High Frequency of Intravenous Injection of Human Adipose Stem Cell Conditioned Medium Improved Embryo Development of Mice in Advanced Maternal Age through Antioxidant Effects

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Approval
2.2. Chemicals
2.3. Isolation of Human Adipose Stem Cell and Preparation of Conditioned Medium
2.4. Animals and Treatments
2.5. Oocyte Collection
2.6. Parthenogenetic Activation and Evaluation of Parthenote Development
2.7. Assessment of Natural Mating and Implantation
2.8. Gene Analysis by Quantitative Real-Time Polymerase Chain Reaction
2.9. Statistical Analysis
3. Results
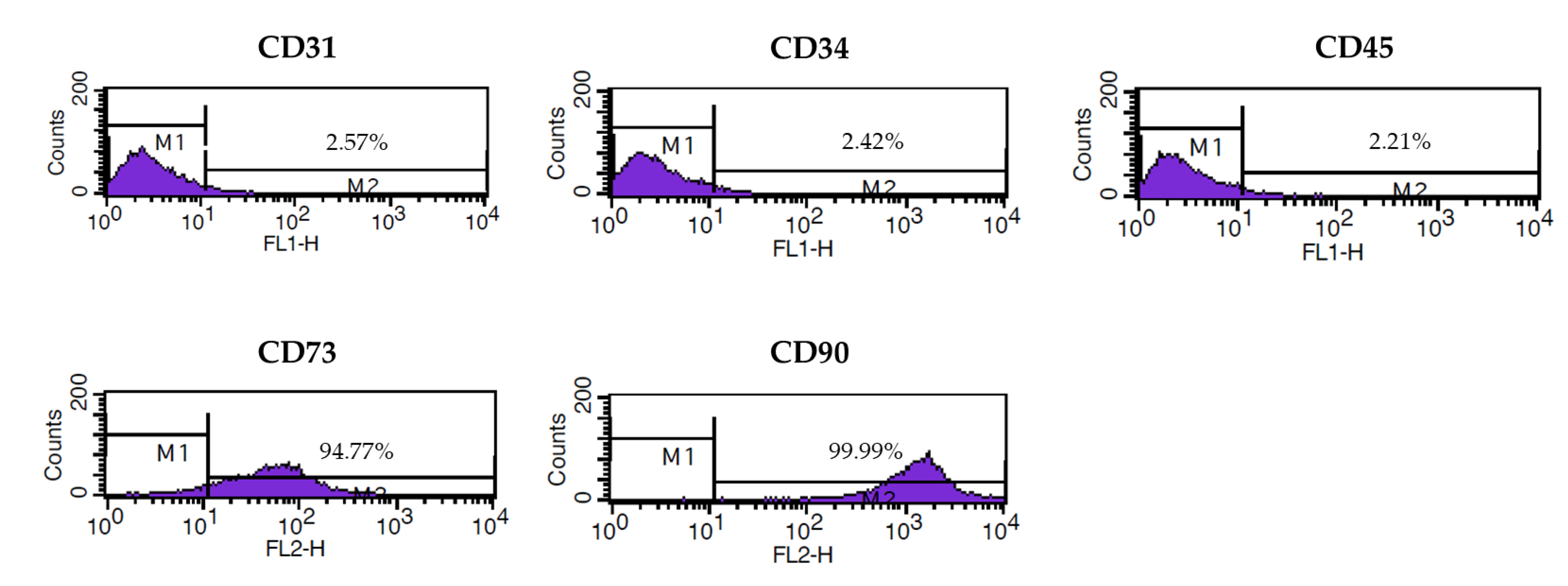
3.1. Characterization of Isolated ASCs
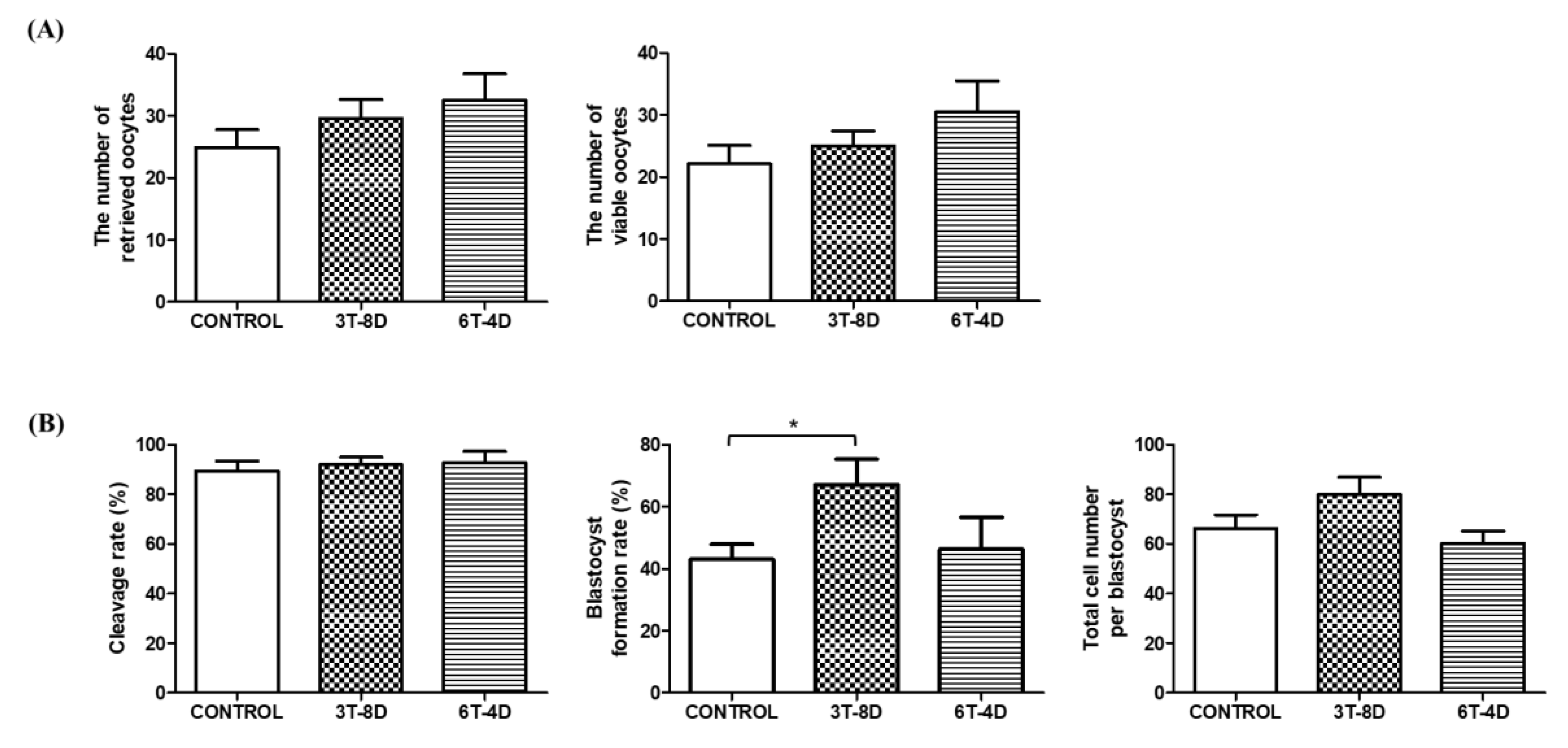
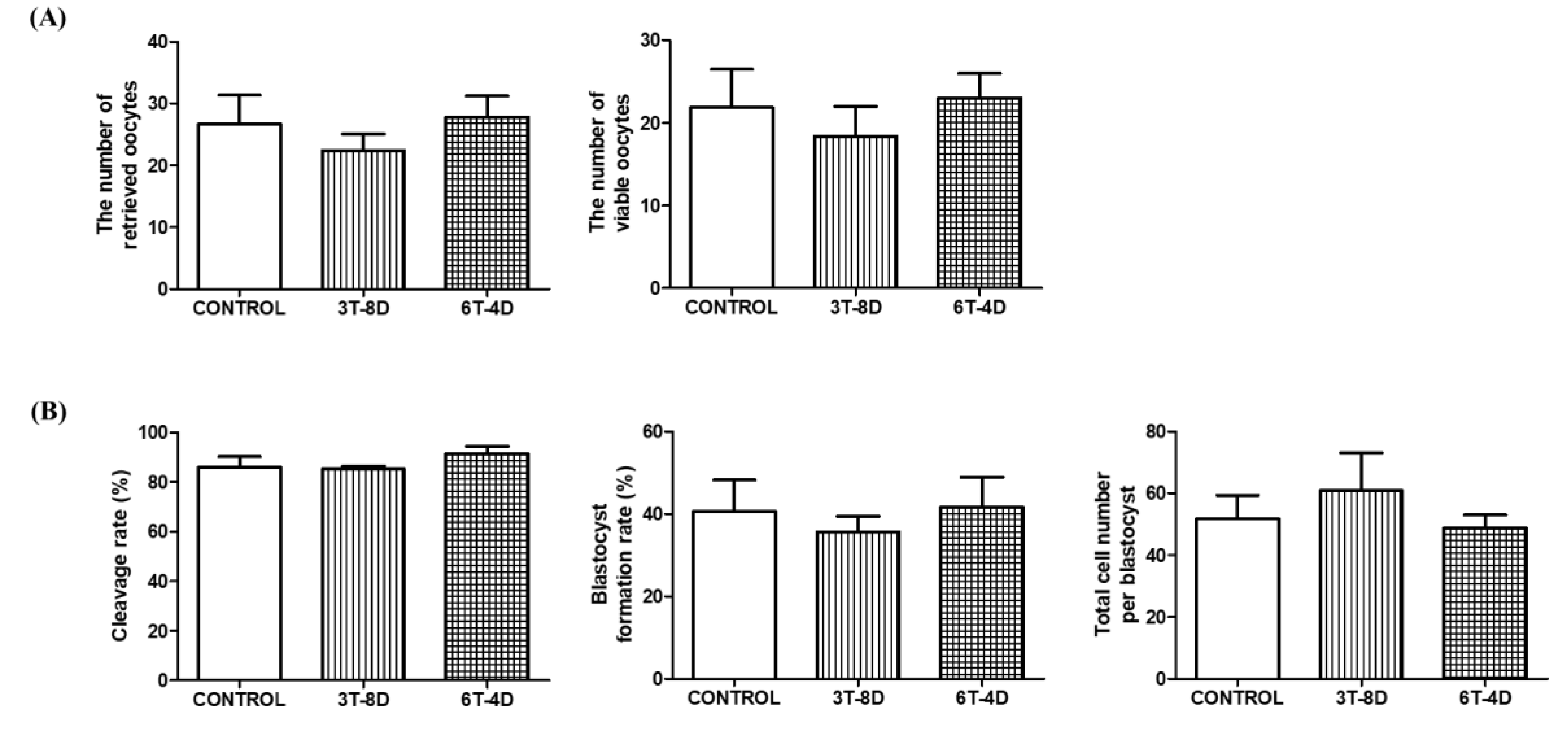
3.2. Ovulation and in Vitro Development of Parthenotes from 4- and 6-Month-Old Mice
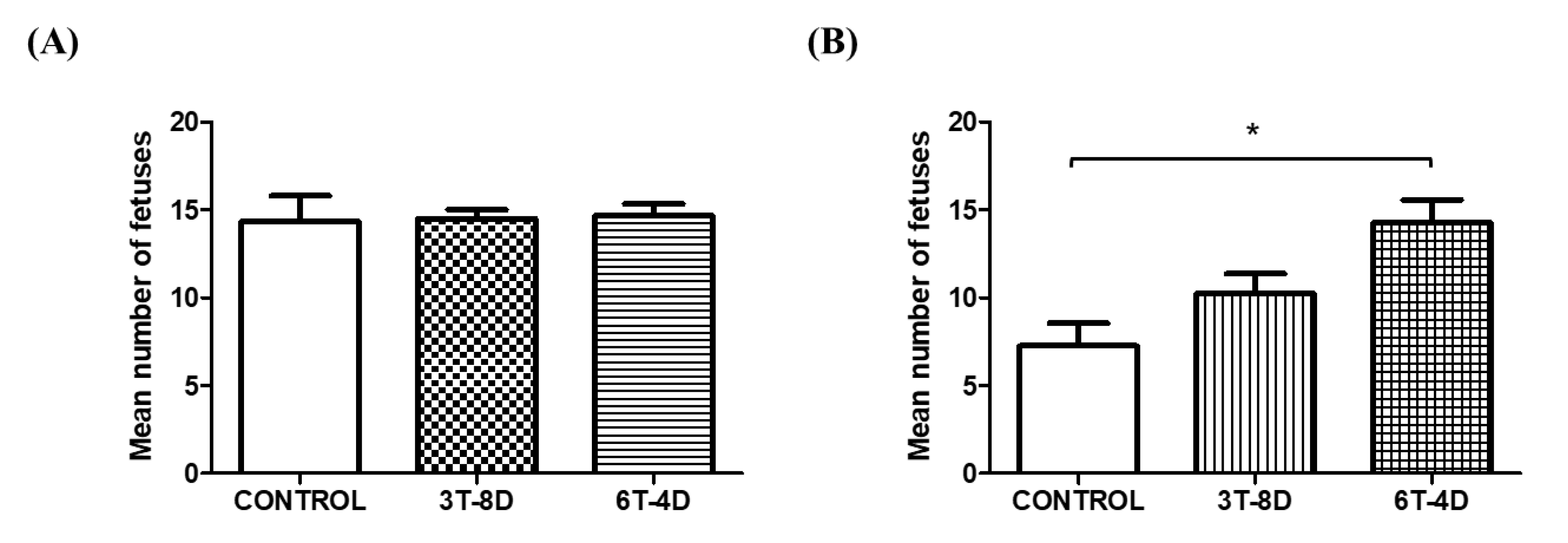
3.3. Implanted Fetuses of 4- and 6-Month-Old Mice
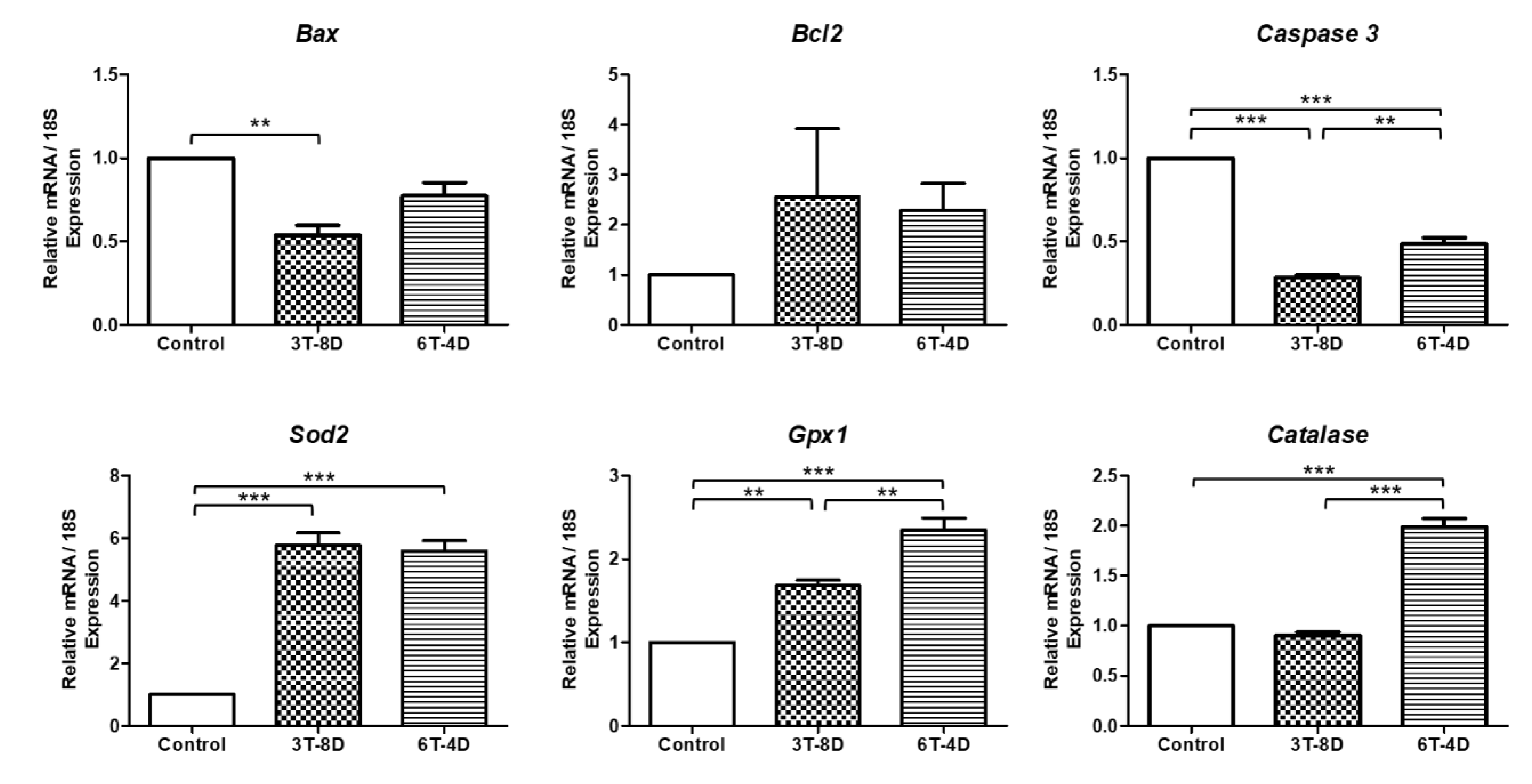
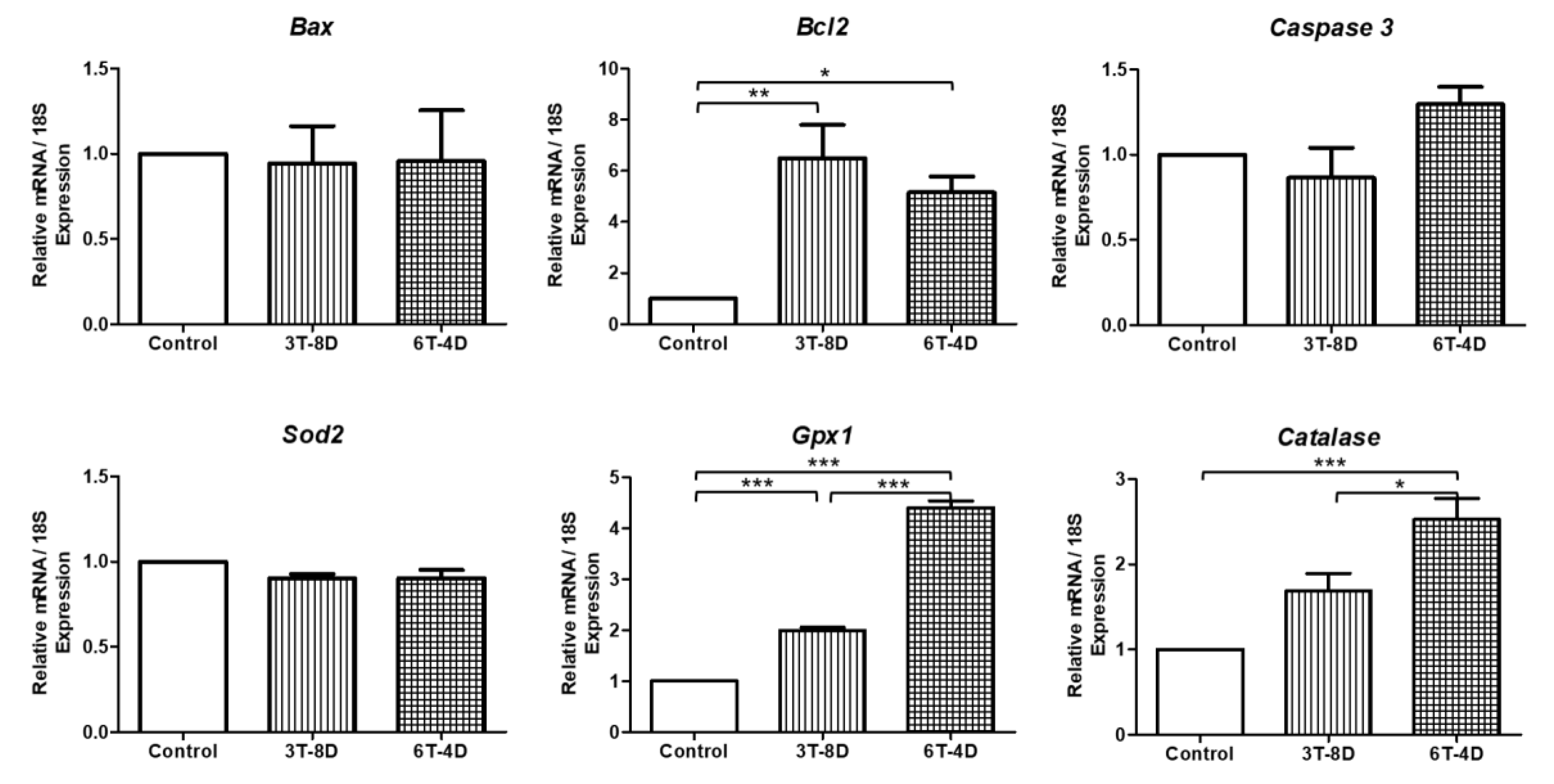
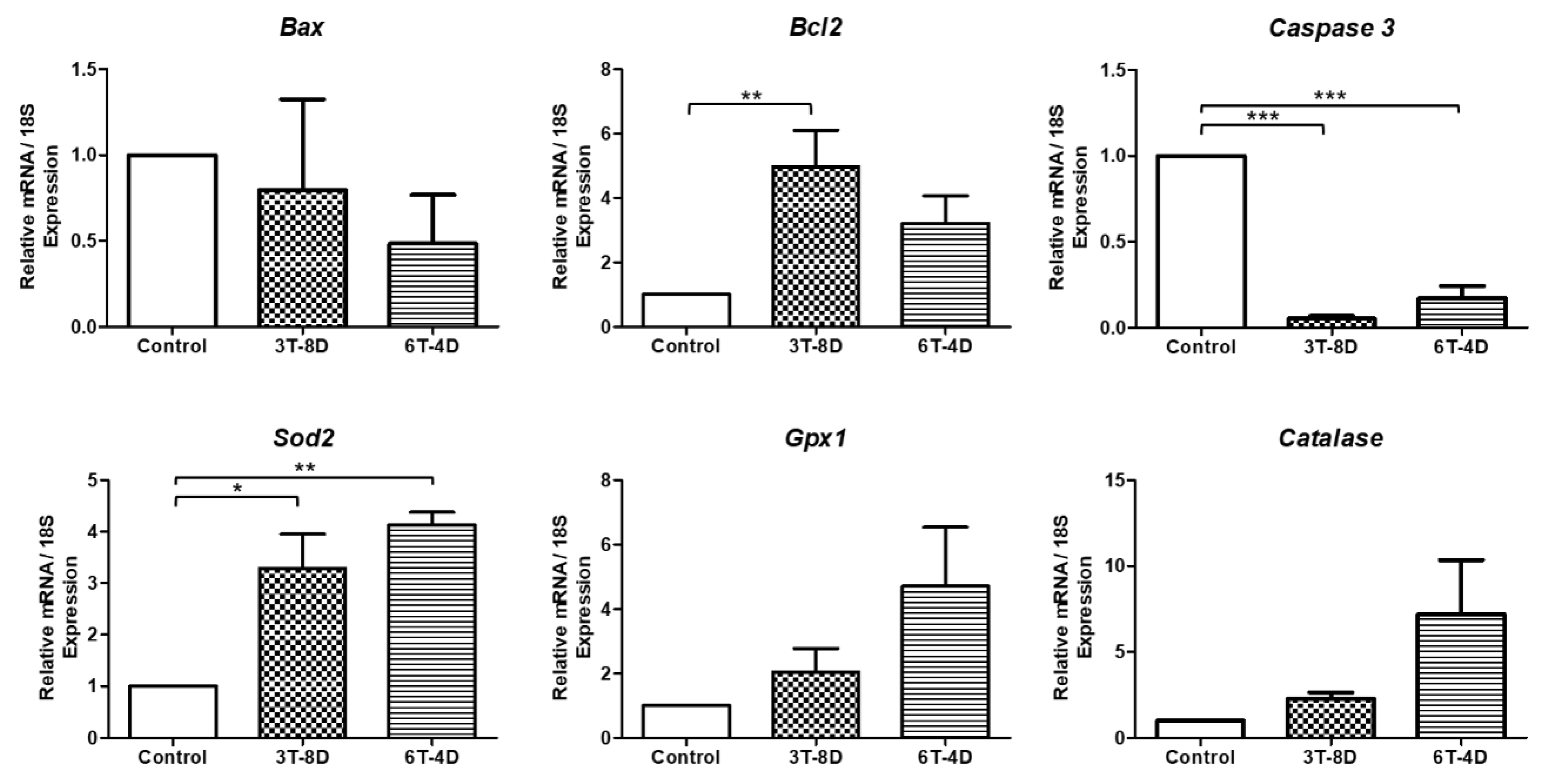
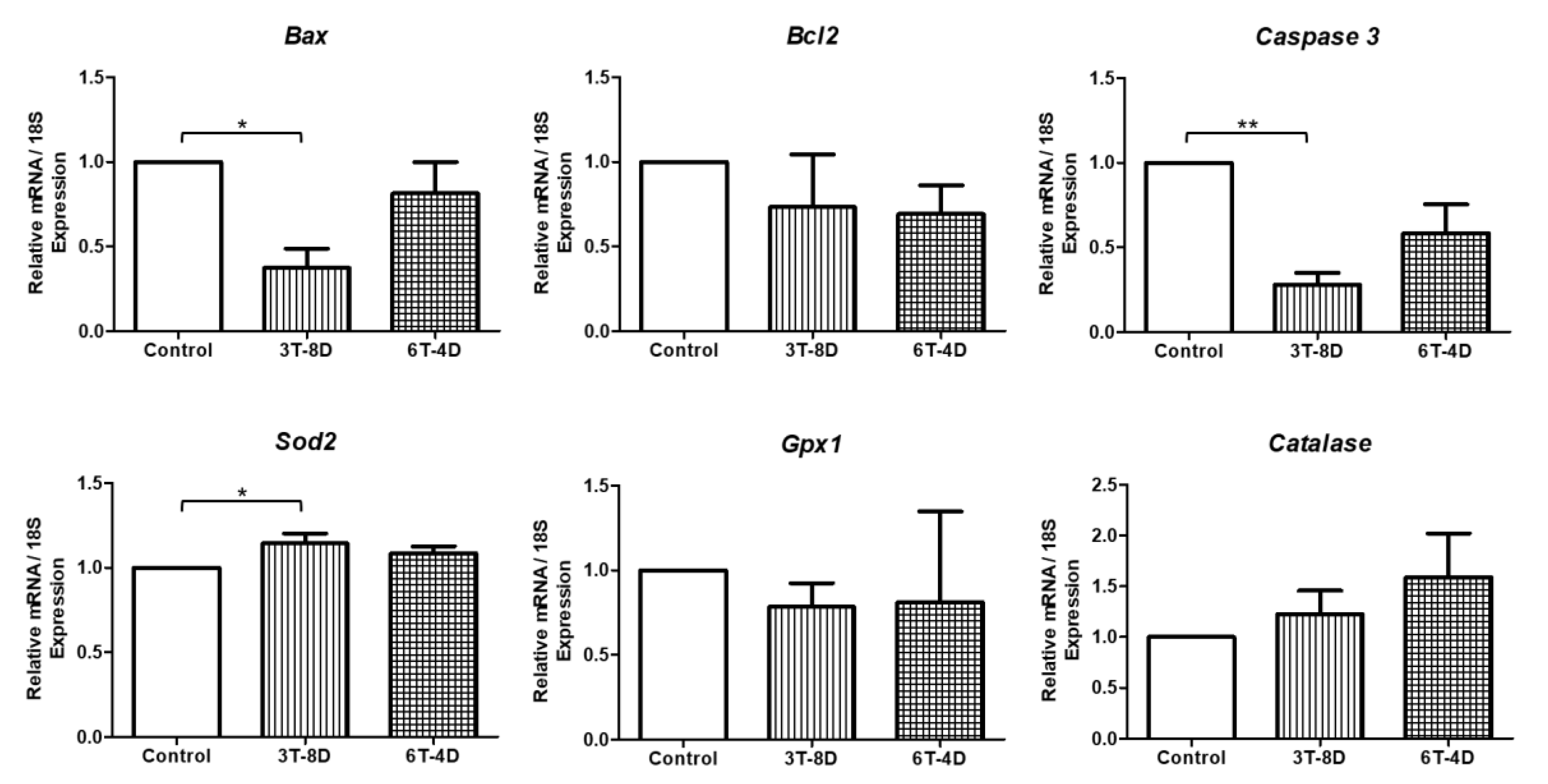
3.4. Anti-Apoptotic and Anti-Oxidative Effects of Human ASC-CMs in the Ovaries of Pregnant Mice
3.5. Anti-Apoptotic and Anti-Oxidative Effects of Human ASC-CM in the Uterus of Pregnant Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- SF2.3: Age of mothers at childbirth and age-specific fertility. Available online: https://www.oecd.org/els/soc/SF_2_3_Age_mothers_childbirth.pdf (accessed on 20 May 2020).
- Shan, D.; Qiu, P.Y.; Wu, Y.X.; Chen, Q.; Li, A.L.; Ramadoss, S.; Wang, R.R.; Hu, Y.Y. Pregnancy Outcomes in Women of Advanced Maternal Age: A Retrospective Cohort Study from China. Sci. Rep. 2018, 8, 12239. [Google Scholar] [CrossRef] [PubMed]
- Joseph, K.S.; Allen, A.C.; Dodds, L.; Turner, L.A.; Scott, H.; Liston, R. The perinatal effects of delayed childbearing. Obstet. Gynecol. 2005, 105, 1410–1418. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.A.; Farquhar, C.; Sullivan, E.A. Donor age is a major determinant of success of oocyte donation/recipient programme. Hum. Reprod. 2012, 27, 118–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fritz, R.; Jindal, S. Reproductive aging and elective fertility preservation. J. Ovarian. Res. 2018, 11, 66. [Google Scholar] [CrossRef] [PubMed]
- Yanez, L.Z.; Han, J.; Behr, B.B.; Pera, R.A.R.; Camarillo, D.B. Human oocyte developmental potential is predicted by mechanical properties within hours after fertilization. Nat. Commun. 2016, 7, 10809. [Google Scholar] [CrossRef] [Green Version]
- Balasch, J.; Gratacos, E. Delayed childbearing: Effects on fertility and the outcome of pregnancy. Fetal. Diagn. Ther. 2011, 29, 263–273. [Google Scholar] [CrossRef]
- Yaron, Y.; Botchan, A.; Amit, A.; Kogosowski, A.; Yovel, I.; Lessing, J.B. Endometrial receptivity: The age-related decline in pregnancy rates and the effect of ovarian function. Fertil. Steril. 1993, 60, 314–318. [Google Scholar] [CrossRef]
- Simchen, M.J.; Shulman, A.; Wiser, A.; Zilberberg, E.; Schiff, E. The aged uterus: Multifetal pregnancy outcome after ovum donation in older women. Hum. Reprod. 2009, 24, 2500–2503. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, A.; Gupta, S.; Sharma, R.K. Role of oxidative stress in female reproduction. Reprod. Biol. Endocrinol. 2005, 3, 28. [Google Scholar] [CrossRef] [Green Version]
- Ra, J.C.; Shin, I.S.; Kim, S.H.; Kang, S.K.; Kang, B.C.; Lee, H.Y.; Kim, Y.J.; Jo, J.Y.; Yoon, E.J.; Choi, H.J.; et al. Safety of intravenous infusion of human adipose tissue-derived mesenchymal stem cells in animals and humans. Stem. Cells. Dev. 2011, 20, 1297–1308. [Google Scholar] [CrossRef]
- Bertolini, F.; Lohsiriwat, V.; Petit, J.Y.; Kolonin, M.G. Adipose tissue cells, lipotransfer and cancer: A challenge for scientists, oncologists and surgeons. Biochim. Biophys. Acta. 2012, 1826, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.S.; Park, B.S.; Sung, J.H. The wound-healing and antioxidant effects of adipose-derived stem cells. Expert. Opin. Biol. Ther. 2009, 9, 879–887. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.X.; Chen, S.R.; Su, P.P.; Huang, F.H.; Shi, Y.C.; Shi, Q.Y.; Lin, S. Using Mesenchymal Stem Cells to Treat Female Infertility: An Update on Female Reproductive Diseases. Stem. Cells. Int. 2019, 2019, 9071720. [Google Scholar] [CrossRef] [PubMed]
- Kay, A.G.; Long, G.; Tyler, G.; Stefan, A.; Broadfoot, S.J.; Piccinini, A.M.; Middleton, J.; Kehoe, O. Mesenchymal Stem Cell-Conditioned Medium Reduces Disease Severity and Immune Responses in Inflammatory Arthritis. Sci. Rep. 2017, 7, 18019. [Google Scholar] [CrossRef]
- Pawitan, J.A. Prospect of stem cell conditioned medium in regenerative medicine. Biomed. Res. Int. 2014, 2014, 965849. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Zhou, J.; Zhang, X.; Liu, Y.; Chen, J.; Hu, B.; Song, J.; Zhang, Y. Strategies to Optimize Adult Stem Cell Therapy for Tissue Regeneration. Int. J. Mol. Sci. 2016, 17, 982. [Google Scholar] [CrossRef] [Green Version]
- Bateman, M.E.; Strong, A.L.; Gimble, J.M.; Bunnell, B.A. Concise Review: Using Fat to Fight Disease: A Systematic Review of Nonhomologous Adipose-Derived Stromal/Stem Cell Therapies. Stem. Cells. 2018, 36, 1311–1328. [Google Scholar] [CrossRef] [Green Version]
- Turner, P.V.; Brabb, T.; Pekow, C.; Vasbinder, M.A. Administration of substances to laboratory animals: Routes of administration and factors to consider. J. Am. Assoc. Lab. Anim. Sci. 2011, 50, 600–613. [Google Scholar]
- Lu, J.; Wang, Z.; Cao, J.; Chen, Y.; Dong, Y. A novel and compact review on the role of oxidative stress in female reproduction. Reprod. Biol. Endocrinol. 2018, 16, 80. [Google Scholar] [CrossRef]
- Peng, C.; Wang, X.; Chen, J.; Jiao, R.; Wang, L.; Li, Y.M.; Zuo, Y.; Liu, Y.; Lei, L.; Ma, K.Y.; et al. Biology of ageing and role of dietary antioxidants. Biomed. Res. Int. 2014, 2014, 831841. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; He, G.; Chen, M.; Zuo, T.; Xu, W.; Liu, X. The Role of Antioxidant Enzymes in the Ovaries. Oxid. Med. Cell. Longev. 2017, 2017, 4371714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narita, M.; Shimizu, S.; Ito, T.; Chittenden, T.; Lutz, R.J.; Matsuda, H.; Tsujimoto, Y. Bax interacts with the permeability transition pore to induce permeability transition and cytochrome c release in isolated mitochondria. Proc. Natl. Acad. Sci. USA 1998, 95, 14681–14686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gross, A.; McDonnell, J.M.; Korsmeyer, S.J. BCL-2 family members and the mitochondria in apoptosis. Genes. Dev. 1999, 13, 1899–1911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Yuan, J. Caspases in apoptosis and beyond. Oncogene 2008, 27, 6194–6206. [Google Scholar] [CrossRef] [Green Version]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Mildmay-White, A.; Khan, W. Cell Surface Markers on Adipose-Derived Stem Cells: A Systematic Review. Current. Stem. Cell. Res. Ther. 2017, 12, 484–492. [Google Scholar] [CrossRef]
- Kim, G.A.; Lee, Y.; Kim, H.J.; Oh, H.J.; Kang, S.K.; Ra, J.C.; Lee, B.C. Intravenous human endothelial progenitor cell administration into aged mice enhances embryo development and oocyte quality by reducing inflammation, endoplasmic reticulum stress and apoptosis. J. Vet. Med. Sci. 2018, 80, 1905–1913. [Google Scholar] [CrossRef]
- Tovey, M.G. Detection and Quantification of Antibodies to Biopharmaceuticals: Practical and ApplieZd Considerations; Wiley: Hoboken, NJ, USA, 2011; pp. 404–412. [Google Scholar]
- Ola, M.S.; Nawaz, M.; Ahsan, H. Role of Bcl-2 family proteins and caspases in the regulation of apoptosis. Mol. Cell. Biochem. 2011, 351, 41–58. [Google Scholar] [CrossRef]
- De Brucker, M.; Tournaye, H.; Haentjens, P.; Verheyen, G.; Collins, J.; Camus, M. Assisted reproduction counseling in women aged 40 and above: A cohort study. J. Assist. Reprod. Genet. 2013, 30, 1431–1438. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Lin, H.; Kong, S.; Wang, S.; Wang, H.; Wang, H.; Armant, D.R. Physiological and molecular determinants of embryo implantation. Mol. Aspects. Med. 2013, 34, 939–980. [Google Scholar] [CrossRef] [Green Version]
- Son, W.Y.; Lee, S.Y.; Lim, J.H. Fertilization, cleavage and blastocyst development according to the maturation timing of oocytes in in vitro maturation cycles. Hum. Reprod. 2005, 20, 3204–3207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krisher, R.L. The effect of oocyte quality on development. J. Anim. Sci. 2004, 82 (E-Suppl), E14–E23. [Google Scholar] [CrossRef]
- Suzuki, H.; Togashi, M.; Moriguchi, Y.; Adachi, J. Relationship between Age-Related Decline in Fertility and In Vitro Fertilization Rate in IVCS Mice. J. Reprod. Develop. 1994, 40, 107–116. [Google Scholar] [CrossRef] [Green Version]
- Shin, H.J.; Cho, Y.M.; Shin, H.J.; Kim, H.D.; Choi, K.M.; Kim, M.G.; Shin, H.D.; Chung, M.W. Comparison of commonly used ICR stocks and the characterization of Korl:ICR. Lab. Anim. Res. 2017, 33, 8–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geisert, R.D.; Schmitt, R.A.M. Early embryonic survival in the pig: Can it be improved? J. Anim. Sci. 2001, 80. [Google Scholar] [CrossRef]
- Fan, Z.; Xiao, Y.; Chen, Y.; Wu, X.; Zhang, G.; Wang, Q.; Xie, C. Effects of catechins on litter size, reproductive performance and antioxidative status in gestating sows. Anim. Nutr. 2015, 1, 271–275. [Google Scholar] [CrossRef]
- Meng, Q.; Guo, T.; Li, G.; Sun, S.; He, S.; Cheng, B.; Shi, B.; Shan, A. Dietary resveratrol improves antioxidant status of sows and piglets and regulates antioxidant gene expression in placenta by Keap1-Nrf2 pathway and Sirt1. J. Anim. Sci. Biotechnol. 2018, 9, 34. [Google Scholar] [CrossRef]
- Nelson, S.M.; Telfer, E.E.; Anderson, R.A. The ageing ovary and uterus: New biological insights. Hum. Reprod. Update. 2013, 19, 67–83. [Google Scholar] [CrossRef] [Green Version]
- Diedrich, K.; Fauser, B.C.; Devroey, P.; Griesinger, G.; The Fifth Evian Annual Reproduction Workshop. The role of the endometrium and embryo in human implantation. Hum. Reprod. Update. 2007, 13, 365–377. [Google Scholar] [CrossRef]
- Alviggi, C.; Humaidan, P.; Howles, C.M.; Tredway, D.; Hillier, S.G. Biological versus chronological ovarian age: Implications for assisted reproductive technology. Reprod. Biol. Endocrinol. 2009, 7, 101. [Google Scholar] [CrossRef] [Green Version]
- Qazi, I.H.; Angel, C.; Yang, H.; Pan, B.; Zoidis, E.; Zeng, C.J.; Han, H.; Zhou, G.B. Selenium, Selenoproteins, and Female Reproduction: A Review. Molecules 2018, 23, 3053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, J.; Luderer, U. Oxidative damage increases and antioxidant gene expression decreases with aging in the mouse ovary. Biol. Reprod. 2011, 84, 775–782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.; Li, S.; Liu, H.; Gong, Y.; Bai, H.; Huang, W.; Liu, Q.; Guan, L.; Fan, P. Association of GPx1 P198L and CAT C-262T Genetic Variations with Polycystic Ovary Syndrome in Chinese Women. Front. Endocrinol. (Lausanne). 2019, 10, 771. [Google Scholar] [CrossRef] [PubMed]
- Baud, O.; Greene, A.E.; Li, J.; Wang, H.; Volpe, J.J.; Rosenberg, P.A. Glutathione peroxidase-catalase cooperativity is required for resistance to hydrogen peroxide by mature rat oligodendrocytes. J. Neurosci. 2004, 24, 1531–1540. [Google Scholar] [CrossRef] [Green Version]
- Molavian, H.; Madani Tonekaboni, A.; Kohandel, M.; Sivaloganathan, S. The Synergetic Coupling among the Cellular Antioxidants Glutathione Peroxidase/Peroxiredoxin and Other Antioxidants and its Effect on the Concentration of H2O2. Sci. Rep. 2015, 5, 13620. [Google Scholar] [CrossRef]
- Kang, W.C.; Oh, P.C.; Lee, K.; Ahn, T.; Byun, K. Increasing injection frequency enhances the survival of injected bone marrow derived mesenchymal stem cells in a critical limb ischemia animal model. Korean. J. Physiol. Pharmacol. 2016, 20, 657–667. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Xiong, Y.Y.; Li, Q.; Hu, M.J.; Huang, P.S.; Xu, J.Y.; Tian, X.Q.; Jin, C.; Liu, J.D.; Qian, L.; et al. Optimization of Timing and Times for Administration of Atorvastatin-Pretreated Mesenchymal Stem Cells in a Preclinical Model of Acute Myocardial Infarction. Stem. Cells. Transl. Med. 2019, 8, 1068–1083. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | GenBank Accession No. | Primer Sequences (5’–3’) | References |
|---|---|---|---|
| 18S rRNA | NR_003278.3 | F: ACCGCGGTTCTATTTTGTTG | [28,29] |
| R: CCCTCTTAATCATGGCCTCA | |||
| Bax | NM_007527.3 | F: ACCAAGAAGCTGAGCGAGTG | [23,24] |
| R: TGCAGCTCCATATTGCTGTC | |||
| Bcl2 | NM_009741.5 | F: ATGATAACCGGGAGATCGTG | [24,30] |
| R: AGCCCCTCTGTGACAGCTTA | |||
| Caspase3 | NM_001284409.1 | F: TGTCATCTCGCTCTGGTACG | [24,25] |
| R: ATTTCAGGCCCATGAATGTC | |||
| Sod2 | NM_013671.3 | F: CTGTCTTCAGCCACACCAGA | [20,21,22] |
| R: CTGCTCTTCCAAAGGTCCTG | |||
| Gpx1 | NM_008160.6 | F: CCGACCCCAAGTACATCATT | [20,21,22] |
| R: CCCACCAGGAACTTCTCAAA | |||
| Catalase | NM_009804.2 | F: TTGACAGAGAGCGGATTCCT | [20,21,22] |
| R: TCTGGTGATATCGTGGGTGA |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ra, K.; Oh, H.J.; Kim, G.A.; Kang, S.K.; Ra, J.C.; Lee, B.C. High Frequency of Intravenous Injection of Human Adipose Stem Cell Conditioned Medium Improved Embryo Development of Mice in Advanced Maternal Age through Antioxidant Effects. Animals 2020, 10, 978. https://doi.org/10.3390/ani10060978
Ra K, Oh HJ, Kim GA, Kang SK, Ra JC, Lee BC. High Frequency of Intravenous Injection of Human Adipose Stem Cell Conditioned Medium Improved Embryo Development of Mice in Advanced Maternal Age through Antioxidant Effects. Animals. 2020; 10(6):978. https://doi.org/10.3390/ani10060978
Chicago/Turabian StyleRa, Kihae, Hyun Ju Oh, Geon A Kim, Sung Keun Kang, Jeong Chan Ra, and Byeong Chun Lee. 2020. "High Frequency of Intravenous Injection of Human Adipose Stem Cell Conditioned Medium Improved Embryo Development of Mice in Advanced Maternal Age through Antioxidant Effects" Animals 10, no. 6: 978. https://doi.org/10.3390/ani10060978