Determination of Fatty Acids Profile in Original Brown Cows Dairy Products and Relationship with Alpine Pasture Farming System


 , ,
, , 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Farm Conditions, Animals, and Cheesemaking Procedure
2.2. Milk and Cheese Sampling
2.3. Milk and Cheese Samples Analysis
2.4. Statistical Analysis
3. Results
3.1. Milk and Cheese Fatty Acids Composition
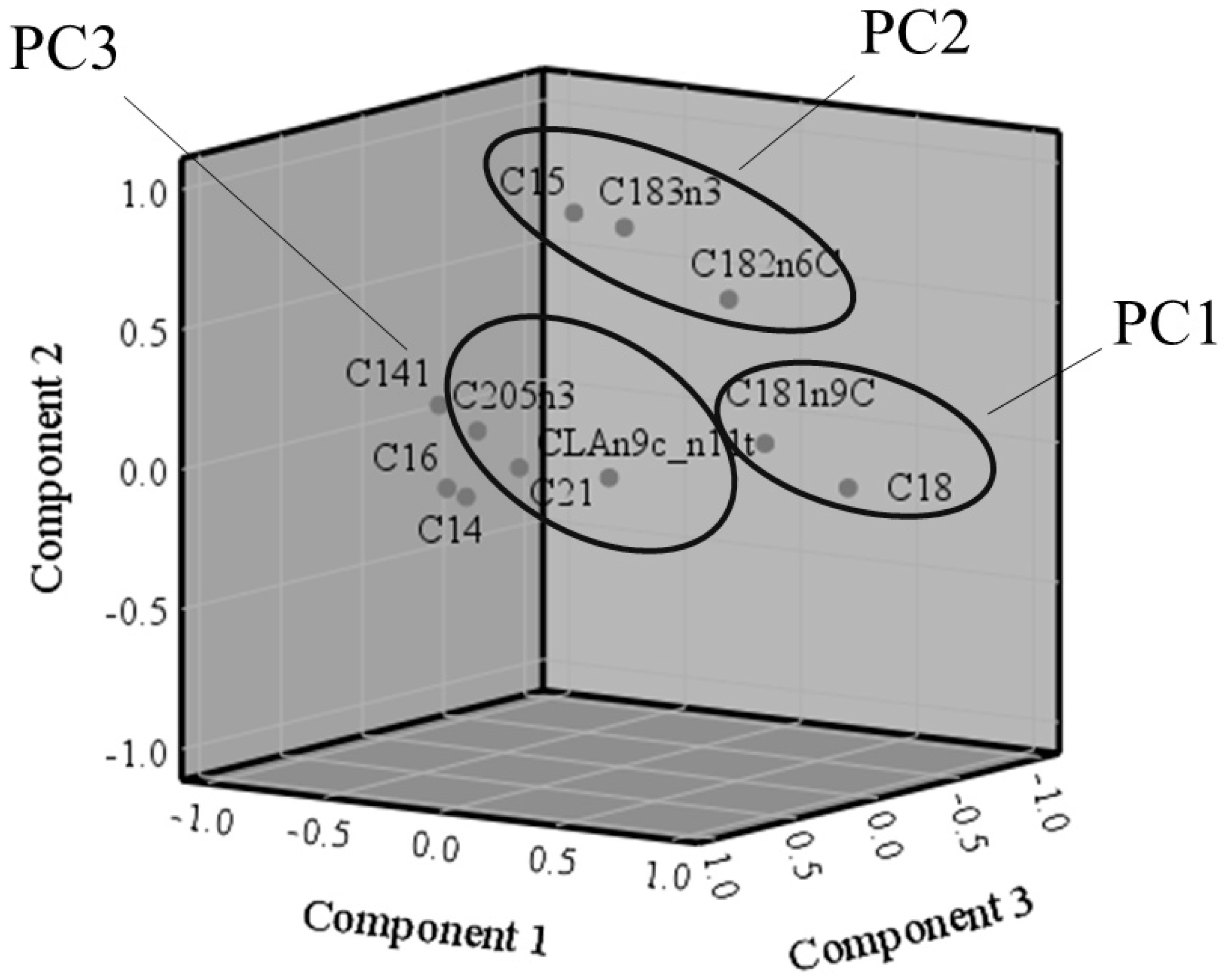
3.2. Principal Component Analysis
3.3. Characterization of Fatty Acid Profile on Milk and Cheese from Grazing and No Grazing Dairy Farming Systems
4. Discussion
4.1. Relationships between Different Fatty Acids in Milk and Cheese: The Principal Component Analysis
4.2. Characterization of Fatty Acids Profile on Milk and Cheese from Grazing and No Grazing Dairy Farming Systems
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Niedermayr, J.; Hoffmann, C.; Stawinoga, A.; Streifeneder, T. Agro-structural patterns in the Alps 2000–2010. OGA Jahrb. J. Austrian Soc. Agric. Econ. 2015, 24, 275–284. [Google Scholar]
- Bazzi, G.; Foglini, C.; Brambilla, M.; Saino, N.; Rubolini, D. Habitat management effects on Prealpine grassland bird communities. Ital. J. Zool. 2015, 82, 251–261. [Google Scholar] [CrossRef]
- Parolo, G.; Abeli, T.; Gusmeroli, F.; Rossi, G. Large-scale heterogeneous cattle grazing affects plant diversity and forage value of Alpine species-rich Nardus pastures. Grass Forage Sci. 2011, 66, 541–550. [Google Scholar] [CrossRef]
- Laiolo, P.; Dondero, F.; Ciliento, E.; Rolando, A. Consequences of pastoral abandonment for the structure and diversity of the alpine avifauna. J. Appl. Ecol. 2004, 41, 294–304. [Google Scholar] [CrossRef]
- Scarpa, R.; Notaro, S.; Louviere, J.; Raffaelli, R. Exploring scale effects of best/worst rank ordered choice data to estimate benefits of tourism in alpine grazing commons. Am. J. Agric. Econ. 2011, 93, 809–824. [Google Scholar] [CrossRef]
- Castrica, M.; Tedesco, D.E.A.; Panseri, S.; Ferrazzi, G.; Ventura, V.; Frisio, D.G.; Balzaretti, C.M. Pet food as the most concrete strategy for using food waste as feedstuff within the European context: A feasibility study. Sustainability 2018, 10, 2035. [Google Scholar] [CrossRef] [Green Version]
- Curone, G.; Filipe, J.; Cremonesi, P.; Trevisi, E.; Amadori, M.; Pollera, C.; Castiglioni, B.; Turin, L.; Tedde, V.; Vigo, D.; et al. Research in Veterinary Science What we have lost: Mastitis resistance in Holstein Friesians and in a local cattle breed. Res. Vet. Sci. 2018, 116, 88–98. [Google Scholar] [CrossRef] [PubMed]
- Curone, G.; Filipe, J.; Cremonesi, P.; Piccioli-Cappelli, F.; Trevisi, E.; Amadori, M. Relevance of the dairy cow biodiversity in the development of a profitable and environmentally sustainable livestock. CAB Rev. Perspect. Agric. Vet. Sci. Nutr. Nat. Resour. 2019, 14, 1–11. [Google Scholar] [CrossRef]
- Cremonesi, P.; Ceccarani, C.; Curone, G.; Severgnini, M.; Pollera, C.; Bronzo, V.; Riva, F.; Addis, M.F.; Filipe, J.; Amadori, M.; et al. Milk microbiome diversity and bacterial group prevalence in a comparison between healthy Holstein Friesian and Rendena cows. PLoS ONE 2018, 13, e0205054. [Google Scholar] [CrossRef] [Green Version]
- Matson, P.A.; Parton, W.J.; Power, A.G.; Swift, M.J. Agricultural intensification and ecosystem properties. Science 1997, 277, 504–509. [Google Scholar] [CrossRef] [Green Version]
- Tilman, D.; Cassman, K.G.; Matson, P.A.; Naylor, R.; Polasky, S. Agricultural sustainability and intensive production practices. Nature 2002, 418, 671–677. [Google Scholar] [CrossRef]
- Frison, E.A.; Cherfas, J.; Hodgkin, T. Agricultural biodiversity is essential for a sustainable improvement in food and nutrition security. Sustainability 2011, 3, 238–253. [Google Scholar] [CrossRef] [Green Version]
- Biscarini, F.; Nicolazzi, E.; Alessandra, S.; Boettcher, P.; Gandini, G. Challenges and opportunities in genetic improvement of local livestock breeds. Front. Genet. 2015, 6, 33. [Google Scholar] [CrossRef] [Green Version]
- Boettcher, P.J.; Hoffmann, I.; Baumung, R.; Drucker, A.G.; McManus, C.; Berg, P.; Stella, A.; Nilsen, L.; Moran, D.; Naves, M.; et al. Genetic resources and genomics for adaptation of livestock to climate change. Front. Genet. 2014, 5, 461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FAO. In vivo Conservation of Animal Genetic Resources; FAO: Rome, Italy, 2013; ISBN 0306-5251. (Print)r0306-5251 (Linking). [Google Scholar]
- FAO. The Second Report on the State of the World’s Animal Genetic Resources for Food and Agriculture; Rischkowsky, B., Pilling, D., Eds.; FAO: Rome, Italy, 2007; ISBN 9789251057629. [Google Scholar]
- Pietroboni, L.; Vento, R. Bruna linea originale. rb-la Razza Bruna 2017, 3, 23–24. [Google Scholar]
- Hagger, C. Estimates of genetic diversity in the brown cattle population of Switzerland obtained from pedigree information. J. Anim. Breed. Genet. 2005, 122, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Bieber, A.; Wallenbeck, A.; Leiber, F.; Fuerst-Waltl, B.; Winckler, C.; Gullstrand, P.; Walczak, J.; Wójcik, P.; Neff, A.S. Production level, fertility, health traits, and longevity in local and commercial dairy breeds under organic production conditions in Austria, Switzerland, Poland, and Sweden. J. Dairy Sci. 2019, 102, 5330–5341. [Google Scholar] [CrossRef]
- Bhati, M.; Kadri, N.K.; Crysnanto, D.; Pausch, H. Assessing genomic diversity and signatures of selection in Original Braunvieh cattle using whole-genome sequencing data. BMC Genom. 2020, 21, 27. [Google Scholar] [CrossRef] [Green Version]
- Stergiadis, S.; Bieber, A.; Franceschin, E.; Isensee, A.; Eyre, M.D.; Maurer, V.; Chatzidimitriou, E.; Cozzi, G.; Bapst, B.; Stewart, G.; et al. Impact of US Brown Swiss genetics on milk quality from low-input herds in Switzerland: Interactions with grazing intake and pasture type. Food Chem. 2015, 175, 609–618. [Google Scholar] [CrossRef]
- Blewett, H.J.; Gerdung, C.A.; Ruth, M.R.; Proctor, S.D.; Field, C.J. Vaccenic acid favourably alters immune function in obese JCR:LA-cp rats. Br. J. Nutr. 2009, 102, 526–536. [Google Scholar] [CrossRef] [Green Version]
- Jacome-Sosa, M.M.; Borthwick, F.; Mangat, R.; Uwiera, R.; Reaney, M.J.; Shen, J.; Quiroga, A.D.; Jacobs, R.L.; Lehner, R.; Proctor, S.D. Diets enriched in trans-11 vaccenic acid alleviate ectopic lipid accumulation in a rat model of NAFLD and metabolic syndrome. J. Nutr. Biochem. 2014, 25, 692–701. [Google Scholar] [CrossRef]
- Lim, J.N.; Oh, J.J.; Wang, T.; Lee, J.S.; Kim, S.H.; Kim, Y.J.; Lee, H.G. Trans-11 18:1 vaccenic acid (TVA) has a direct anti-carcinogenic effect on MCF-7 human mammary adenocarcinoma cells. Nutrients 2014, 6, 627–636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, J.; Wang, Y.; Fan, X.; Wu, H.; Han, J.; Yang, M.; Lu, L.; Nie, G. Trans-vaccenic acid inhibits proliferation and induces apoptosis of human nasopharyngeal carcinoma cells via a mitochondrial-mediated apoptosis pathway. Lipids Health Dis. 2019, 18, 46. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; England, A.; Sinclair, C.; Merkosky, F.; Chan, C.B. Trans-11 vaccenic acid improves glucose homeostasis in a model of type 2 diabetes by promoting insulin secretion via GPR40. J. Funct. Foods 2019, 60, 846–857. [Google Scholar] [CrossRef]
- Yang, B.; Chen, H.; Stanton, C.; Ross, R.P.; Zhang, H.; Chen, Y.Q.; Chen, W. Review of the roles of conjugated linoleic acid in health and disease. J. Funct. Foods 2015, 15, 314–325. [Google Scholar] [CrossRef]
- Simopoulos, A.P. The importance of the omega-6/omega-3 fatty acid ratio in cardiovascular disease and other chronic diseases. Exp. Biol. Med. 2008, 233, 674–688. [Google Scholar] [CrossRef]
- Coppa, M.; Gorlier, A.; Lonati, M.; Martin, B.; Russo, E.M.; Lombardi, G. The management of the transition from hayto pasture-based diets affects milk fatty acid kinetics. Dairy Sci. Technol. 2012, 92, 279–295. [Google Scholar] [CrossRef] [Green Version]
- Chilliard, Y.; Glasser, F.; Ferlay, A.; Bernard, L.; Rouel, J.; Doreau, M. Diet, rumen biohydrogenation and nutritional quality of cow and goat milk fat. Eur. J. Lipid Sci. Technol. 2007, 109, 828–855. [Google Scholar] [CrossRef]
- Kraft, J.; Collomb, M.; Möckel, P.; Sieber, R.; Jahreis, G. Differences in CLA isomer distribution of cow’s milk lipids. Lipids 2003, 38, 657–664. [Google Scholar] [CrossRef]
- Capuano, E.; Van Der Veer, G.; Boerrigter-Eenling, R.; Elgersma, A.; Rademaker, J.; Sterian, A.; Van Ruth, S.M. Verification of fresh grass feeding, pasture grazing and organic farming by cows farm milk fatty acid profile. Food Chem. 2014, 164, 234–241. [Google Scholar] [CrossRef]
- Coppa, M.; Martin, B.; Agabriel, C.; Chassaing, C.; Sibra, C.; Constant, I.; Graulet, B.; Andueza, D. Authentication of cow feeding and geographic origin on milk using visible and near-infrared spectroscopy. J. Dairy Sci. 2012, 95, 5544–5551. [Google Scholar] [CrossRef] [PubMed]
- Coppa, M.; Chassaing, C.; Ferlay, A.; Agabriel, C.; Laurent, C.; Borreani, G.; Barcarolo, R.; Baars, T.; Kusche, D.; Harstad, O.M.; et al. Potential of milk fatty acid composition to predict diet composition and authenticate feeding systems and altitude origin of European bulk milk. J. Dairy Sci. 2015, 98, 1539–1551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faustini, M.; Pastorino, G.Q.; Colombani, C.; Chiesa, L.M.; Panseri, S.; Vigo, D.; Curone, G. Volatilome in Milk for Grana Padano and Parmigiano Reggiano Cheeses: A first survey. Vet. Sci. 2019, 6, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Callaghan, T.F.; Hennessy, D.; McAuliffe, S.; Kilcawley, K.N.; O’Donovan, M.; Dillon, P.; Ross, R.P.; Stanton, C. Effect of pasture versus indoor feeding systems on raw milk composition and quality over an entire lactation. J. Dairy Sci. 2016, 99, 9424–9440. [Google Scholar] [CrossRef] [PubMed]
- Caredda, M.; Addis, M.; Ibba, I.; Leardi, R.; Scintu, M.F.; Piredda, G.; Sanna, G. Building of prediction models by using Mid-Infrared spectroscopy and fatty acid profile to discriminate the geographical origin of sheep milk. LWT Food Sci. Technol. 2017, 75, 131–136. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- International Organization for Standardization (ISO). Milk Fat-Preparation of Fatty Acid Methyl Esters; ISO Publications: Geneva, Switzerland, 2002; Volume ISO 15884|. [Google Scholar]
- Papaloukas, L.; Sinapis, E.; Arsenos, G.; Kyriakou, G.; Basdagianni, Z. Effect of season on fatty acid and terpene profiles of milk from Greek sheep raised under a semi-extensive production system. J. Dairy Res. 2016, 83, 375–382. [Google Scholar] [CrossRef]
- Garson, G.D. Factor Analysis; Statistical Associates Publishers: Asheboro, NC, USA, 2013. [Google Scholar]
- Righi, C.; Menchetti, L.; Orlandi, R.; Moscati, L.; Mancini, S.; Diverio, S. Welfare assessment in shelter dogs by using physiological and immunological parameters. Animals 2019, 9, 340. [Google Scholar] [CrossRef] [Green Version]
- Salque, M.; Bogucki, P.I.; Pyzel, J.; Sobkowiak-Tabaka, I.; Grygiel, R.; Szmyt, M.; Evershed, R.P. Earliest evidence for cheese making in the sixth millennium bc in northern Europe. Nature 2013, 493, 522–525. [Google Scholar] [CrossRef]
- Saroj, S.; Ganguly, S. Nutritional role of milk fatty acids to human health and its functional and biochemical properties. Asian J. Anim. Sci. 2015, 10, 220–225. [Google Scholar] [CrossRef]
- Garson, G.D. Discriminant Function Analysis; Statistical Associates Publishers: Asheboro, NC, USA, 2012. [Google Scholar]
- Pituch, K.A.; Stevens, J.P. Applied Multivariate Statistics for the Social Sciences, 6th ed.; Routledge: New York, NY, USA, 2016. [Google Scholar]
- Stergiadis, S.; Bieber, A.; Chatzidimitriou, E.; Franceschin, E.; Isensee, A.; Rempelos, L.; Baranski, M.; Maurer, V.; Cozzi, G.; Bapst, B.; et al. Impact of US Brown Swiss genetics on milk quality from low-input herds in Switzerland: Interactions with season. Food Chem. 2019, 251, 93–102. [Google Scholar] [CrossRef] [Green Version]
- Palmquist, D.L.; Davis, C.L.; Brown, R.E.; Sachan, D.S. Availability and Metabolism of Various Substrates in Ruminants. V. Entry Rate into the Body and Incorporation into Milk Fat of d(−)β-Hydroxybutyrate. J. Dairy Sci. 1969, 52, 633–638. [Google Scholar] [CrossRef]
- Parodi, P.W. Milk fat in human nutrition. Aust. J. Dairy Technol. 2004, 59, 3–59. [Google Scholar]
- Palmquist, D.L.; Mattos, W. Turnover of Lipoproteins and Transfer to Milk Fat of Dietary (1-Carbon-14) Linoleic Acid in Lactating Cows. J. Dairy Sci. 1978, 61, 561–565. [Google Scholar] [CrossRef]
- Prado, L.A.; Schmidely, P.; Nozière, P.; Ferlay, A. Milk saturated fatty acids, odd- and branched-chain fatty acids, and isomers of C18:1, C18:2, and C18:3n-3 according to their duodenal flows in dairy cows: A meta-analysis approach. J. Dairy Sci. 2019, 102, 3053–3070. [Google Scholar] [CrossRef] [Green Version]
- Clapham, W.M.; Foster, J.G.; Neel, J.P.S.; Fedders, J.M. Fatty acid composition of traditional and novel forages. J. Agric. Food Chem. 2005, 53, 10068–10073. [Google Scholar] [CrossRef]
- Jenkins, T.C. Lipid metabolism in the rumen. J. Dairy Sci. 1993, 76, 103–126. [Google Scholar] [CrossRef]
- Palmquist, D.L. Milk Fat: Origin of Fatty Acids and Influence of Nutritional Factors Thereon BT. In Advanced Dairy Chemistry Volume 2 Lipids; Springer: Boston, MA, USA, 2006; Volume 2, pp. 43–92. [Google Scholar] [CrossRef]
- Saini, R.K.; Keum, Y.S. Omega-3 and omega-6 polyunsaturated fatty acids: Dietary sources, metabolism, and significance—A review. Life Sci. 2018, 203, 255–267. [Google Scholar] [CrossRef]
- Glasser, F.; Doreau, M.; Maxin, G.; Baumont, R. Fat and fatty acid content and composition of forages: A meta-analysis. Anim. Feed Sci. Technol. 2013, 185, 19–34. [Google Scholar] [CrossRef]
- Dewhurst, R.J.; Shingfield, K.J.; Lee, M.R.F.; Scollan, N.D. Increasing the concentrations of beneficial polyunsaturated fatty acids in milk produced by dairy cows in high-forage systems. Anim. Feed Sci. Technol. 2006, 131, 168–206. [Google Scholar] [CrossRef]
- Shingfield, K.J.; Bonnet, M.; Scollan, N.D. Recent developments in altering the fatty acid composition of ruminant-derived foods. Animal 2013, 7, 132–162. [Google Scholar] [CrossRef] [PubMed]
- Vlaeminck, B.; Fievez, V.; Cabrita, A.R.J.; Fonseca, A.J.M.; Dewhurst, R.J. Factors affecting odd- and branched-chain fatty acids in milk: A review. Anim. Feed Sci. Technol. 2006, 131, 389–417. [Google Scholar] [CrossRef]
- Hurtaud, C.; Dutreuil, M.; Coppa, M.; Agabriel, C.; Martin, B. Characterization of milk from feeding systems based on herbage or corn silage with or without flaxseed and authentication through fatty acid profile. Dairy Sci. Technol. 2014, 94, 103–123. [Google Scholar] [CrossRef]
- Bonanno, A.; Tornambè, G.; Bellina, V.; De Pasquale, C.; Mazza, F.; Maniaci, G.; Di Grigoli, A. Effect of farming system and cheesemaking technology on the physicochemical characteristics, fatty acid profile, and sensory properties of Caciocavallo Palermitano cheese. J. Dairy Sci. 2013, 96, 710–724. [Google Scholar] [CrossRef]
- Falchero, L.; Lombardi, G.; Gorlier, A.; Lonati, M.; Odoardi, M.; Cavallero, A. Variation in fatty acid composition of milk and cheese from cows grazed on two alpine pastures. Dairy Sci. Technol. 2010, 90, 657–672. [Google Scholar] [CrossRef]
- Xu, T.; Tao, H.; Chang, G.; Zhang, K.; Xu, L.; Shen, X. Lipopolysaccharide derived from the rumen down-regulates stearoyl-CoA desaturase 1 expression and alters fatty acid composition in the liver of dairy cows fed a high-concentrate diet. BMC Vet. Res. 2015, 11, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, R.G. The composition of bovine milk lipids: January 1995 to December 2000. J. Dairy Sci. 2002, 85, 295–350. [Google Scholar] [CrossRef]
- Rymer, C.; Givens, D.I.; Wahle, K.W.J. Dietary strategies for increasing docosahexaenoic acid (DHA) and eicosapentaenoic acid (EPA) concentrations in bovine milk: A review. CAB Rev. 2003, 73, 9R–25R. [Google Scholar]
- La Terra, S.; Marino, V.M.; Manenti, M.; Licitra, G.; Carpino, S. Increasing pasture intakes enhances polyunsaturated fatty acids and lipophilic antioxidants in plasma and milk of dairy cows fed total mix ration. Dairy Sci. Technol. 2010, 90, 687–698. [Google Scholar] [CrossRef] [Green Version]
- Khanal, R.C.; Dhiman, T.R. Biosynthesis of Conjugated Linoleic Acid (CLA): A Review. Pak. J. Nutr. 2004, 3, 72–81. [Google Scholar] [CrossRef]
- Jenkins, T.C.; Martin, S.A. Factors affecting conjugated linoleic acid and trans-C18:1 fatty acid production by mixed ruminal bacteria. J. Anim. Sci. 2002, 80, 3347–3352. [Google Scholar] [CrossRef]
- Wyss, U. Influence of conservation method on fatty acid composition of herbages of a permanent grassland meadow. Grassl. Sci. Eur. 2012, 17, 323–325. [Google Scholar]
- Tempesta, T.; Vecchiato, D. An analysis of the territorial factors affecting milk purchase in Italy. Food Qual. Prefer. 2013, 27, 35–43. [Google Scholar] [CrossRef]
- Mosley, E.E.; McGuire, M.A. Methodology for the in vivo measurement of the Δ9- desaturation of myristic, palmitic, and stearic acids in lactating dairy cattle. Lipids 2007, 42, 939–945. [Google Scholar] [CrossRef] [PubMed]
- Mitani, T.; Kobayashi, K.; Ueda, K.; Kondo, S. Discrimination of “grazing milk” using milk fatty acid profile in the grassland dairy area in Hokkaido. Anim. Sci. J. 2016, 87, 233–241. [Google Scholar] [CrossRef] [Green Version]
- Gaspardo, B.; Lavrenčič, A.; Levart, A.; Del Zotto, S.; Stefanon, B. Use of milk fatty acids composition to discriminate area of origin of bulk milk. J. Dairy Sci. 2010, 93, 3417–3426. [Google Scholar] [CrossRef]
- Khanal, R.C.; Dhiman, T.R.; Boman, R.L. Changes in fatty acid composition of milk from lactating dairy cows during transition to and from pasture. Livest. Sci. 2008, 114, 164–175. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fatty Acids | Farm | Tot | ||||
|---|---|---|---|---|---|---|
| A | B | |||||
| Mean | SD | Mean | SD | Mean | SD | |
| Saturated fatty acids | ||||||
| C6:0 | 0.63 | 0.20 | 0.62 | 0.47 | 0.63 | 0.01 |
| C8:0 | 0.62 | 0.28 | 0.42 | 0.38 | 0.52 | 0.14 |
| C10:0 | 2.01 | 0.41 | 2.20 | 0.40 | 2.10 | 0.14 |
| C11:0 | 0.08 | 0.08 | 0.10 | 0.07 | 0.09 | 0.02 |
| C12:0 | 2.74 | 0.41 | 3.34 | 0.42 | 3.04 | 0.42 |
| C13:0 | 0.06 | 0.02 | 0.05 | 0.02 | 0.06 | 0.01 |
| C14:0 | 11.52 | 1.12 | 13.37 | 1.41 | 12.44 | 1.31 |
| C15:0 | 1.80 | 0.29 | 1.52 | 0.10 | 1.66 | 0.20 |
| C16:0 | 32.17 | 3.36 | 36.79 | 4.80 | 34.48 | 3.27 |
| C17:0 | 0.94 | 0.22 | 0.80 | 0.18 | 0.87 | 0.10 |
| C18:0 | 11.75 | 1.81 | 10.28 | 2.40 | 11.01 | 1.04 |
| C20:0 | 0.22 | 0.06 | 0.19 | 0.03 | 0.20 | 0.03 |
| C21:0 | 0.07 | 0.06 | 0.03 | 0.01 | 0.05 | 0.03 |
| C22:0 | 0.11 | 0.03 | 0.08 | 0.01 | 0.09 | 0.02 |
| C23:0 | 0.05 | 0.02 | 0.04 | 0.01 | 0.05 | 0.01 |
| C24:0 | 0.09 | 0.02 | 0.06 | 0.01 | 0.07 | 0.02 |
| Tot SFA | 64.85 | 4.04 | 69.89 | 3.94 | 67.37 | 3.57 |
| Monounsaturated fatty acids | ||||||
| C14:1 n9 | 0.95 | 0.17 | 1.10 | 0.25 | 1.02 | 0.11 |
| C16:1 n9 | 1.53 | 0.42 | 1.57 | 0.40 | 1.55 | 0.03 |
| C18:1 n6c | 1.01 | 1.59 | 1.73 | 2.72 | 1.37 | 0.51 |
| C18:1 n6t | 0.30 | 0.17 | 0.37 | 0.13 | 0.33 | 0.05 |
| C18:1 n9c | 23.17 | 2.25 | 19.53 | 3.33 | 21.35 | 2.57 |
| C18:1 n9t | 2.97 | 1.80 | 2.31 | 2.31 | 2.64 | 0.47 |
| C20:1 n9 | 0.18 | 0.05 | 0.10 | 0.05 | 0.14 | 0.06 |
| C22:1 n9 | 0.02 | 0.01 | 0.01 | 0.01 | 0.02 | 0.01 |
| C24:1 n9 | 0.02 | 0.01 | 0.01 | 0.00 | 0.02 | 0.01 |
| Tot MFA | 30.14 | 3.23 | 26.72 | 3.36 | 28.43 | 2.41 |
| n6 Polynsaturated fatty acids | ||||||
| C18:2 n6c | 1.76 | 0.41 | 1.55 | 0.30 | 1.66 | 0.15 |
| C18:2 n6t | 0.28 | 0.24 | 0.08 | 0.05 | 0.18 | 0.14 |
| C18:3 n6 | 0.01 | 0.00 | 0.01 | 0.00 | 0.01 | 0.01 |
| C20:2 n6 | 0.04 | 0.02 | 0.03 | 0.01 | 0.04 | 0.01 |
| C20:3 n6 | 0.06 | 0.01 | 0.06 | 0.01 | 0.06 | 0.00 |
| C20:4 n6 | 0.09 | 0.01 | 0.10 | 0.01 | 0.09 | 0.00 |
| C22:2 n6 | 0.05 | 0.03 | 0.03 | 0.01 | 0.04 | 0.01 |
| Tot n 6 PUFA | 2.29 | 0.37 | 1.85 | 0.31 | 2.07 | 0.31 |
| n3 Polynsaturated fatty acids | ||||||
| C18:3 n3 | 1.44 | 0.25 | 0.75 | 0.11 | 1.09 | 0.48 |
| C20:3 n3 | 0.02 | 0.01 | 0.02 | 0.01 | 0.02 | 0.01 |
| C20:5 n3 | 0.09 | 0.03 | 0.06 | 0.01 | 0.07 | 0.02 |
| C22:6 n3 | 0.01 | 0.00 | 0.01 | 0.00 | 0.01 | 0.00 |
| Tot n3 PUFA | 1.56 | 0.25 | 0.84 | 0.12 | 1.20 | 0.51 |
| Stereoisomers of conjugated linoleic acid | ||||||
| CLA n9c, n11t | 1.41 | 0.70 | 0.96 | 0.37 | 1.18 | 0.32 |
| n6/n3 ratio | 1.47 | 0.28 | 2.22 | 0.56 | 1.73 | 0.53 |
| Fatty Acids | Farm | Tot | ||||
|---|---|---|---|---|---|---|
| A | B | |||||
| Mean | SD | Mean | SD | Mean | SD | |
| Saturated fatty acids | ||||||
| C6:0 | 0.36 | 0.31 | 0.29 | 0.28 | 0.33 | 0.05 |
| C8:0 | 0.65 | 0.20 | 0.49 | 0.36 | 0.57 | 0.12 |
| C10:0 | 2.24 | 0.37 | 2.32 | 0.65 | 2.28 | 0.05 |
| C11:0 | 0.06 | 0.08 | 0.15 | 0.10 | 0.11 | 0.06 |
| C12:0 | 2.99 | 0.43 | 3.53 | 0.52 | 3.26 | 0.38 |
| C13:0 | 0.05 | 0.05 | 0.05 | 0.03 | 0.05 | 0.01 |
| C14:0 | 12.32 | 1.19 | 13.92 | 1.34 | 13.12 | 1.13 |
| C15:0 | 1.89 | 0.31 | 1.61 | 0.18 | 1.75 | 0.20 |
| C16:0 | 33.16 | 2.83 | 35.55 | 3.56 | 34.35 | 1.69 |
| C17:0 | 1.05 | 0.15 | 0.80 | 0.17 | 0.92 | 0.18 |
| C18:0 | 11.17 | 1.91 | 10.54 | 2.43 | 10.85 | 0.45 |
| C20:0 | 0.21 | 0.06 | 0.18 | 0.02 | 0.20 | 0.03 |
| C21:0 | 0.05 | 0.02 | 0.03 | 0.01 | 0.04 | 0.01 |
| C22:0 | 0.12 | 0.04 | 0.08 | 0.01 | 0.10 | 0.03 |
| C23:0 | 0.04 | 0.03 | 0.05 | 0.01 | 0.05 | 0.01 |
| C24:0 | 0.07 | 0.02 | 0.06 | 0.01 | 0.06 | 0.01 |
| Tot SFA | 66.44 | 3.16 | 69.62 | 2.99 | 68.03 | 2.25 |
| Monounsaturated fatty acids | ||||||
| C14:1 n9 | 1.03 | 0.14 | 1.10 | 0.25 | 1.07 | 0.06 |
| C16:1 n9 | 1.52 | 0.50 | 1.45 | 0.48 | 1.49 | 0.05 |
| C18:1 n6c | 1.89 | 2.91 | 1.44 | 2.08 | 1.67 | 0.32 |
| C18:1 n6t | 0.32 | 0.13 | 0.37 | 0.15 | 0.34 | 0.036 |
| C18:1 n9c | 21.75 | 2.85 | 20.44 | 3.29 | 21.10 | 0.92 |
| C18:1 n9t | 2.53 | 1.62 | 2.38 | 0.82 | 2.45 | 0.11 |
| C20:1 n9 | 0.10 | 0.08 | 0.10 | 0.05 | 0.10 | 0.01 |
| C22:1 n9 | 0.02 | 0.02 | 0.01 | 0.00 | 0.02 | 0.01 |
| C24:1 n9 | 0.03 | 0.03 | 0.01 | 0.00 | 0.02 | 0.01 |
| Tot MFA | 29.18 | 2.62 | 27.31 | 2.86 | 28.24 | 1.33 |
| n6 Polynsaturated fatty acids | ||||||
| C18:2 n6c | 1.94 | 0.32 | 1.61 | 0.24 | 1.78 | 0.23 |
| C18:2 n6t | 0.13 | 0.07 | 0.07 | 0.04 | 0.10 | 0.04 |
| C18:3 n6 | 0.02 | 0.01 | 0.01 | 0.00 | 0.01 | 0.01 |
| C20:2 n6 | 0.06 | 0.08 | 0.04 | 0.01 | 0.05 | 0.02 |
| C20:3 n6 | 0.07 | 0.05 | 0.06 | 0.02 | 0.06 | 0.01 |
| C20:4 n6 | 0.07 | 0.04 | 0.09 | 0.02 | 0.08 | 0.02 |
| C22:2 n6 | 0.03 | 0.02 | 0.03 | 0.01 | 0.03 | 0.01 |
| Tot n 6 PUFA | 2.32 | 0.37 | 1.91 | 0.26 | 2.12 | 0.29 |
| n3 Polynsaturated fatty acids | ||||||
| C18:3 n3 | 1.48 | 0.38 | 0.81 | 0.14 | 1.15 | 0.47 |
| C20:3 n3 | 0.02 | 0.00 | 0.02 | 0.00 | 0.02 | 0.01 |
| C20:5 n3 | 0.07 | 0.03 | 0.05 | 0.01 | 0.06 | 0.01 |
| C22:6 n3 | 0.05 | 0.07 | 0.01 | 0.00 | 0.03 | 0.03 |
| Tot n3 PUFA | 1.62 | 0.37 | 0.89 | 0.16 | 1.26 | 0.52 |
| Stereoisomers of conjugated linoleic acid | ||||||
| CLA n9c, n11t | 0.73 | 0.37 | 0.42 | 0.09 | 0.58 | 0.22 |
| n6/n3 ratio | 1.43 | 0.27 | 2.15 | 0.54 | 1.68 | 0.51 |
| Item | Component | ||
|---|---|---|---|
| PC1. C < 18 − C18 | PC2. C18 PUFA & C15 | PC3. C > 18 & CLA | |
| C18:0 | 0.947 | −0.025 | −0.047 |
| C16:0 | −0.893 | −0.235 | −0.208 |
| C14:0 | −0.861 | −0.274 | −0.281 |
| C14:1 n9 | −0.853 | 0.081 | −0.103 |
| C18:1 n9c | 0.725 | 0.139 | 0.147 |
| C18:3 n3 | 0.222 | 0.882 | 0.294 |
| C15:0 | −0.123 | 0.873 | 0.110 |
| C18:2 n6c | 0.474 | 0.609 | 0.011 |
| C21:0 | 0.174 | 0.101 | 0.870 |
| C20:5 n3 | −0.062 | 0.197 | 0.791 |
| CLA n9c, n11t | 0.423 | 0.062 | 0.674 |
| % Variance explained | 45.7 | 18.8 | 11.9 |
| Cumulative % variance explained | 76.4 | ||
| Product | Variable | Standardized Canonical Discriminant Function Coefficients | Wilks’ Lambda | p Value |
|---|---|---|---|---|
| Milk | PC2.C18 PUFA & C15 | 0.901 | 0.567 | <0.001 |
| PC3.C > 18 & CLA | 0.766 | 0.747 | 0.012 | |
| PC1. C < 18 − C18 | 0.396 | 0.867 | 0.079 | |
| Cheese | PC2.C18 PUFA & C15 | 0.988 | 0.618 | 0.001 |
| PC3.C > 18 & CLA | 0.780 | 0.843 | 0.055 | |
| PC1. C < 18 − C18 | 0.766 | 0.961 | 0.355 |
| Product | Farm | Predicted Group Membership | Total | |
|---|---|---|---|---|
| Grazing System | No Grazing System | |||
| Milk 1 | Grazing system | 9 (75.0) | 3 (25.0) | 12 |
| No grazing system | 1 (8.3) | 11 (91.7) | 12 | |
| Cheese 2 | Grazing system | 9 (75.0) | 3 (25.0) | 12 |
| No grazing system | 0 (0.0) | 12 (100.0) | 12 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Agradi, S.; Curone, G.; Negroni, D.; Vigo, D.; Brecchia, G.; Bronzo, V.; Panseri, S.; Chiesa, L.M.; Peric, T.; Danes, D.; et al. Determination of Fatty Acids Profile in Original Brown Cows Dairy Products and Relationship with Alpine Pasture Farming System. Animals 2020, 10, 1231. https://doi.org/10.3390/ani10071231
Agradi S, Curone G, Negroni D, Vigo D, Brecchia G, Bronzo V, Panseri S, Chiesa LM, Peric T, Danes D, et al. Determination of Fatty Acids Profile in Original Brown Cows Dairy Products and Relationship with Alpine Pasture Farming System. Animals. 2020; 10(7):1231. https://doi.org/10.3390/ani10071231
Chicago/Turabian StyleAgradi, Stella, Giulio Curone, Daniele Negroni, Daniele Vigo, Gabriele Brecchia, Valerio Bronzo, Sara Panseri, Luca Maria Chiesa, Tanja Peric, Doina Danes, and et al. 2020. "Determination of Fatty Acids Profile in Original Brown Cows Dairy Products and Relationship with Alpine Pasture Farming System" Animals 10, no. 7: 1231. https://doi.org/10.3390/ani10071231