1. Introduction
In modern ruminant production systems, to achieve maximum performance, the inclusion of plentiful amounts of concentrate in the diet is commonly practiced in the intensive feedlot management system of dairy cows and goats [
1,
2]. Relatively high fiber feeding and stable microbial community are the indispensable conditions for the health of ruminants [
3]. However, at present, the high-concentrate (HC) diet is widely adopted to provide adequate protein and energy supply for meeting higher performance needs at the finishing stage of ruminants [
4]. Appropriate increase of dietary concentrate level can improve the performance of ruminants [
5], whereas high inclusion rates of concentrates affects the conditions of microorganisms inhabiting the gastro-intestines, including the acidity, osmolality, and the contents of fermentable substrates [
6,
7]. Dietary starch content can affect the rumen and hindgut bacteria when raising dietary concentrate proportion, but these effects vary greatly among ruminant species [
8,
9].
The animal digestive tract is colonized by a dense, dynamic and highly complex community of microorganisms composed mainly of bacteria, whose total number exceeds 10
14 cells, with many unique strains [
10]. Intestinal microflora play a key role in nutrient metabolism and development of the immune system [
11,
12]. Among the elements affecting bacteria in the intestine, a major factor is the ability to utilize the available nutrients to achieve high growth rates to avoid washout and appease a reaction-ready immune system [
13]. The large amount of carbohydrates promotes the growth of bacteria in the small intestine [
14]. In contrast, in the hindgut, plenty of nutrients for bacteria are derived from undigested carbohydrates and resistant starch as well as undigested protein in the diets. There these nutrients undergo microbial fermentation, resulting in the production of metabolites, such as VFA (volatile fatty acid) [
15]. Many previous studies have revealed that an increasing fermentation in the rumen and hindgut typically occurs concurrently with HC feeding and increments of VFA and LPS (lipopolysaccharide), as well as reduced pH in the hindgut of ruminants, causing an unhealthy gastrointestinal environment [
16,
17]. Such studies were necessary in order to enhance the understanding of the relationship between HC diet and the intestinal bacteria of goats, as this would lay a foundation for the development of strategies to prevent this disorder.
The development and application of Illumina MiSeq sequencing methods may help to explain the underlying mechanism of comprehensive variation of intestinal bacteria caused by feeding HC diet. Our previous studies have noted that feeding a HC diet causes subacute ruminal acidosis [
18] and alters total VFA production in the ileum [
19], indicating that a HC diet challenge increases the risk of poor gastrointestinal health. Therefore, in the present study, we hypothesized that a HC diet might cause changes in the ileal metabolic activities and bacterial community, and further induce ileal epithelial injury in goats. The objective of this study was to investigate the changes in the ileal microbiota, metabolites, biochemical parameters and histomorphology during HC feeding. Additionally, the relationship among alterations in the ileal bacterial composition and metabolites and biochemical indices in the intestinal digesta were also evaluated. The reason for choosing rice grain as the concentrate supplements was that rice is planted on a large scale in the south of China.
4. Discussion
The influence of feeding goats with rice on the gut micro-environment and intestinal health is still unknown. In previous studies by our group and others, the HC diet appeared to disrupt the balance of the ruminal microbiota in ruminants by Illumina MiSeq sequencing methods [
9,
24,
25]. However, little information was available on microbial response of the small intestine to the HC diet using a high-throughput method [
26]. In the present study, we used high-throughput 16S rRNA gene sequencing to assess the shifts in the microbial adaptation of goat’s ileum in response to a HC diet. Our data provide a detailed picture of the ileal metabolites related to microorganisms in goats.
The intestinal microbial community imparts specific functions in the host such as nutrient absorption, drug metabolism, maintenance of structural integrity of the gut barrier, immunoregulation, and protection against pathogens [
27]. In accordance with a previous study which reported that feeding an HC diet could reduce the bacterial richness and functionality of the microbiota in the digestive tract of ruminants [
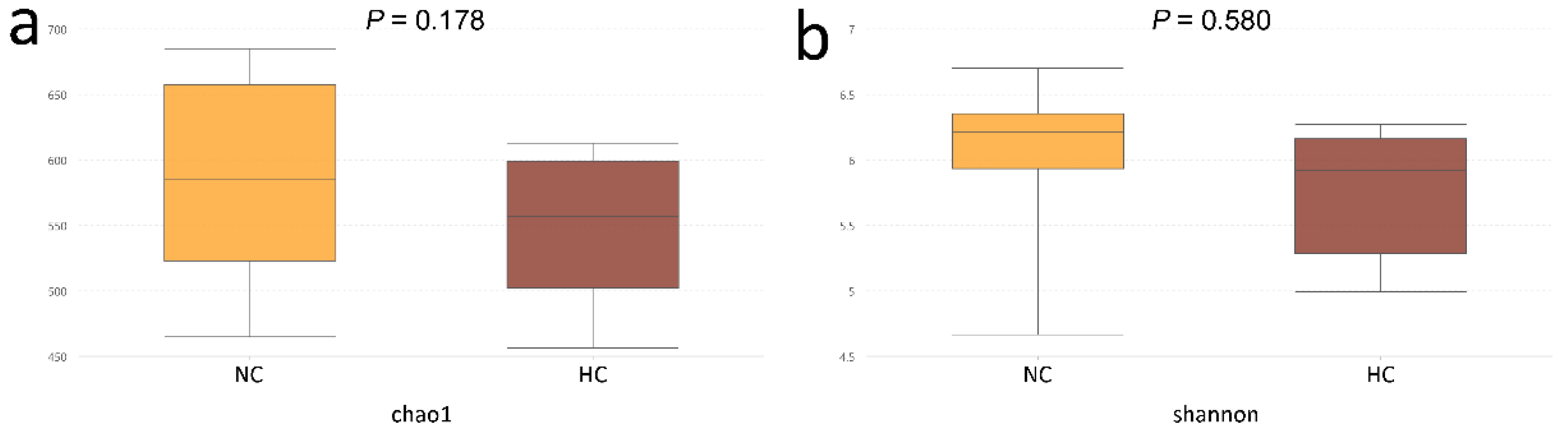
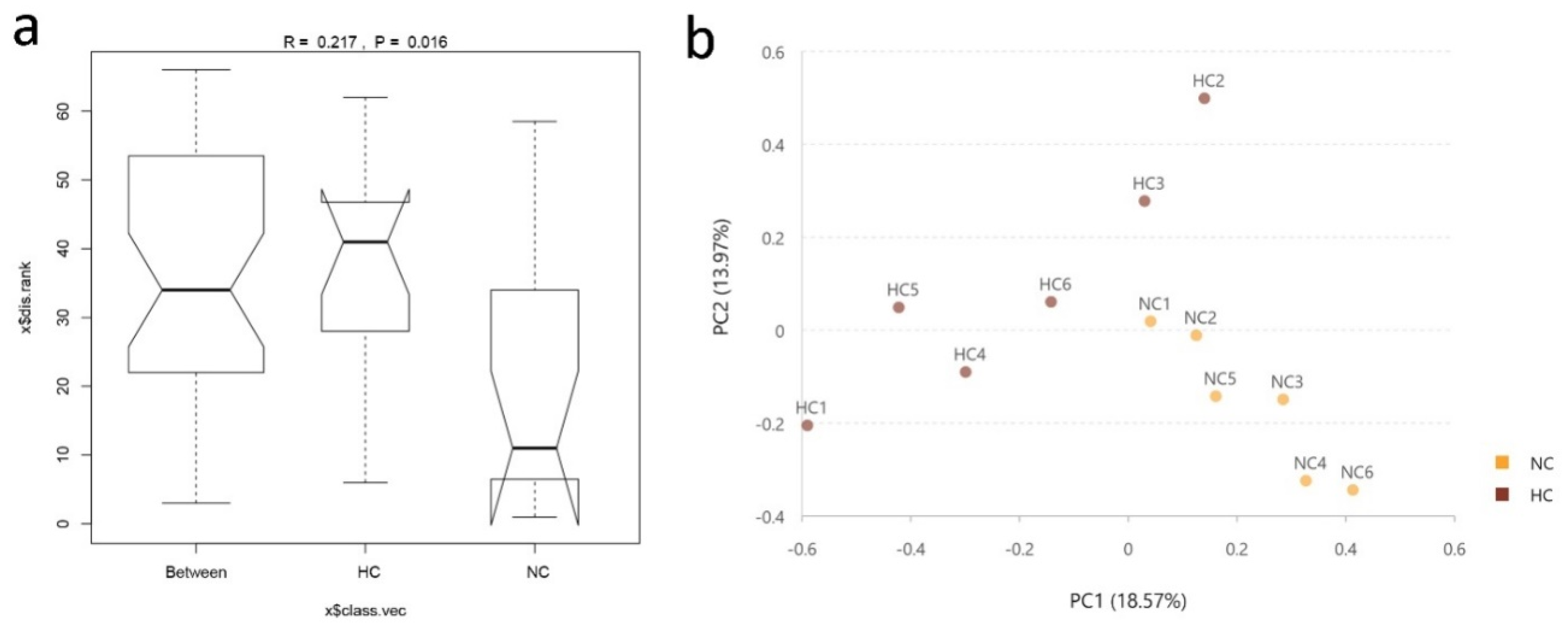
28], the Chao1 and Shannon values in our results showed a sharp decline in the ileum of goats fed an HC diet. In addition, the results of the PCA and Anosim analyses further revealed the difference in bacterial diversity composition between the NC and HC groups, also indicated that feeding a HC diet changed the bacterial diversity in the ileum.
In agreement with earlier studies of the intestinal bacterial communities of goats [
17,
19], our results revealed that the Firmicutes was the major phylum in the ileal digesta of goats. Bacteria belonging to the Firmicutes are known for a fermentative metabolism and degradation of carbon sources, protein and amino acid [
29]. Thus, the high abundance of Firmicutes in ileal digesta bacteria emphasized that Firmicutes played a role in the utilization of carbohydrates, protein and amino acid. Tao et al. [
30] deduced that the second-most abundant bacterial community, Bacteroidetes of Gram-negative bacteria were increased in the hindgut by a HC diet. However, in the current research, Tenericutes was the second most abundant bacterial community in the ileum. It is possible that different digestive location or diet composition has different microbial flora, because it is known that the composition of the gut microbial community is related to the diet type or host phylogeny [
31]. It was previously found that LPS was produced abundantly in the digesta derived from Bacteroides spp. [
32]. In the current study, the Bacteroides population was less than 1%. Maybe that could explain why there was no difference in LPS concentration in the ileal digesta. Jenkins et al. [
33] reported that species belonging to the family of Christensenellaceae were common in the rumen and that these species played a key role in preserving gastrointestinal structure and function. Compared with the NC group, we detected no difference in proportion of the Christensenellaceae_R-7_group (Family, butyrate producer) in ileal microbial response of goats to the HC diet. Besides, our results somehow showed no difference in butyrate concentration between the two groups. Generally speaking, Cyanobacteria phylum is not regarded as intestinal bacteria in goats although it had been detected previously in human and other animal gut [
34]. In this study, the abundance of the Cyanobacteria phylum had a downward trend. Ley et al. [
35] reported that there was a logical intestine-associated branch rooted deep in the Cyanobacteria group, and this group may represent descendants of non-photosynthetic ancestral cyanobacteria that have adapted to life in the gastrointestinal tract of animals.
In the present study, a great number of Ruminococcaceae was found in the ileum digesta of the HC group. This is not consistent with the findings recorded in the cecum of goats fed a high-grain diet [
36]. This difference may be due to different responses of ileal microbes to diets, and the HC diet provided suitable nutrient conditions for Ruminococcaceae to grow [
37], resulting in its rise.
Clostridium is well known as an intestinal colonizer. It has been reported that some
Clostridium spp., as opportunistic pathogens, are causative agents of intestinal enteric diseases in goats [
38]. Moreover,
Clostridium phylotypes can negatively impact the intestinal barrier [
39]. An increase in the richness of
Clostridium_
sensu_
stricto_1 in the present study hints that feeding a HC diet may have a harmful effect on the ileal health of goats. Liu et al. [
36] found that a HC diet in goats increased the population of Turicibacter, Treponema and Prevotella in the cecum. Turicibacter in the gut may cause subclinical infection or have some other adverse reactions on the gastrointestinal tract [
40], however, the present study revealed that there was no difference in Turicibacter, and Treponema and Prevotella were not detected in the ileum of goats. In addition, HC diet contained a greater number of
Ruminococcus_
gauvreauii_group,
Ruminococcus 1, and
Ruminococcus_1 in the ileal digesta than that of the NC group.
Ruminococcus is the most dominant genus found in the large intestine of healthy sheep [
41] and plays a vital role in degrading starch [
42,
43]. So, the
Ruminococcus spp. elevation in the ileum digesta may suggest that HC diet improves the ability of the ileal bacteria to digest starch.
It has been revealed that the gut microbiota makes an essential functional contribution in maintaining the health of the gastrointestinal tract for the host [
44,
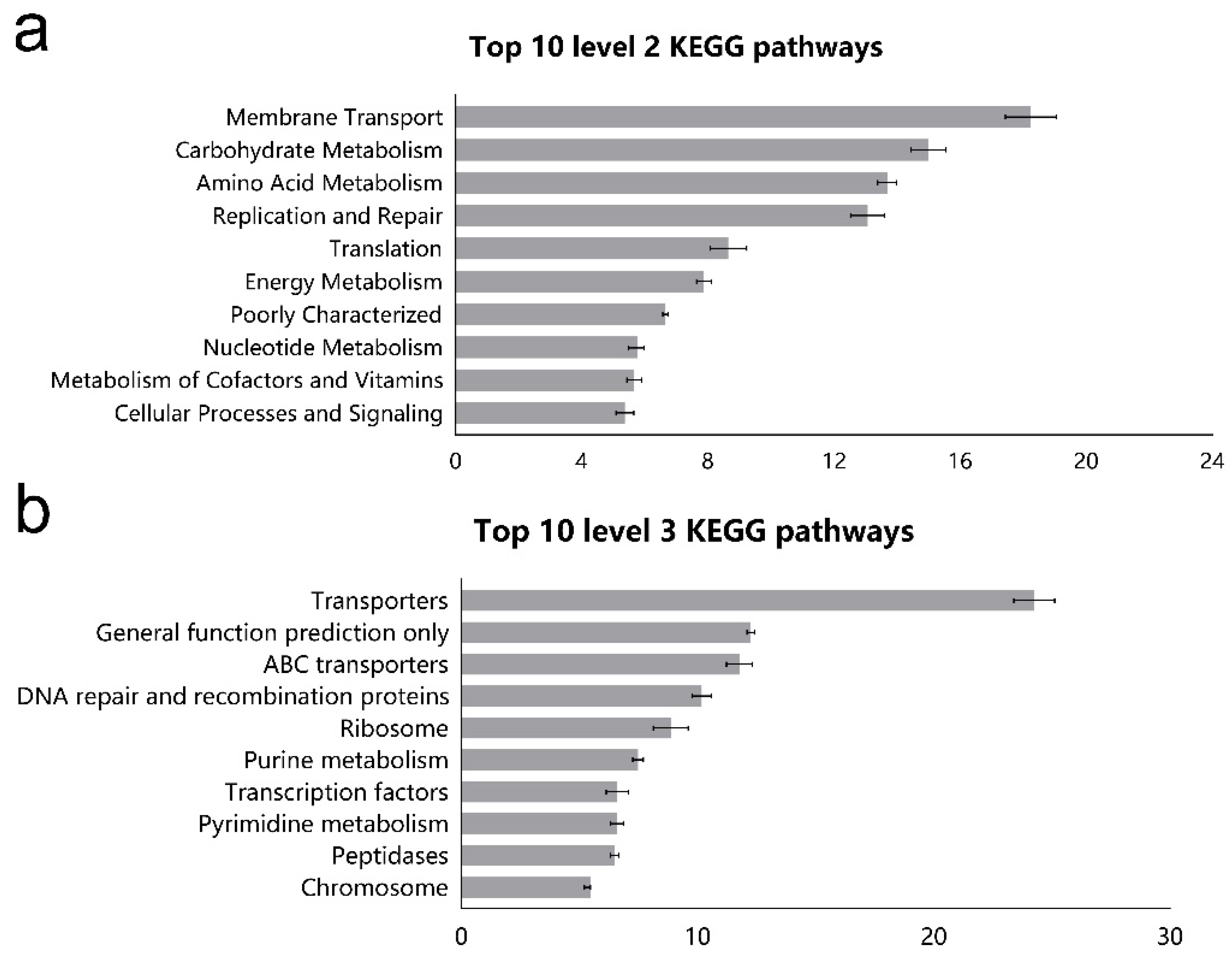
45]. Our results showed that the most abundant gene categories in the ileum of goats were related to the function of membrane transport, amino acid metabolism, and energy metabolism, which is similar to the results of previous studies that were conducted in sheep [
23]. The system of membrane transport is essential to communicate with the tissues and environment and import molecules into cells and export waste from the cells [
46]. PICRUSt from the current study suggested that the relative abundance of genes associated with amino acid metabolism, carbohydrate metabolism, and lipid metabolism was up-regulated by HC diet, indicating that HC diet enhanced the metabolic function of microbes. This shift of coordination in gene expression implies a selective change in metabolic pathways favoring the use of carbohydrates as fuel to sustain energy expenditure for the ileal microflora. Besides, the ileal microflora was predicted to have greater capabilities for replication and repair, and this may be due to the rapid turnover rate of ileal bacteria. Small-intestinal microbiota are proven to be the key sensor of dietary signals that allow the host to adapt to variations in lipid digestion and absorption, and a reference strain
Clostridium genus can increase oleic acid uptake and the expression of genes involved in triglyceride synthesis [
47]. Thus, the observed increment in the
Clostridium_
sensu_
stricto_1 may be related to regulating lipid metabolism in the ileum of goats. The nicotinate and nicotinamide metabolism and vitamin B6 metabolism are essential to maintain sufficient biological response involved in the metabolism of carbohydrates, lipids and proteins [
48]. So, we observed that the enhanced vitamin-related pathway was accompanied by boosting the metabolic activity of major nutrients. In fact, vitamins cannot be synthesized by the animal body but can be synthesized by commensal bacteria [
49], and vitamins in the ileum mainly originate from the diet and commensal microorganisms. An increase in
Clostridium abundance, which represents vitamin producers, also confirmed this viewpoint [
50]. In addition, the HC diet altered the ileal microorganism populations in particular carbohydrate and protein degraders such as
Clostridium_
sensu_
stricto_1 and some Ruminococcaceae spp. (
Eubacterium_
coprostanoligenes_group,
Ruminococcus 1 and
Ruminococcus_1). So ileal microbial response to a HC diet involved upregulation of most genes involved in metabolism of macronutrients (carbohydrates, lipids, and proteins) and micronutrients (vitamins), reflecting an augmentation of the metabolic activity in the ileum.
Previous studies on VFA in the rumen and hindgut of ruminants have explored feeding a HC diet have been explored [
16,
17], however, little information is available on changes of VFA pattern in the ileum during digestion of a HC diet. Our previous research revealed that the HC diet decreased the pH and increased TVFA concentrations in the ileum [
19]. Interestingly, about a two-fold increase in TVFA concentrations was detected in the HC group compared with the NC group. Thus, we inferred that the higher VFA in the ileum of HC-fed goats may be due to rapid degradation of carbohydrates or the excessive VFA escaped from the rumen into the ileum of goats. The over-fermentation of VFA in hindgut increases the osmotic pressure of digesta, and the high osmotic pressure partially damages the intestinal epithelium [
16], so the damage may be similar in the ileum. Studies of Steele et al. [
51] and Liu et al. [
52] have shown that feeding ruminants with a HC diet will increase the concentration of LPS in the rumen and cause ruminal epithelial damage. Besides, the intestinal epithelium is composed of only a single layer of epithelial cells, whereas the rumen epithelium consists of a multilayered squamous epithelium [
6]. Thus, differences between the structures of the ileal and ruminal epithelia are likely to make the ileal epithelium more susceptible to damage than the rumen responding to the HC diet. Nonetheless, Zebeli et al. [
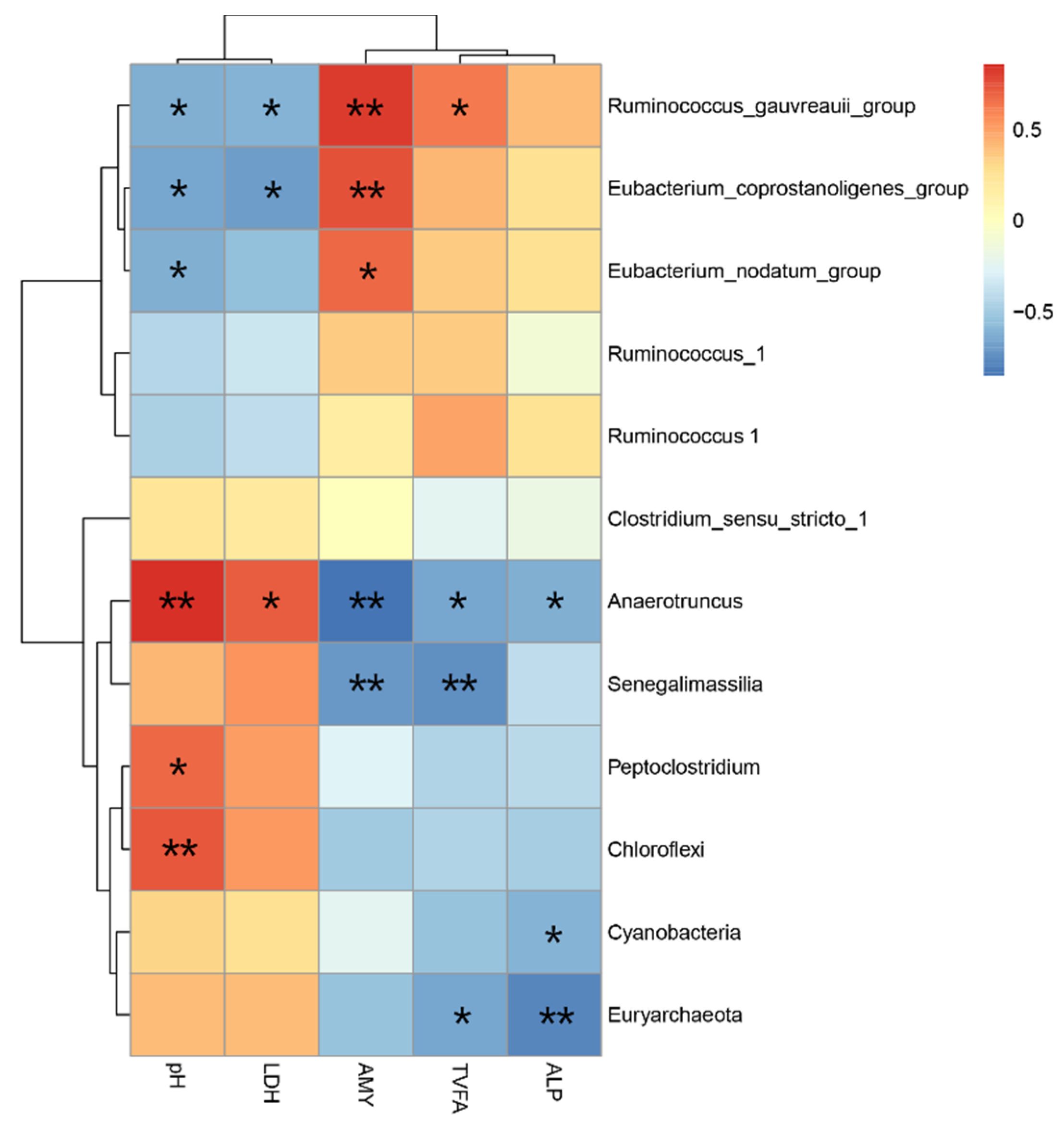
53] indicated that LPS is released by Gram-negative bacteria after lysis at low pH and there were no significantly negative correlations between the concentration of LPS in the ileum and the proportion of Gram-negative bacteria. These findings demonstrate that an HC diet feeding results in a low ileal pH and that the latter could not lead to the death and lysis of Gram-negative bacteria in the ileum, therefore, it had not increased the concentration of LPS in the ileum. In addition, the elevated level of enzymes (AST, ALT, and ALP) indicated some damage or changes in membrane permeability [
54], rather than damage to the intestinal villus and crypt in the HC group. The abundance of the genera Anaerotruncus, Cyanobacteria, and Euryarchaeota was negatively correlated with ALP activity, suggesting that they might be involved in the ALP metabolism of the ileum. However, what role these genera played in the intestinal barrier needs to be clarified. The LDH is released during tissue damage and as a marker of pathological changes. In particular, feedback inhibition by LDH can reduce the conversion of pyruvate to LACT at high lactate concentration [
55]. Our data display no significant difference in ileal LACT concentration between treatments and did not detect LDH in the HC diet. Maybe the LDH is all used to neutralize LACT and further investigation is needed to explain this phenomenon. AMY activity of fermenting microorganisms is an important factor in the fermentation of starch to lactate [
56]. Previous studies have demonstrated that the concentration of AMY was related to dry matter intake and nutrient composition, and more starch intake may enhance the AMY concentration in the small intestine [
57,
58]. In this study, we found that the HC diet significantly increased the activity of AMY, and
Eubacterium_
coprostanoligenes_group,
Eubacterium_
nodatum_group and
Ruminococcus_
gauvreauii_group were positively correlated with the activity of AMY in the ileum. These results indicate that more starch may flow into the ileum from the rumen due to the HC diet.
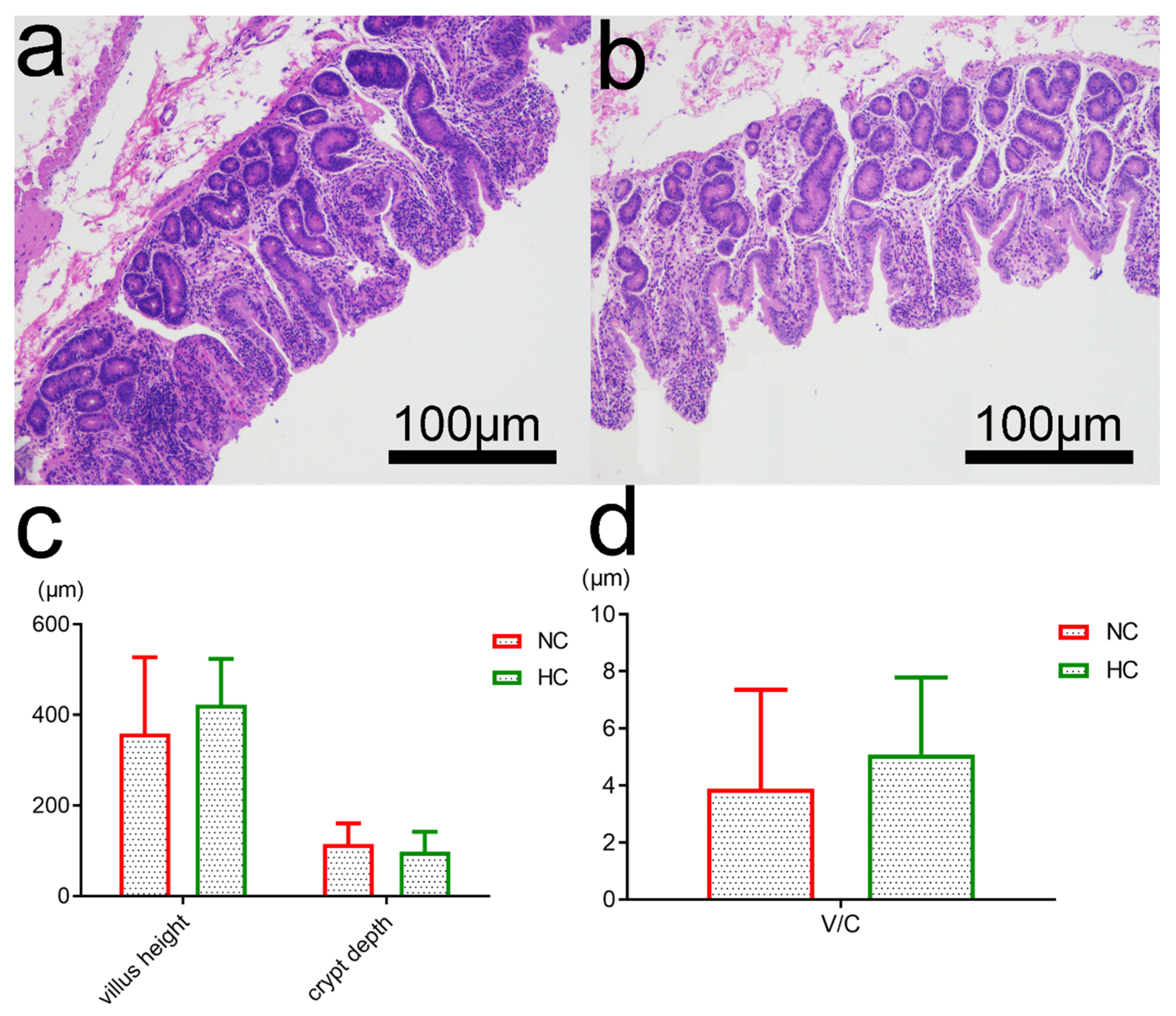
Previous studies have revealed that feeding a HC diet to ruminants caused a high risk of damage to the histological integrity and functions of the ruminal epithelium in ruminants [
51,
59]. However, there are few reports on the influence of a HC diet on ileal epithelial structure and bacterial function. In this study, after five weeks of feeding, the ileal epithelial structure of HC-fed goats was changed slightly. The HC diet did not significantly alter the villus height and crypt depth of ileum and caused only little sloughing of the epithelial structure. Overall, the findings indicate that, in the small intestine of goats fed with a HC diet, a slightly damaged epithelial barrier function may be associated with changes in VFA fermentation, enzyme metabolism and composition of microbiota. Therefore, these findings indicate that the HC diet may have a potentially adverse impact on ileal function.

 ,
, 






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}