaurin. Vet. Med. 2006, 51, 81–88. [Google Scholar] [CrossRef] [Green Version]
- Zeitz, J.O.; Fennhoff, J.; Kluge, H.; Stangl, G.I.; Eder, K. Effects of dietary fats rich in lauric and myristic acid on performance, intestinal morphology, gut microbes, and meat quality in broilers. Poult. Sci. 2015, 94, 2404–2413. [Google Scholar] [CrossRef] [PubMed]
- Spranghers, T.; Michiels, J.; Vrancx, J.; Ovyn, A.; Eeckhout, M.; De Clercq, P.; De Smet, S. Gut antimicrobial effects and nutritional value of black soldier fly (Hermetia illucens L.) prepupae for weaned piglets. Anim. Feed Sci. Tech. 2018, 235, 33–42. [Google Scholar] [CrossRef]
- Baltić, B.; Starčević, M.; Đorđević, J.; Mrdović, B.; Marković, R. Importance of medium chain fatty acids in animal nutrition. IOP Conf. Ser. Earth Environ. Sci. 2017, 85, 012048. [Google Scholar] [CrossRef] [Green Version]
- Skřivanová, V.; Marounek, M. A note on the effect of triacylglycerols of caprylic and capric fatty acid on performance, mortality, and digestibility of nutrients in young rabbits. Anim. Feed Sci. Tech. 2006, 127, 161–168. [Google Scholar] [CrossRef]
- Boyen, F.; Haesebrouck, F.; Vanparys, A.; Volf, J.; Mahu, M.; Van Immerseel, F.; Rychlik, I.; Dewulf, J.; Ducatelle, R.; Pasmans, F. Coated fatty acids alter virulence properties of Salmonella Typhimurium and decrease intestinal colonization of pigs. Vet. Microbiol. 2008, 132, 319–327. [Google Scholar] [CrossRef]
- Rodriguez-Calleja, J.M.; Garcia-Lopez, I.; Garcia Lopez, M.L.; Santos, J.A.; Otero, A. Rabbit meats as a source of bacterial foodborne pathogens. J. Food Prot. 2006, 69, 1106–1112. [Google Scholar] [CrossRef]
- De Cesare, A.; Parisi, A.; Mioni, R.; Comin, D.; Lucchi, A.; Manfreda, G. Listeria monocytogenes circulating in rabbit meat products and slaughterhouses in Italy: Prevalence data and comparison among typing results. Foodborne Pathog. Dis. 2017, 14, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Kylie, K.; McEwen, S.A.; Boerlin, P.; Reid-Smith, M.J.; Weese, J.S.; Turner, P.V. Prevalence of antimicrobial resistance in fecal Escherichia coli and Salmonella enterica in Canadian commercial meat, companion, laboratory, and shelter rabbits (Oryctolagus cuniculus) and its association with routine antimicrobial use in commercial meat rabbits. Prev. Vet. Med. 2017, 147, 53–57. [Google Scholar] [PubMed]
- Massacci, F.R.; Magistrali, C.F.; Cucco, L.; Curcio, L.; Bano, L.; Mangili, P.M.; Scoccia, E.; Bisgaard, M.; Aalbæk, B.; Christensen, H. Characterization of Pasteurella multocida involved in rabbit infections. Vet. Microbiol. 2018, 213, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Tazzoli, M.; Trocino, A.; Birolo, M.; Radaelli, G.; Xiccato, G. Optimizing feed efficiency and nitrogen excretion in growing rabbits by increasing dietary energy with high-starch, high-soluble fibre, low-insoluble fibre supply at low protein levels. Livest. Sci. 2015, 172, 59–68. [Google Scholar] [CrossRef]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Schmieder, R.; Edwards, R. Quality control and preprocessing of metagenomic datasets. Bioinformatics 2011, 27, 863–864. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [Green Version]
- Dixon, A.L.; Liang, L.; Moffatt, M.F.; Chen, W.; Heath, S.; Wong, K.C.C.; Taylor, J.; Burnett, E.; Gut, I.; Farrall, M.; et al. A genome-wide association study of global gene expression. Nat. Genet. 2007, 39, 1202–1207. [Google Scholar] [CrossRef]
- Mustafa, N.E.M.; Mariod, A.A.; Matthäus, B. Antibacterial activity of Aspongopus Viduatus (melon bug) oil. J. Food Saf. 2008, 28, 577–586. [Google Scholar] [CrossRef]
- Kabara, J.J.; Swieczkowski, D.M.; Conley, D.J.; Truant, J.P. Fatty Acids and Derivatives as Antimicrobial Agents. Antimicrob. Agents Chemother. 1972, 2, 23–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desbois, A.P. Potential applications of antimicrobial fatty acids in medicine, agriculture and other industries. Recent Pat. Antiinfect. Drug Discov. 2012, 7, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Davies, J.; Davies, D. Origins and evolution of antibiotic resistance. Microbiol. Mol. Biol. Rev. 2010, 74, 417–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peeters, J.E.; Maertens, L.; Orsenigo, R.; Colin, M. Influence of dietary beet pulp on caecal VFA, experimental colibacillosis and iota-enterotoxaemia in rabbits. Anim. Feed Sci. Tech. 1995, 51, 123–139. [Google Scholar] [CrossRef]
- Dayrit, F.M. The properties of lauric acid and their significance in coconut oil. J. Am. Oil Chem. Soc. 2015, 92, 1–15. [Google Scholar] [CrossRef]
- Kylie, K.; Weese, J.S.; Turner, P.V. Comparison of the fecal microbiota of domestic commercial meat, laboratory, companion, and shelter rabbits (Oryctolagus cuniculi). BMC Vet. Res. 2018, 14, 143. [Google Scholar] [CrossRef]
- Fang, S.; Chen, X.; Zhou, L.; Wang, C.; Chen, Q.; Lin, R.; Xiao, T.; Gan, Q. Faecal microbiota and functional capacity associated with weaning weight in meat rabbits. Microb. Biotechnol. 2019, 12, 1441–1452. [Google Scholar] [CrossRef]
- Sudo, N. Role of gut microbiota in brain function and stress-related pathology. Biosci. Microbiota. Food. Health 2019, 38, 75–80. [Google Scholar] [CrossRef] [Green Version]
- Velasco-Galilea, M.; Piles, M.; Vinas, M.; Rafel, O.; Gonzalez-Rodriguez, O.; Guivernau, M.; Sánchez, J.P. Rabbit microbiota changes throughout the intestinal tract. Front. Microbiol. 2018, 9, 2144. [Google Scholar] [CrossRef]
- Chen, S.Y.; Deng, F.; Jia, X.; Liu, H.; Zhang, G.W.; Lai, S.J. Gut microbiota profiling with differential tolerance against the reduced dietary fibre level in rabbit. Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Dabbou, S.; Ferrocino, I.; Kovitvadhi, A.; Dabbou, S.; Bergagna, S.; Dezzuto, D.; Schiavone, A.; Cocolin, L.; Gai, F.; Santoro, V.; et al. Bilberry pomace in rabbit nutrition: Effects on growth performance, apparent digestibility, caecal traits, bacterial community and antioxidant status. Animal 2019, 13, 53–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, Z.L.; Wu, G.; Zhu, W.Y. Amino acid metabolism in intestinal bacteria: Links between gut ecology and host health. Front. Biosci. 2011, 16, 1768–1786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- North, M.K.; Dalle Zotte, A.; Hoffman, L. Composition of rabbit caecal microbiota and the effects of dietary quercetin supplementation and sex thereupon. World Rabbit Sci. 2019, 27, 185–198. [Google Scholar] [CrossRef] [Green Version]
- Dao, M.C.; Everard, A.; Aron-Wisnewsky, J.; Sokolovska, N.; Prifti, E.; Verger, E.O.; Kayser, B.D.; Levenez, F.; Chilloux, J.; Hoyles, L.; et al. Akkermansia muciniphila and improved metabolic health during a dietary intervention in obesity: Relationship with gut microbiome richness and ecology. Gut 2016, 65, 426–436. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Gallego, C.; Pohl, S.; Salminen, S.; De Vos, W.M.; Kneifel, W. Akkermansia muciniphila: A novel functional microbe with probiotic properties. Benef. Microbes 2016, 7, 571–584. [Google Scholar] [CrossRef]
- Zhai, Q.; Feng, S.; Arjan, N.; Chen, W. A next generation probiotic, Akkermansia muciniphila. Crit. Rev. Food Sci. Nutr. 2019, 59, 3227–3236. [Google Scholar] [CrossRef]
- Dunfield, P.F.; Yuryev, A.; Senin, P.; Smirnova, A.V.; Stott, M.B.; Hou, S.; Ly, B.; Saw, J.H.; Zhou, Z.; Ren, Y.; et al. Methane oxidation by an extremely acidophilic bacterium of the phylum Verrucomicrobia. Nature 2007, 450, 879–882. [Google Scholar] [CrossRef]
- Martinez-Garcia, M.; Brazel, D.M.; Swan, B.K.; Arnosti, C.; Chain, P.S.G.; Reitenga, K.G.; Xie, G.; Poulton, N.J.; Gomez, M.L.; Masland, D.E.D.; et al. Capturing single cell genomes of active polysaccharide degraders: An unexpected contribution of verrucomicrobia. PLoS ONE 2012, 7, e35314. [Google Scholar] [CrossRef] [Green Version]
- Wertz, J.T.; Kim, E.; Breznak, J.A.; Schmidt, T.M.; Rodrigues, J.L. Genomic and physiological characterization of the Verrucomicrobia isolate Diplosphaera colitermitum gen. Nov., sp. nov., reveals microaerophily and nitrogen fixation genes. Appl. Environ. Microbiol. 2012, 78, 1544–1555. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Finegold, S.M.; Song, Y.; Lawson, P.A. Reclassification of Clostridium coccoides, Ruminococcus hansenii, Ruminococcus hydrogenotrophicus, Ruminococcus luti, Ruminococcus productus and Ruminococcus schinkii as Blautia coccoides gen. nov., comb. nov., Blautia hansenii comb. nov., Blautia hydrogenotrophica comb. nov., Blautia luti comb. nov., Blautia producta comb. nov., Blautia schinkii comb. nov. and description of Blautia wexlerae sp. nov., isolated from human faeces. Int. J. Sys. Evol. Microbiol. 2008, 58, 1896–1902. [Google Scholar]
- Chen, J.; Zhang, H.; Wu, X.; Shang, S.; Yan, J.; Chen, Y.; Zhang, H.; Tang, X. Characterization of the gut microbiota in the golden takin (Budorcas taxicolor bedfordi). AMB Express 2017, 7, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gulino, L.M.; Ouwerkerk, D.; Kang, A.Y.; Maguire, A.J.; Kienzle, M.; Klieve, A.V. Shedding light on the microbial community of the macropod foregut using 454-amplicon pyrosequencing. PLoS ONE 2013, 8, e61463. [Google Scholar] [CrossRef]
- Tims, S.; Derom, C.; Jonkers, D.M.; Vlietinck, R.; Saris, W.H.; Kleerebezem, M.; De Vos, W.M.; Zoetendal, E.G. Microbiota conservation and BMI signatures in adult monozygotic twins. ISME J. 2013, 7, 707–717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clark, A.; Mach, N. Exercise-induced stress behavior, gut-microbiota-brain axis and diet: A systematic review for athletes. J. Int. Soc. Sports Nutr. 2016, 13, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gosalbes, M.J.; Durbán, A.; Pignatelli, M.; Abellan, J.J.; Jiménez-Hernández, N.; Pérez-Cobas, A.E.; Latorre, A.; Moya, A. Metatranscriptomic approach to analyze the functional human gut microbiota. PLoS ONE 2011, 6, e17447. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Diets | |||
|---|---|---|---|
| Ingredients | C | HID | TMD |
| Dehydrated alfalfa meal (17 g CP/100 g) | 32 | 32 | 32 |
| Alfalfa hay | 7.5 | 7.5 | 7.5 |
| Wheat bran | 23.5 | 23.5 | 23.5 |
| Barley meal | 10 | 10 | 10 |
| Dried sugar beet pulp | 16 | 16 | 16 |
| Soybean meal (44 g CP/100 g) | 7 | 7 | 7 |
| Soybean oil | 1.5 | - | - |
| Hermetia illucens fat | - | 1.5 | - |
| Tenebrio molitor fat | - | - | 1.5 |
| Cane molasses | 1.2 | 1.2 | 1.2 |
| Dicalcium phosphate | 0.3 | 0.3 | 0.3 |
| Salt | 0.4 | 0.4 | 0.4 |
| L–methionine (98 g methionine/100 g) | 0.1 | 0.1 | 0.1 |
| Vitamin–mineral premixa | 0.5 | 0.5 | 0.5 |
| Chemical Composition | |||
| Dry matter, % | 89.4 | 89.2 | 89.6 |
| Ash, % DM | 8.58 | 7.77 | 7.75 |
| Crude protein, % DM | 17.0 | 16.8 | 16.3 |
| Ether extract, % DM | 4.22 | 3.92 | 3.87 |
| Neutral detergent fiber (aNDF), % DM | 40.2 | 41.7 | 40.5 |
| Acid detergent fiber (ADF), % DM | 21.7 | 23.0 | 22.8 |
| Acid detergent lignin (ADL), % DM | 4.81 | 5.09 | 5.02 |
| Gross energy, MJ/kg DM | 18.50 | 18.50 | 18.62 |
| Fatty acids | Dietary Fats | Experimental Diets | ||||
|---|---|---|---|---|---|---|
| S | HI | TM | C | HID | TMD | |
| C12:0 | 0.2 | 480 | 2.3 | 0.5 | 203 | 3.0 |
| C14:0 | 0.5 | 103 | 22.2 | 0.9 | 44.7 | 13.3 |
| C16:0 | 104 | 127 | 176 | 121 | 161 | 184 |
| C18:0 | 44.3 | 19.0 | 23.1 | 28.4 | 20.8 | 22.2 |
| BCFAs | 0.1 | 2.9 | 0.8 | 2.4 | 4.1 | 3.4 |
| C16:1 n-7 | 0.9 | 32.0 | 16.6 | 0.12 | 19.9 | 10.4 |
| C18:1 n-9 | 230 | 91.1 | 378 | 201 | 127 | 273 |
| C18:2 n-6 | 515 | 90.0 | 332 | 521 | 310 | 389 |
| C18:3 n-3 | 70.3 | 10.1 | 18.0 | 74.3 | 62.8 | 55.1 |
| SFA1 | 158 | 748 | 231 | 165 | 454 | 240 |
| UFA1 | 842 | 252 | 769 | 835 | 546 | 760 |
| MUFA1 | 254 | 141 | 411 | 236 | 169 | 309 |
| PUFA1 | 588 | 111 | 358 | 599 | 377 | 451 |
| ∑n3 | 70.5 | 11.7 | 18.3 | 74.7 | 62.8 | 55.1 |
| ∑n6 | 516 | 91.1 | 333 | 523 | 311 | 391 |
| Growth Conditions | Pasteurella multocida | |||||
|---|---|---|---|---|---|---|
| T4 | T6 | T8 | T10 | T12 | T24 | |
| Control (A) | 4.09 (0.19) | 4.14 (0.06) | 4.76 (0.32)—A | 4.80 (0.34)—A | 5.69 (0.49)—A | 6.82 (0.44)—A |
| Control (B) | 4.11 (0.17) | 3.96 (0.12) | 4.17 (0.26)—A | 4.60 (0.40)—A | 5.44 (0.57)—A | 6.54 (0.39)—A |
| TSB + TM fat | 3.95 (0.44) | 3.75 (0.39) | 3.08 (0.35)—B | 3.21 (0.30)—B | 2.91 (0.86)—B | 3.90 (1.21)—B |
| TSB + HI fat | 3.92 (0.20) | 3.48 (0.17) | 3.61 (0.19)—B | 2.80 (0.59)—B | 2.67 (0.49)—B | 2.06 (0.36)—C |
| ANOVA | N.S | N.S | F = 4.99; p = 0.01 | F = 5.07; p = 0.01 | F = 6.36; p < 0.01 | F = 13.75; p < 0.01 |
| Yersinia enterocolitica | ||||||
| Control (A) | 4.36 (0.16)—A | 5.19 (0.28)—A | 6. 76 (0.61)—A | 7.32 (0.37)—A | 8.18 (0.37)—A | 9.95 (0.30)—A |
| Control (B) | 4.10 (0.21)—A | 5.05 (0.37)—A | 5.91 (0.41)—A | 6.83 (0.48)—A | 7.35 (0.68)—A | 9.88 (0.33)—A |
| BHI + TM fat | 4.05 (0.31)—A | 4.39 (0.52)—AB | 4.98 (0.74)—AB | 5.04 (0.94)—AB | 5.93 (1.41)—AB | 8.09 (2.13)—A |
| BHI + HI fat | 3.34 (0.10)—B | 2.69 (0.64)—B | 3.01 (1.20)—B | 3.02 (1.32)—B | 2.94 (1.24)—B | 3.98 (2.18)—B |
| ANOVA | F = 3.28; p = 0.05* | F = 6.38; p < 0.01 | F = 4.95; p = 0.01 | F = 7.71; p < 0.01 | F = 6.46; p < 0.01 | F = 5.42; p < 0.01 |
| Listeria monocytogenes | ||||||
| Control (A) | 4.58 (0.35) | 5.58 (0.39) | 6.47 (0.41)—A | 7.41 (0.41)—A | 8.28 (0.46)—A | 10.00 (0.26)—A |
| Control (B) | 4.52 (0.32) | 5.45 (0.42) | 6.26 (0.54)—A | 6.94 (0.61)—A | 7.95 (0.59)—A | 9.96 (0.28)—A |
| BHI + TM fat | 4.20 (0.25) | 4.59 (0.79) | 5.98 (0.46)—A | 6.76 (0.70)—A | 7.63 (0.61)—A | 9.48 (0.03)—A |
| BHI + HI fat | 4.00 (0.92) | 3.66 (0.84) | 4.1 (0.63)—B | 4.48 (0.61)—B | 4.51 (0.62)—B | 4.85 (0.97)—B |
| ANOVA | N.S | N.S | F = 3.30; p = 0.05** | F = 3.98; p = 0.03 | F = 6.85; p < 0.01 | F = 29.92; p < 0.01 |
| C | HID | TMD | SEM | p-Value | |
|---|---|---|---|---|---|
| pH | 6.1 | 5.9 | 5.9 | 0.01 | 0.15 |
| N–NH3 (mmol/L) | 2.2 | 3.0 | 3.1 | 0.23 | 0.25 |
| Total VFA (mmol/L) | 72.4b | 85.3a | 83.9a | 2.31 | 0.03 |
| Acetic acid (C2; mmol/100 mmol VFA) | 77.8 | 78.1 | 76.6 | 0.43 | 0.30 |
| Propionic acid (C3; mmol/100 mmol VFA) | 5.3 | 5.0 | 5.4 | 0.19 | 0.71 |
| Butyric acid (C4; mmol/100 mmol VFA) | 16.2 | 16.1 | 17.2 | 0.42 | 0.53 |
| Valeric acid (C5; mmol/100 mmol VFA) | 0.5 | 0.4 | 0.5 | 0.03 | 0.59 |
| Caproic acid (C6; mmol/100 mmol VFA) | 0.3 | 0.3 | 0.3 | 0.03 | 0.45 |
| C2/C3 ratio | 15.2 | 16.0 | 14.6 | 0.54 | 0.58 |
| C2/C4 ratio | 4.9 | 5.0 | 4.6 | 0.16 | 0.63 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dabbou, S.; Ferrocino, I.; Gasco, L.; Schiavone, A.; Trocino, A.; Xiccato, G.; Barroeta, A.C.; Maione, S.; Soglia, D.; Biasato, I.; et al. Antimicrobial Effects of Black Soldier Fly and Yellow Mealworm Fats and Their Impact on Gut Microbiota of Growing Rabbits. Animals 2020, 10, 1292. https://doi.org/10.3390/ani10081292
Dabbou S, Ferrocino I, Gasco L, Schiavone A, Trocino A, Xiccato G, Barroeta AC, Maione S, Soglia D, Biasato I, et al. Antimicrobial Effects of Black Soldier Fly and Yellow Mealworm Fats and Their Impact on Gut Microbiota of Growing Rabbits. Animals. 2020; 10(8):1292. https://doi.org/10.3390/ani10081292
Chicago/Turabian StyleDabbou, Sihem, Ilario Ferrocino, Laura Gasco, Achille Schiavone, Angela Trocino, Gerolamo Xiccato, Ana C. Barroeta, Sandra Maione, Dominga Soglia, Ilaria Biasato, and et al. 2020. "Antimicrobial Effects of Black Soldier Fly and Yellow Mealworm Fats and Their Impact on Gut Microbiota of Growing Rabbits" Animals 10, no. 8: 1292. https://doi.org/10.3390/ani10081292