Perch Positioning Affects both Laying Hen Locomotion and Forces Experienced at the Keel


Abstract
:Simple Summary
Abstract
1. Introduction
2. Animals, Material and Methods
2.1. Ethical Approval
2.2. Animals and Housing
2.3. Rewarding Apparatus
2.4. Habituation to the Test Pen and Selection of Focal Hens
2.5. Training
2.6. Experimental Design and Test Conditions
2.7. Data Collection
2.7.1. Video Recording
2.7.2. Acceleration Sensors and Calculations
2.8. Statistical Analysis
3. Results
3.1. Peak Force at the Keel
3.1.1. Upward Transitions
3.1.2. Downward Transition
3.2. Impulse
3.2.1. Upward Transition
3.2.2. Downward Transition
3.3. Latency to Transition
3.3.1. Upward Transition
3.3.2. Downward Transition
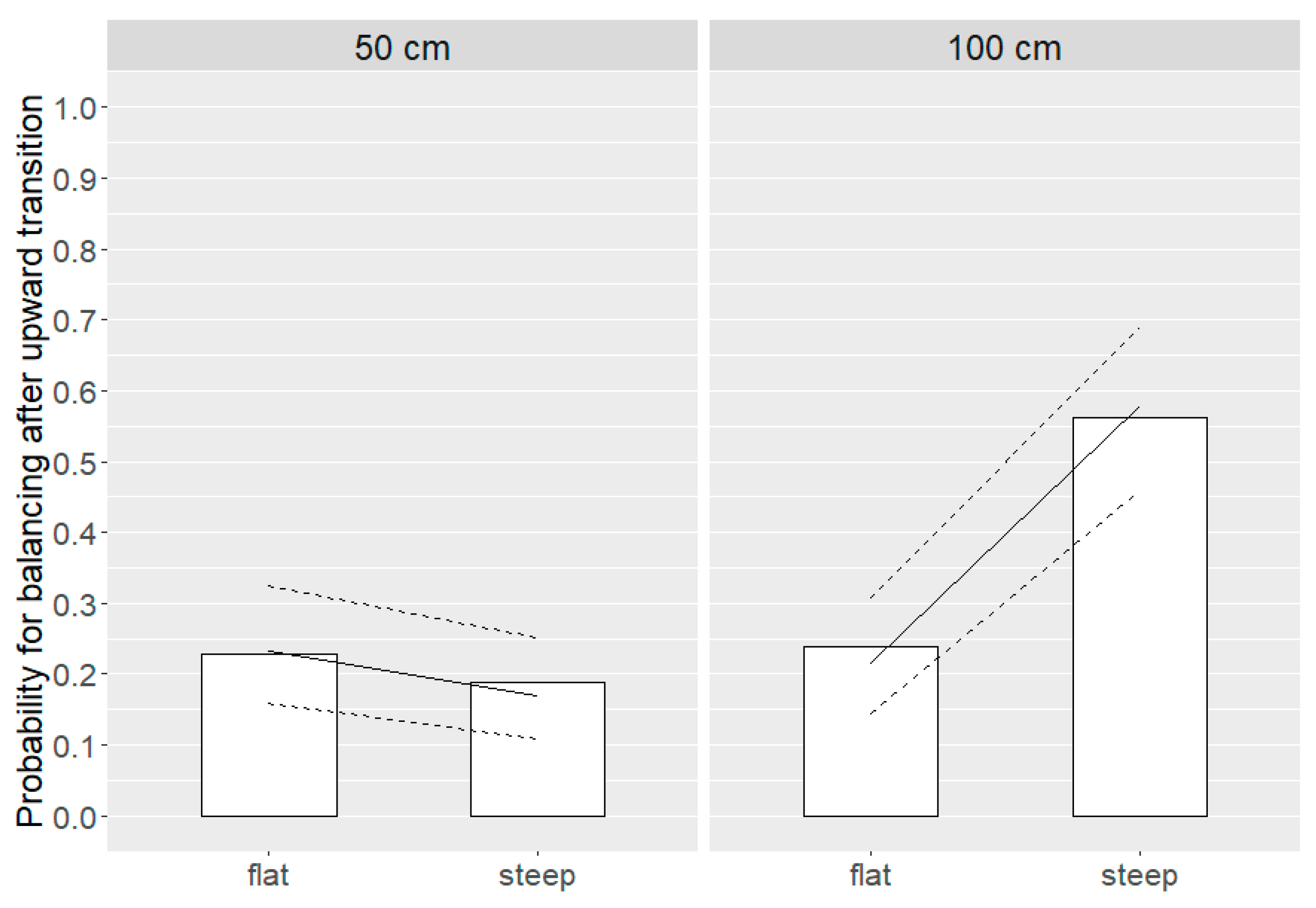
3.4. Balancing Movements at Landing
3.4.1. Upward Transition
3.4.2. Downward Transition
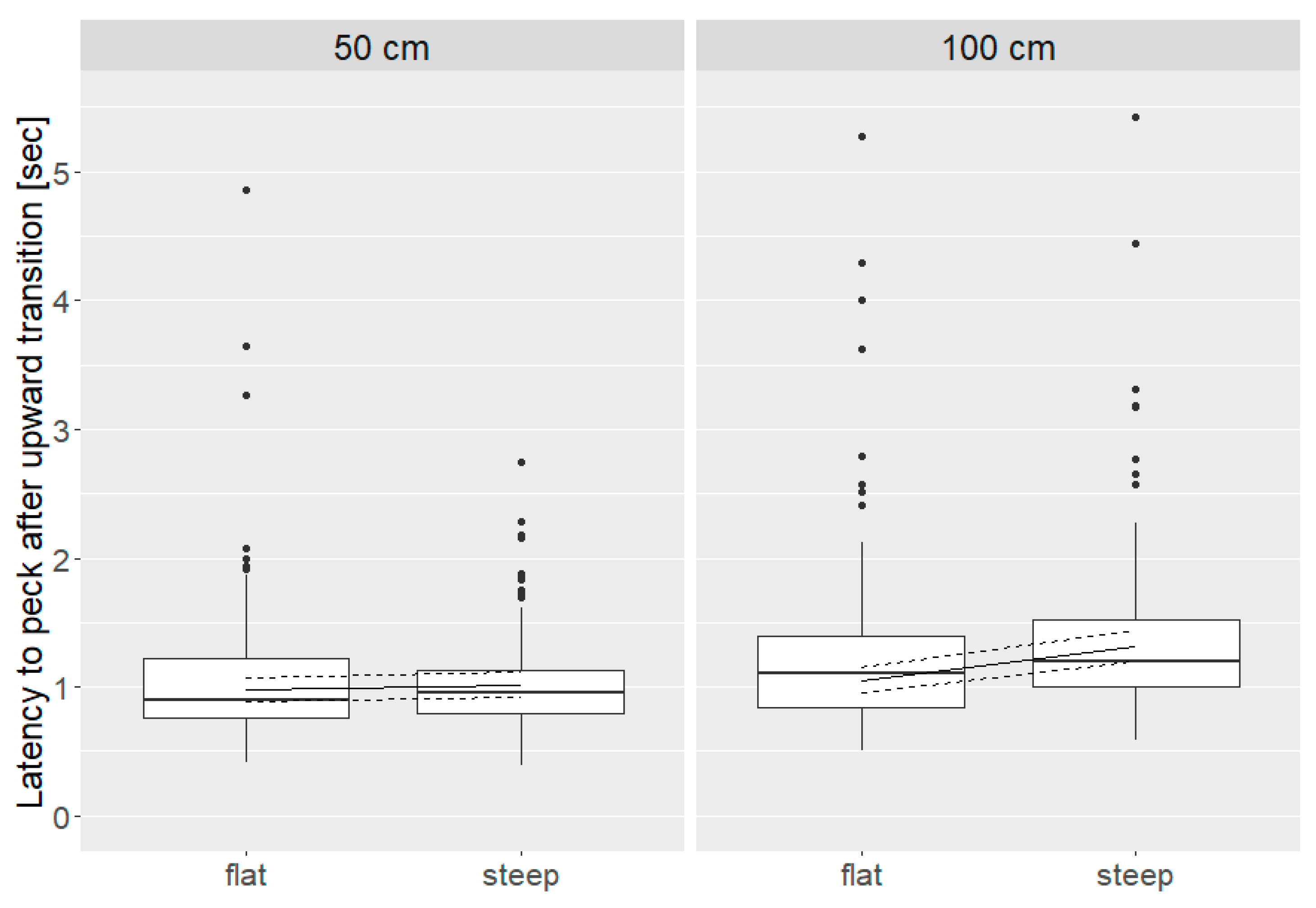
3.5. Latency to First Peck
3.5.1. Upward Transition
3.5.2. Downward Transition
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A—Habituation and Selection Criteria
Appendix B—Training Procedure
Appendix B.1. Stage 1: Feeding from Platform (20 Weeks of Age)
Appendix B.2. Stage 2: Feeding from Perch (21 Weeks of Age)
Appendix B.3. Stage 3: Step from Platform to Perch (22 Weeks of Age)
Appendix B.4. Stage 4: Horizontal Distance (23 Weeks of Age)
Appendix B.5. Stage 5: Angles upwards (24 Weeks of Age)
Appendix B.6. Stage 6: Angles downwards (25 Weeks of Age)
Appendix B.7. Stage 7: Varying Conditions (26 Weeks of Age)
Appendix B.7.1. General Notes
Appendix B.7.2. Determining Time Stamps
References
- Karcher, D.M.; Mench, J.A. Overview of commercial poultry production systems and their main welfare challenges. In Advances in Poultry Welfare Science; Mench, J.A., Ed.; Woodhead Publishing: Camridge, UK, 2018; pp. 3–25. [Google Scholar]
- Cooper, J.J.; Albentosa, M.J. Behavioural Priorities of Laying Hens. Avian Poult. Biol. Rev. 2003, 14, 127–149. [Google Scholar] [CrossRef]
- Weeks, C.A.; Nicol, C.J. Behavioural needs, priorities and preferences of laying hens. World’s Poult. Sci. J. 2006, 62, 296–307. [Google Scholar] [CrossRef]
- Wilkins, L.J.; McKinstry, J.L.; Avery, N.C.; Knowles, T.G.; Brown, S.N.; Tarlton, J.; Nicol, C.J. Influence of housing system and design on bone strength and keel bone fractures in laying hens. Vet. Rec. 2011, 169, 414. [Google Scholar] [CrossRef]
- Rufener, C.; Makagon, M.M. Keel bone fractures in laying hens: A systematic review of prevalence across age, housing systems, and strains. J. Anim. Sci. 2020, in press. [Google Scholar]
- Harlander-Matauschek, A.; Rodenburg, T.B.; Sandilands, V.; Tobalske, B.W.; Toscano, M.J. Causes of keel bone damage and their solutions in laying hens. World’s Poult. Sci. J. 2015, 71, 461–472. [Google Scholar] [CrossRef] [Green Version]
- Moinard, C.; Statham, P.; Haskell, M.J.; McCorquodale, C.; Jones, R.B.; Green, P.R. Accuracy of laying hens in jumping upwards and downwards between perches in different light environments. Appl. Anim. Behav. Sci. 2004, 85, 77–92. [Google Scholar] [CrossRef]
- Scott, G.B.; Lambe, N.R.; Hitchcock, D. Ability of laying hens to negotiate horizontal perches at different heights, separated by different angles. Br. Poult. Sci. 1997, 38, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Lambe, N.R.; Scott, G.B.; Hitchcock, D. Behaviour of Laying Hens Negotiating Perches at Different Heights. Anim. Welf. 1997, 6, 29–41. [Google Scholar]
- Scholz, B.; Kjaer, J.B.; Schrader, L. Analysis of landing behaviour of three layer lines on different perch designs. Br. Poult. Sci. 2014, 55, 419–426. [Google Scholar] [CrossRef]
- Scott, G.B.; Parker, C.A.L. The ability of laying hens to negotiate between horizontal perches. Appl. Anim. Behav. Sci. 1994, 42, 121–127. [Google Scholar] [CrossRef]
- Scott, G.B.; Hughes, B.O.; Lambe, N.R.; Waddington, D. Ability of laying hens to jump between perches: Individual variation and the effects of perch separation and motivation on behaviour. Br. Poult. Sci. 1999, 40, 177–184. [Google Scholar] [CrossRef]
- Stratmann, A.; Fröhlich, E.K.F.; Gebhardt-Henrich, S.G.; Harlander-Matauschek, A.; Würbel, H.; Toscano, M.J. Modification of aviary design reduces incidence of falls, collisions and keel bone damage in laying hens. Appl. Anim. Behav. Sci. 2015, 165, 112–123. [Google Scholar] [CrossRef]
- Thøfner, I.; Hougen, H.P.; Villa, C.; Lynnerup, N.; Christensen, J.P. Pathological characterization of keel bone fractures in laying hens does not support external trauma as the underlying cause. PLoS ONE 2020, 15, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, M.W.; Greenspan, A. Stress Fractures. Radiology 1996, 199, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Knapp, T.P.; Garrett, W.E. Stress Fractures: General Concepts. Clin. Sports Med. 1997, 16, 339–356. [Google Scholar] [CrossRef]
- Banerjee, D.; Daigle, C.L.; Dong, B.; Wurtz, K.; Newberry, R.C.; Siegford, J.M.; Biswas, S. Detection of jumping and landing force in laying hens using wireless wearable sensors. Poult. Sci. 2014, 93, 2724–2733. [Google Scholar] [CrossRef]
- Baker, S.L.; Robison, C.I.; Karcher, D.M.; Toscano, M.J.; Makagon, M.M. Keel impacts and associated behaviors in laying hens. Appl. Anim. Behav. Sci. 2020, 222, 104886. [Google Scholar] [CrossRef]
- Rentsch, A.K.; Rufener, C.B.; Spadavecchia, C.; Stratmann, A.; Toscano, M.J. Laying Hens’ Mobility Is Impaired by Keel Bone Fractures Though Effect Is Not Reversed by Paracetamol Treatment. Appl. Anim. Behav. Sci. 2019. [Google Scholar] [CrossRef]
- Casey-Trott, T.; Heerkens, J.L.T.; Petrik, M.; Regmi, P.; Schrader, L.; Toscano, M.J.; Widowski, T. Methods for assessment of keel bone damage in poultry. Poult. Sci. 2015, 94, 2339–2350. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017; Available online: http://www.r-project (accessed on 27 May 2020).
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Hartig, F. DHARMa: Residual Diagnostics for Hierarchical (Multi-Level/Mixed) Regression Models. R Package Version 0.1.6. 2018. Available online: https://cran.r-project.org/package=DHARMa (accessed on 27 May 2020).
- Halekoh, U.; Højsgaard, S. A Kenward-Roger Approximation and Parametric Bootstrap Methods for Tests in Linear Mixed Models—The R Package pbkrtest. J. Stat. Softw. 2014, 59, 1–30. [Google Scholar] [CrossRef] [Green Version]
- Fox, J. Effect Displays in R for Generalised Linear Models. J. Stat. Softw. 2003, 8, 1–27. [Google Scholar] [CrossRef] [Green Version]
- Moinard, C.; Statham, P.; Green, P.R. Control of landing flight by laying hens: Implications for the design of extensive housing systems. Br. Poult. Sci. 2004, 45, 578–584. [Google Scholar] [CrossRef] [PubMed]
- Green, P.; Moinard, C.; Statham, P. Control of landing flight in laying hens: What are the effects of domestication? In Welfare of the Laying Hen; Perry, G.C., Ed.; CABI Publishing: Wallingford, UK, 2004; p. 403. [Google Scholar]
- Pickel, T.; Scholz, B.; Schrader, L. Perch material and diameter affects particular perching behaviours in laying hens. Appl. Anim. Behav. Sci. 2010, 127, 37–42. [Google Scholar] [CrossRef]
- Mackie, N.; Stratmann, A.; Toscano, M.J.; Tarlton, J.F. Using tri-axial accelerometers to determine hazardous pathways for laying hens in aviaries. In Proceedings of the Xth European Symposium on Poultry Welfare, Ploufragan, France, 19–22 June 2017; Volume 97. [Google Scholar]
- Candelotto, L.; Stratmann, A.; Gebhardt-Henrich, S.G.; Rufener, C.; Van De Braak, T.; Toscano, M.J. Susceptibility to keel bone fractures in laying hens and the role of genetic variation. Poult. Sci. 2017, 96. [Google Scholar] [CrossRef] [PubMed]
- Toscano, M.J.; Wilkins, L.J.; Millburn, G.; Thorpe, K.; Tarlton, J.F. Development of an Ex Vivo Protocol to Model Bone Fracture in Laying Hens Resulting from Collisions. PLoS ONE 2013, 8, e66215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toscano, M.J.; Booth, F.; Richards, G.; Brown, S.N.; Karcher, D.M.; Tarlton, J.F. Modeling collisions in laying hens as a tool to identify causative factors for keel bone fractures and means to reduce their occurrence and severity. PLoS ONE 2018, 13, e0200025. [Google Scholar] [CrossRef] [Green Version]
- Toscano, M.J.; Dunn, I.C.; Christensen, J.-P.; Petow, S.; Kittelsen, K.; Ulrich, R. Explanations for keel bone fractures in laying hens: Are there explanations in addition to elevated egg production? Poult. Sci. 2020. [Google Scholar] [CrossRef]
- Casey-Trott, T.M.; Guerin, M.T.; Sandilands, V.; Torrey, S.; Widowski, T.M. Rearing system affects prevalence of keel-bone damage in laying hens: A longitudinal study of four consecutive flocks. Poult. Sci. 2017, 96, 2029–2039. [Google Scholar] [CrossRef]
- Scholz, B.; Rönchen, S.; Hamann, H.; Hewicker-Trautwein, M.; Distl, O. Keel bone condition in laying hens: A histological evaluation of macroscopically assessed keel bones. Berl. Munch. Tierarztl. Wochenschr. 2008, 121, 89–94. [Google Scholar]
- Sisson, S.; Grossman, J.D.; Getty, R. Sisson and Grossman’s The Anatomy of the Domestic Animals, 5th ed.; Getty, R., Ed.; Saunders: Philadelphia, PA, USA, 1975. [Google Scholar]
- Eusemann, B.K.; Patt, A.; Schrader, L.; Weigend, S.; Thöne-Reineke, C.; Petow, S. The Role of Egg Production in the Etiology of Keel Bone Damage in Laying Hens. Front. Vet. Sci. 2020, 7. [Google Scholar] [CrossRef] [Green Version]
- Stratmann, A.; Fröhlich, E.K.F.; Gebhardt-Henrich, S.G.; Harlander-Matauschek, A.; Würbel, H.; Toscano, M.J. Genetic selection to increase bone strength affects prevalence of keel bone damage and egg parameters in commercially housed laying hens. Poult. Sci. 2016, 95, 975–984. [Google Scholar] [CrossRef]
- Rufener, C.; Abreu, Y.; Asher, L.; Berezowski, J.; Maximiano Sousa, F.; Stratmann, A.; Toscano, M.J. Keel bone fractures are associated with individual mobiltiy of laying hen in aviary systems. Appl. Anim. Behav. Sci. 2019, 217, 48–56. [Google Scholar] [CrossRef]
- Rentsch, A.K.; Rufener, C.; Spadavecchia, C.; Stratmann, A.; Toscano, M.J. Effect of keel bone fractures on laying hen behaviour in a noncage housing system. In Proceedings of the XVth European Poultry Conference; Prukner-Radovčić, E., Medić, H., Eds.; World’s Poultry Science Journal: Dubrovnik, Croatia, 2018; p. 229. [Google Scholar]
- Nasr, M.A.F.; Murrell, J.; Wilkins, L.J.; Nicol, C.J. The effect of keel fractures on egg production parameters, mobility and behaviour in individual laying hens. Anim. Welf. 2012, 21, 127–135. [Google Scholar] [CrossRef]
- Stratmann, A.; Ringgenberg, N. Assessment of aviary systems for laying hens in Switzerland. In KTBL-Schrift 514. Aktuelle Arbeiten zur artgemäßen Tierhaltung 2018; Kuratorium für Technik und Bauwesen in der Landwirtschaft e.V. (KTBL): Darmstadt, Germany, 2018; pp. 137–147. ISBN 978-3-945088-60-9. [Google Scholar]
- Janczak, A.M.; Riber, A.B. Review of rearing-related factors affecting the welfare of laying hens. Poult. Sci. 2015, 94, 1454–1469. [Google Scholar] [CrossRef] [PubMed]
- Heerkens, J.L.T.; Delezie, E.; Rodenburg, T.B.; Kempen, I.; Zoons, J.; Ampe, B.; Tuyttens, F.A.M. Risk factors associated with keel bone and foot pad disorders in laying hens housed in aviary systems: Table 1. Poult. Sci. 2016, 95, 482–488. [Google Scholar] [CrossRef]
- Stratmann, A.; Mühlemann, S.; Vögeli, S.; Ringgenberg, N. Frequency of falls in commercial aviary-housed laying hen flocks and the effects of dusk phase length. Appl. Anim. Behav. Sci. 2019, 216, 26–32. [Google Scholar] [CrossRef]
- Moinard, C.; Rutherford, K.M.D.; Haskell, M.J.; McCorquodale, C.; Jones, R.B.; Green, P.R. Effects of obstructed take-off and landing perches on the flight accuracy of laying hens. Appl. Anim. Behav. Sci. 2005, 93, 81–95. [Google Scholar] [CrossRef]
- Taylor, P.E.; Scott, G.B.; Rose, P. The ability of domestic hens to jump between horizontal perches: Effects of light intensity and perch colour. Appl. Anim. Behav. Sci. 2003, 83, 99–108. [Google Scholar] [CrossRef]
- Heerkens, J.L.T.; Delezie, E.; Ampe, B.; Rodenburg, T.B.; Tuyttens, F.A.M. Ramps and hybrid effects on keel bone and foot pad disorders in modified aviaries for laying hens. Poult. Sci. 2016, 95, 2479–2488. [Google Scholar] [CrossRef]
- MacLachlan, S.; Ali, A.; Stratmann, A.; Toscano, M.; Siegford, J. Prevalence of keel bone damage in laying hens can be influenced by using ramps in pullet rearing and laying hen aviaries. Poultry Sci. 2018. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Description | |
|---|---|---|
| Signal | hh:mm:ss.000 | LED on |
| Take-off start | hh:mm:ss.000 | first forward/upward movement (upward jumps) or forward/downward movement (downward jumps) |
| Take-off end | hh:mm:ss.000 | both feet (and toes) lose contact to take-off platform |
| Landing start | hh:mm:ss.000 | first contact with the landing perch |
| Landing end | hh:mm:ss.000 | both feet on the perch and body of the hen stabilized with both wings folded close to the body, head pointing forward, and tail in a horizontal position |
| First peck | hh:mm:ss.000 | first peck directed towards the reward indicated by head reaching the lowest position and neck feathers erecting |
| Outcome Variable | Effects 1 | p-Value | Pattern | ||
|---|---|---|---|---|---|
| Peak force | Upward | Take-off | Distance | 0.027 | 100 cm > 50 cm |
| Flight | Distance * angle | 0.037 | 50 cm: flat > steep 100 cm: flat = steep | ||
| Landing | Distance | 0.019 | 100 cm > 50 cm | ||
| Downward | Take-off | Hybrid | 0.032 | Brown Nick > Nick Chick | |
| Flight | Hybrid | 0.008 | Brown Nick > Nick Chick | ||
| Distance * angle | 0.038 | 50 cm: flat = steep 100 cm: steep > flat | |||
| Landing | Distance | 0.019 | 100 cm > 50 cm | ||
| Hybrid * angle | 0.026 | Nick Chick: flat = steep Brown Nick: steep > flat Brown Nick > Nick Chick | |||
| Impulse | Upward | Take-off | Distance * angle | 0.036 | 50 cm: flat > steep 100 cm: flat < steep |
| Landing | Distance * angle | 0.001 | 50 cm: flat = steep 100 cm: steep > flat 100 cm > 50 cm | ||
| Downward | Take-off | Hybrid | 0.018 | Brown Nick > Nick Chick | |
| Distance * angle | 0.004 | 50 cm: flat = steep 100 cm: steep > flat | |||
| Landing | Distance | 0.001 | 100 cm > 50 cm | ||
| Latency to transition | Upward | Hybrid | 0.016 | Brown Nick > Nick Chick | |
| Distance * angle | 0.048 | 50 cm: flat = steep 100 cm: steep > flat 100 cm > 50 cm | |||
| Downward | Distance | 0.001 | 100 cm > 50 cm | ||
| Angle | 0.001 | Steep > flat | |||
| Balancing movements | Upward | Distance * angle | 0.001 | 50 cm: flat = steep 100 cm: steep > flat | |
| Downward | Distance | 0.0001 | 100 cm > 50 cm | ||
| Latency to first peck | Upward | Distance * angle | 0.001 | 50 cm: flat = steep 100 cm: steep > flat | |
| Downward | Hybrid | 0.034 | Nick Chick > Brown Nick | ||
| Distance * angle | 0.006 | 50 cm: flat = steep 100 cm: steep > flat 100 cm > 50 cm | |||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rufener, C.; Rentsch, A.K.; Stratmann, A.; Toscano, M.J. Perch Positioning Affects both Laying Hen Locomotion and Forces Experienced at the Keel. Animals 2020, 10, 1223. https://doi.org/10.3390/ani10071223
Rufener C, Rentsch AK, Stratmann A, Toscano MJ. Perch Positioning Affects both Laying Hen Locomotion and Forces Experienced at the Keel. Animals. 2020; 10(7):1223. https://doi.org/10.3390/ani10071223
Chicago/Turabian StyleRufener, Christina, Ana K. Rentsch, Ariane Stratmann, and Michael J. Toscano. 2020. "Perch Positioning Affects both Laying Hen Locomotion and Forces Experienced at the Keel" Animals 10, no. 7: 1223. https://doi.org/10.3390/ani10071223