Survey of Serum Amyloid A and Bacterial and Viral Frequency Using qPCR Levels in Recently Captured Feral Donkeys from Death Valley National Park (California)

 ,
,  ,
,  and
and
Abstract
:Simple Summary
Abstract
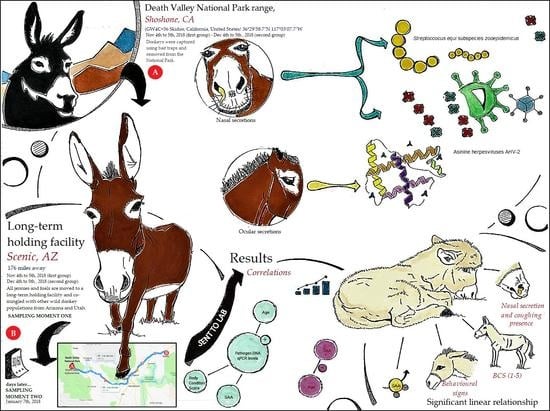
1. Introduction
2. Materials and Methods
2.1. Animal Sample and Sampling
2.2. SAA Levels
2.3. Load Quantification for Viruses and Bacteria
2.4. Quantitative PCR Systems
2.5. RT-Reaction and Quantitative PCR
2.6. Statistical Analyses
2.7. Ethical Approval
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tiller, B. Feral Burro Populations: Distribution and Damage Assessment; Pacific Northwest National Laboratory: Richland, WA, USA, 1997. [Google Scholar]
- Weaver, R.A. Feral burros and wildlife. In Proceedings of the 6th Vertebrate Pest Conference, Anaheim, CA, USA, 5–7 March 1974. [Google Scholar]
- National Research Council. Using Science to Improve the BLM Wild Horse and Burro Program: A Way Forward; National Academies Press: Washington, DC, USA, 2013. [Google Scholar]
- National Research Council/Commission on Natural Resources/National Research Council; Committee on Wild, Free-Roaming Horses Burros. Wild and Free-Roaming Horses and Burros: Current Knowledge and Recommended Research; National Academies Press: Washington, DC, USA, 1980. [Google Scholar]
- Satué, K.; Calvo, A.; Gardón, J.C. Factors influencing serum amyloid type A (SAA) concentrations in horses. Open. J. Vet. Med. 2013, 3, 58. [Google Scholar] [CrossRef] [Green Version]
- Nolen-Walston, R. How to interpret serum amyloid A concentrations. In Proceedings of the 65th Annual American Association of Equine Practitioners (AAEP), Denver, CO, USA, 7–11 December 2019; pp. 130–137. [Google Scholar]
- Kay, G.; Tligui, N.; Semmate, N.; Azrib, R.; González, F.J.N.; Brizgys, L.; McLean, A. Determining factors and interspecific modeling for serum amyloid a concentrations in working horses, donkeys, and mules. Res. Vet. Sci. 2019, 125, 256–265. [Google Scholar] [CrossRef] [PubMed]
- Jacobsen, S.; Andersen, P.H. The acute phase protein serum amyloid A (SAA) as a marker of inflammation in horses. Equine Vet. Educ. 2007, 19, 38–46. [Google Scholar] [CrossRef]
- Pusterla, N.; Kass, P.H.; Mapes, S.; Johnson, C.; Barnett, D.; Vaala, W.; Gutierrez, C.; McDaniel, R.; Whitehead, B.; Manning, J. Surveillance programme for important equine infectious respiratory pathogens in the USA. Vet. Rec. 2011, 169, 12. [Google Scholar] [CrossRef] [PubMed]
- Timoney, J.F. The pathogenic equine streptococci. Vet. Res. 2004, 35, 397–409. [Google Scholar] [CrossRef] [Green Version]
- Wilson, W.D. Equine herpesvirus 1 myeloencephalopathy. Vet. Clin. North Am. Equine Pract. 1997, 13, 53–72. [Google Scholar] [CrossRef]
- Van Maanen, C. Equine herpesvirus 1 and 4 infections: An update. Vet. Q. 2002, 24, 57–78. [Google Scholar] [CrossRef]
- Allen, G.; Kydd, J.; Slater, J.; Smith, K. Recent advances in understanding the pathogenesis, epidemiology, and immunological control of equid herpesvirus-1 (EHV-1) abortion. Proceedings of the 8th International Conference on Equine Infectious Diseases. J. Equine Vet. Sci. 1999, 8, 129–146. [Google Scholar]
- Reed, S.M.; Toribio, R.E. Equine herpesvirus 1 and 4. Vet. Clin. North Am. Equine Pract. 2004, 20, 631–642. [Google Scholar] [CrossRef]
- Ostlund, E.N. The equine herpesviruses. Vet. Clin. North Am. Equine Pract. 1993, 9, 283–294. [Google Scholar] [CrossRef]
- Fortier, G.; Van Erck, E.; Pronost, S.; Lekeux, P.; Thiry, E. Equine gammaherpesviruses: Pathogenesis, epidemiology and diagnosis. Vet. J. 2010, 186, 148–156. [Google Scholar] [CrossRef] [PubMed]
- Ficorilli, N.; Studdert, M.; Crabb, B. The nucleotide sequence of Asinine Herpesvirus 3 glycoprotein G indicates that the donkey virus is closely related to equine herpesvirus 1. Arch. Virol. 1995, 140, 1653–1662. [Google Scholar] [CrossRef] [PubMed]
- Bell, S.A.; Pusterla, N.; Balasuriya, U.B.; Mapes, S.M.; Nyberg, N.L.; Maclachlan, N.J. Isolation of a gammaherpesvirus similar to asinine herpesvirus-2 (AHV-2) from a mule and a survey of mules and donkeys for AHV-2 infection by real-time PCR. Vet. Microbiol. 2008, 130, 176–183. [Google Scholar] [CrossRef]
- Kleiboeker, S.M.; Schommer, S.K.; Johnson, P.J.; Ehlers, B.; Turnquist, S.E.; Boucher, M.; Kreeger, J.M. Association of two newly recognized herpesviruses with interstitial pneumonia in donkeys (Equus asinus). J. Vet. Diagn. Investig. 2002, 14, 273–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleiboeker, S.M.; Turnquist, S.E.; Johnson, P.J.; Kreeger, J.M. Detection and nucleotide sequencing of a DNA-packaging protein gene of equine gammaherpesviruses. J. Vet. Diagn. Investig. 2004, 16, 67–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thiemann, A. Respiratory disease in the donkey. Equine Vet. Educ. 2012, 24, 469–478. [Google Scholar] [CrossRef]
- Vengust, M.; Wen, X.; Bienzle, D. Herpesvirus-associated neurological disease in a donkey. J. Vet. Diagn. Investig. 2008, 20, 820–823. [Google Scholar] [CrossRef]
- Browning, G.; Ficorilli, N.; Studdert, M. Asinine Herpesvirus genomes: Comparison with those of the equine herpesviruses. Arch. Virol. 1988, 101, 183–190. [Google Scholar] [CrossRef]
- Goodrich, E.L.; Behling-Kelly, E. Clinical Pathology of Donkeys and Mules. Vet. Clin. North Am. Equine Pract. 2019, 35, 433–455. [Google Scholar] [CrossRef]
- Valle, E.; Raspa, F.; Giribaldi, M.; Barbero, R.; Bergagna, S.; Antoniazzi, S.; Mc Lean, A.K.; Minero, M.; Cavallarin, L. A functional approach to the body condition assessment of lactating donkeys as a tool for welfare evaluation. PeerJ 2017, 5, e3001. [Google Scholar] [CrossRef] [Green Version]
- Polidori, P.; Vincenzetti, S. Chapter 4. Farm management and feeding strategies for donkey milk production. In Agricultural Research Updates; Mandhatri, P.G.S., Ed.; Nova Science Publishers, Inc.: Hauppauge, NY, USA, 2017; p. 93. [Google Scholar]
- Navas González, F.J.; Jordana Vidal, J.; León Jurado, J.M.; Arando Arbulu, A.; McLean, A.K.; Delgado Bermejo, J.V. Genetic parameter and breeding value estimation of donkeys’ problem-focused coping styles. Behav. Processes 2018, 153, 66–76. [Google Scholar] [CrossRef] [PubMed]
- Veterinary Medicine PCR Laboratory, UCDavis. q-PCR diagnostic submission packet. Available online: https://pcrlab.vetmed.ucdavis.edu/sites/g/files/dgvnsk6571/files/inline-files/DiagnosticPacket08.2019.pdf (accessed on 15 March 2020).
- Derrick, B.; Toher, D.; White, P. How to compare the means of two samples that include paired observations and independent observations: A companion to Derrick, Russ, Toher and White (2017). Tutor. Quant. Methods Psychol. 2017, 13, 120–126. [Google Scholar] [CrossRef] [Green Version]
- Moder, K. Alternatives to F-test in one way ANOVA in case of heterogeneity of variances (a simulation study). Psychol. Test Assess. Model. 2010, 52, 343–353. [Google Scholar]
- Rasch, D.; Kubinger, K.D.; Moder, K. The two-sample t test: Pre-testing its assumptions does not pay off. Stat. Pap. 2011, 52, 219–231. [Google Scholar] [CrossRef]
- IBM Corp. IBM SPSS Statistics for Windows; 25.0; IBM Corp: Armonk, NY, USA, 2017. [Google Scholar]
- Derrick, B. The Partially Overlapping Samples t-test. Package ‘Partiallyoverlapping’ for RStudio. Available online: https://cran.r-project.org/web/packages/Partiallyoverlapping/index.html (accessed on 4 May 2020).
- RStudio Team. RStudio 1.1.463; RStudio: Integrated Development for R. RStudio, Inc.: Boston, MA, USA, 2016. [Google Scholar]
- Oliveira, F.G.; Cook, R.F.; Naves, J.H.F.; Oliveira, C.H.S.; Diniz, R.S.; Freitas, F.J.C.; Lima, J.M.; Sakamoto, S.M.; Leite, R.C.; Issel, C.J.; et al. Equine infectious anemia prevalence in feral donkeys from Northeast Brazil. Prev. Vet. Med. 2017, 140, 30–37. [Google Scholar] [CrossRef]
- Hartley, C.A.; Dynon, K.J.; Mekuria, Z.H.; El-Hage, C.M.; Holloway, S.A.; Gilkerson, J.R. Equine gammaherpesviruses: Perfect parasites? Vet. Microbiol. 2013, 167, 86–92. [Google Scholar] [CrossRef]
- Barrandeguy, M.E.; Carossino, M. Infectious diseases in donkeys and mules: An overview and update. J. Equine Vet. Sci. 2018, 65, 98–105. [Google Scholar] [CrossRef]
- Edington, N.; Bridges, C.; Huckle, A. Experimental reactivation of equid herpesvirus 1 (EHV 1) following the administration of corticosteroids. Equine Vet. J. 1985, 17, 369–372. [Google Scholar] [CrossRef]
- Osterrieder, K. Herpesvirales. In Fenner’s Veterinary Virology, 5th ed.; MacLachlan, N.J., Dubovi, E.J., Eds.; Academic Press: Boston, MA, USA, 2015; pp. 189–216. [Google Scholar]
- Rushton, J.O.; Kolodziejek, J.; Nell, B.; Nowotny, N. Prevalence of Asinine Herpesvirus type 5 (AsHV-5) infection in clinically normal Lipizzaner horses. Vet. J. 2014, 200, 200–203. [Google Scholar] [CrossRef]
- Fortier, G.; Van Erck, E.; Fortier, C.; Richard, E.; Pottier, D.; Pronost, S.; Miszczak, F.; Thiry, E.; Lekeux, P. Herpesviruses in respiratory liquids of horses: Putative implication in airway inflammation and association with cytological features. Vet. Microbiol. 2009, 139, 34–41. [Google Scholar] [CrossRef] [Green Version]
- Goehring, L.S. Donkeys. In Robinson’s Current Therapy in Equine Medicine, 1st ed.; Sprayberry, K.A., Robinson, N.E., Eds.; Elsevier: Philadelphia, PA, USA, 2015; pp. 155–157. [Google Scholar]
- Rickards, K.J.; Thiemann, A.K. Respiratory Disorders of the Donkey. Vet. Clin. N. Am. Equine Pract. 2019, 35, 561–573. [Google Scholar] [CrossRef] [PubMed]
- Laus, F.; Preziuso, S.; Spaterna, A.; Beribe, F.; Tesei, B.; Cuteri, V. Clinical and epidemiological investigation of chronic upper respiratory diseases caused by beta-haemolytic Streptococci in horses. Comp. Immunol. Microbiol. Infect. Dis. 2007, 30, 247–260. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.; Gao, N.; Waller, A.; Cook, F.; Fan, S.; Yuan, D.; Du, Y.; Li, F.; Norimine, J.; Zhu, W. An outbreak of strangles associated with a novel genotype of Streptococcus equi subspecies equi in donkeys in China during 2018. Equine Vet. J. 2019, 51, 743–748. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Assay Name | Gene, NCBI a Accession # | Assay Location (bp) | Amplicon Length (bp) |
|---|---|---|---|
| Asinine Herpesvirus 2 | Polymerase, EU165547 | 100 | 81 |
| Asinine Herpesvirus 3 | Glycoprotein B, U24184 | 740 | 145 |
| Asinine Herpesvirus 5 | Polymerase, AY054993 | 600 | 64 |
| Equine Herpesvirus 1 | Glycoprotein B, NC_001491 | 400 | 89 |
| Equine Herpesvirus 1, neuropathogenic | ORF 30, KF644574 | 200 | 92 |
| Equine Herpesvirus 1, non-neuropathogenic | ORF 30, KX101095 | 200 | 92 |
| Equine Herpesvirus 4 | Glycoprotein B, AF030027 | 440 | 77 |
| Streptococcus equi subspecies equi | M Protein, AF012927 | 150 | 185 |
| Streptococcus equi subspecies zooepidemicus | ITS, EU860336 | 80 | 88 |
| Influenza A (H3N8) | Hemagglutinin Precursor, EF541443 | 350 | 200 |
| Equine rhinitis A virus | RNA polymerase, X96870 | 150 | 111 |
| Equine rhinitis B virus | RNA polymerase, X96871 | 350 | 87 |
| Glyceraldehyde-3-phosphate dehydrogenase | GAPDH, AF097179 | 60 | 105 |
| Parameters | Mean | SEM | SD | Skewness | Kurtosis | |
|---|---|---|---|---|---|---|
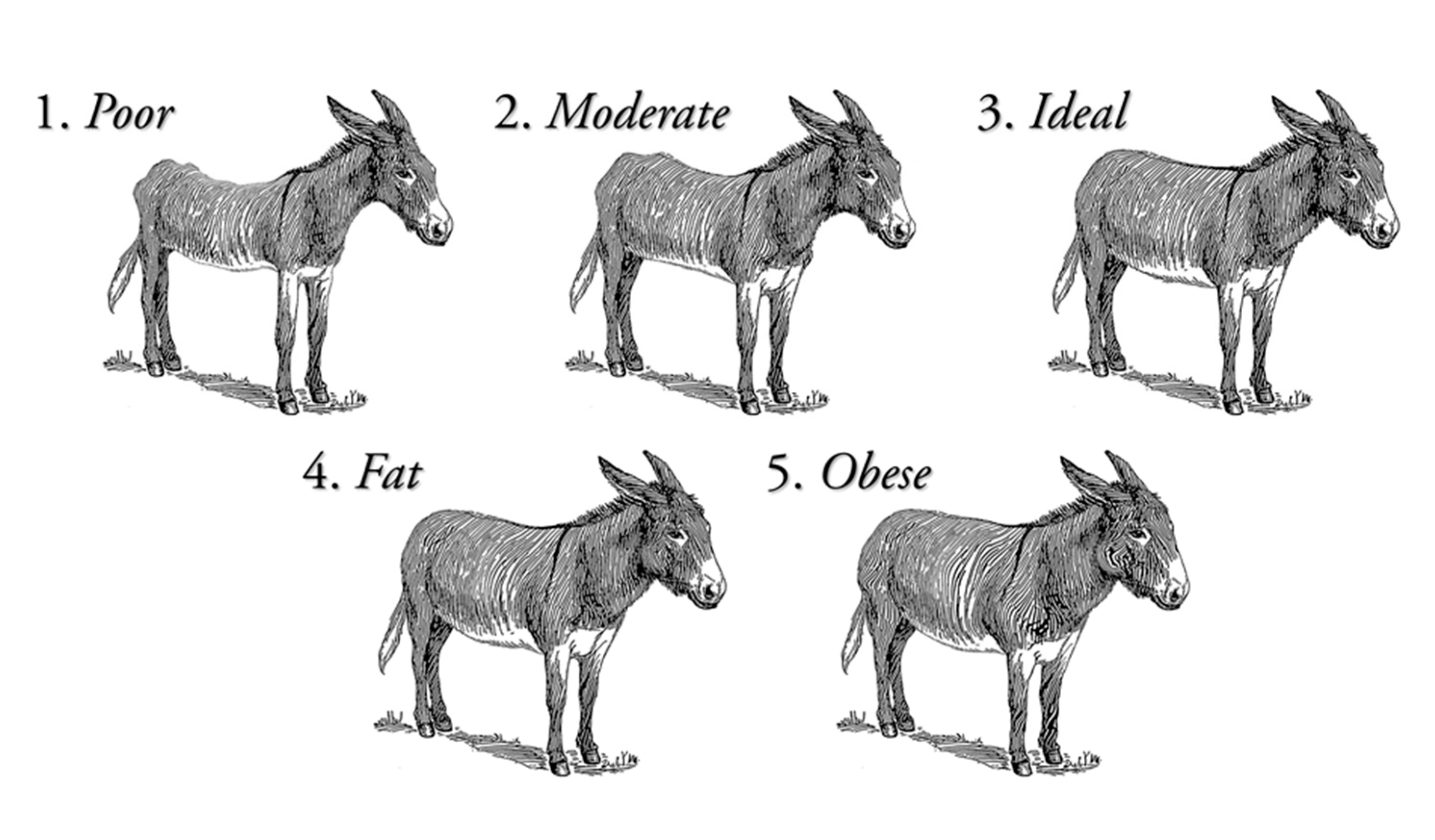
| Clinical Examination Signs | Body condition score (BCS) | 3.12 | 0.09 | 0.83 | −0.15 | −0.96 |
| Behavior signs | 2.11 | 0.04 | 0.38 | 1.14 | 3.13 | |
| Skin/hair condition | 1.29 | 0.10 | 0.94 | 3.21 | 9.14 | |
| Lameness presence | 1.96 | 0.02 | 0.19 | −5.13 | 24.88 | |
| Ocular discharge presence | 1.82 | 0.04 | 0.38 | −1.73 | 1.01 | |
| Nasal discharge presence | 1.66 | 0.05 | 0.48 | −0.68 | −1.57 | |
| Abnormal breathing presence | 1.07 | 0.03 | 0.26 | 3.41 | 9.88 | |
| Coughing presence | 1.93 | 0.03 | 0.26 | −3.41 | 9.88 | |
| SAA | SAA maximal concentration (mg/L) | 10.80 | 3.72 | 34.27 | 5.42 | 33.75 |
| Pathogen Load qPCR-Assay | Asinine Herpesvirus 2 (AHV-2) aka EHV7 | 39.69 | 0.14 | 1.30 | −4.33 | 18.30 |
| Asinine Herpesvirus 3 (AHV-3) aka EHV8 | 39.74 | 0.14 | 1.30 | −5.92 | 36.83 | |
| Asinine Herpesvirus 5 (AHV-5) | 34.39 | 0.50 | 4.59 | −0.51 | −0.38 | |
| Equine Herpesvirus 1 (EHV-1) | 40.00 | 0.00 | 0.00 | N/A | N/A | |
| Equine Herpesvirus 1 (EHV-1) neuropathogenic | 40.00 | 0.00 | 0.00 | N/A | N/A | |
| Equine Herpesvirus 1 (EHV-1) non-neuropathogenic | 40.00 | 0.00 | 0.00 | N/A | N/A | |
| Equine herpesvirus 4 (EHV-4) | 40.00 | 0.00 | 0.00 | N/A | N/A | |
| Streptococcus equi subspecies equi | 39.94 | 0.04 | 0.39 | −6.77 | 46.45 | |
| Streptococcus equi subspecies zooepidemicus | 35.04 | 0.44 | 4.08 | -0.75 | 0.07 | |
| Influenza AH3N8 | 40.00 | 0.00 | 0.00 | N/A | N/A | |
| Equine rhinitis A virus | 40.00 | 0.00 | 0.00 | N/A | N/A | |
| Equine rhinitis B virus | 40.00 | 0.00 | 0.00 | N/A | N/A | |
| Controls | Glyceraldehyde 3 phosphate dehydrogenase (First control) | 29.24 | 0.38 | 3.49 | 0.83 | 1.91 |
| Glyceraldehyde 3 phosphate dehydrogenase (Second control) | 32.13 | 0.41 | 3.78 | 0.27 | −0.87 | |
| Fibrinogen | 0.44 | 0.09 | 0.50 | 4.56 | 23.39 |
| Parameters | Mean | SEM | SD | Skewness | Kurtosis | |
|---|---|---|---|---|---|---|
| Clinical Examination Signs | Body condition score | 2.43 | 0.08 | 0.37 | 0.97 | 1.81 |
| Behavior signs | 1.96 | 0.07 | 0.36 | −0.65 | 6.34 | |
| Skin/Hair condition | 1.00 | 0.00 | 0.00 | N/A | N/A | |
| Lameness presence | 2.00 | 0.00 | 0.00 | N/A | N/A | |
| Ocular discharge presence | 1.87 | 0.07 | 0.34 | −2.42 | 4.21 | |
| Nasal discharge presence | 1.42 | 0.10 | 0.50 | 0.36 | −2.05 | |
| Abnormal breathing presence | 1.13 | 0.07 | 0.34 | 2.42 | 4.21 | |
| Coughing presence | 1.13 | 0.07 | 0.34 | 2.42 | 4.21 | |
| SAA | SAA maximal concentration (mg/L) | 17.42 | 10.73 | 52.58 | 4.43 | 20.53 |
| Pathogen Load qPCR-Assay | Asinine Herpesvirus 2 (AHV-2) aka EHV7 | 38.03 | 0.90 | 4.40 | −2.57 | 6.18 |
| Asinine Herpesvirus 3 (AHV-3) aka EHV8 | 39.94 | 0.06 | 0.31 | −4.90 | 24.00 | |
| Asinine Herpesvirus 5 (AHV-5) | 32.65 | 0.89 | 4.36 | −1.37 | 2.84 | |
| Equine Herpesvirus 1 (EHV-1) | 40.00 | 0.00 | 0.00 | N/A | N/A | |
| Equine Herpesvirus 1 (EHV-1) neuropathogenic | 40.00 | 0.00 | 0.00 | N/A | N/A | |
| Equine Herpesvirus 1 (EHV-1) non-neuropathogenic | 40.00 | 0.00 | 0.00 | N/A | N/A | |
| Equine Herpesvirus 4 (EHV-4) | 40.00 | 0.00 | 0.00 | N/A | N/A | |
| Streptococcus equi subspecies equi | 40.00 | 0.00 | 0.00 | N/A | N/A | |
| Streptococcus equi subspecies zooepidemicus | 33.04 | 0.69 | 3.38 | -0.65 | 0.77 | |
| Influenza AH3N8 | 40.00 | 0.00 | 0.00 | N/A | N/A | |
| Equine rhinitis A virus | 40.00 | 0.00 | 0.00 | N/A | N/A | |
| Equine rhinitis B virus | 40.00 | 0.00 | 0.00 | N/A | N/A | |
| Controls | Glyceraldehyde 3 phosphate dehydrogenase (First control) | 23.61 | 0.97 | 4.76 | 2.04 | 5.02 |
| Glyceraldehyde 3 phosphate dehydrogenase (Second control) | 29.37 | 0.85 | 4.17 | 1.45 | 1.75 | |
| Fibrinogen | 0.63 | 0.54 | 2.67 | 4.73 | 22.74 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jerele, S.; Davis, E.; Mapes, S.; Pusterla, N.; Navas González, F.J.; Iglesias Pastrana, C.; Abdelfattah, E.M.; McLean, A. Survey of Serum Amyloid A and Bacterial and Viral Frequency Using qPCR Levels in Recently Captured Feral Donkeys from Death Valley National Park (California). Animals 2020, 10, 1086. https://doi.org/10.3390/ani10061086
Jerele S, Davis E, Mapes S, Pusterla N, Navas González FJ, Iglesias Pastrana C, Abdelfattah EM, McLean A. Survey of Serum Amyloid A and Bacterial and Viral Frequency Using qPCR Levels in Recently Captured Feral Donkeys from Death Valley National Park (California). Animals. 2020; 10(6):1086. https://doi.org/10.3390/ani10061086
Chicago/Turabian StyleJerele, Sara, Eric Davis, Samantha Mapes, Nicola Pusterla, Francisco Javier Navas González, Carlos Iglesias Pastrana, Essam Mahmoud Abdelfattah, and Amy McLean. 2020. "Survey of Serum Amyloid A and Bacterial and Viral Frequency Using qPCR Levels in Recently Captured Feral Donkeys from Death Valley National Park (California)" Animals 10, no. 6: 1086. https://doi.org/10.3390/ani10061086