Molecular Characterization, Expression Profiling, and SNP Analysis of the Porcine RNF20 Gene


Abstract
:Simple Summary
Abstract
1. Introduction
2. Methods and Materials
2.1. Populations and DNA Samples
2.2. PCR Amplification
2.3. Bioinformatic Analysis
2.4. Western Blot Analysis
2.5. PCR–RFLP
2.6. Association Analysis
3. Result
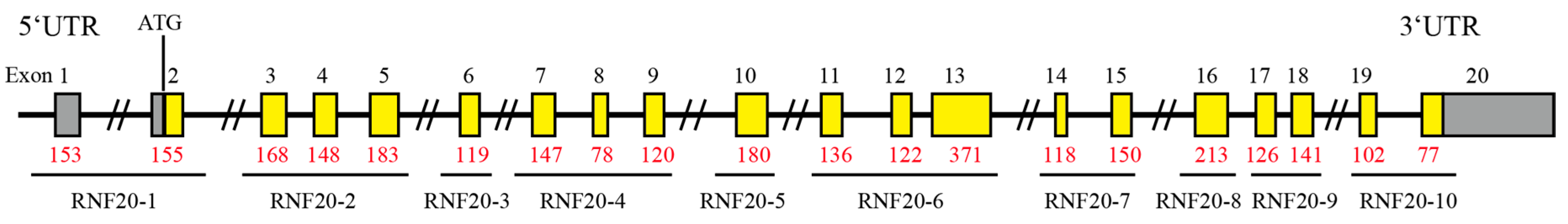
3.1. Exon Sequences of the Porcine RNF20 Gene
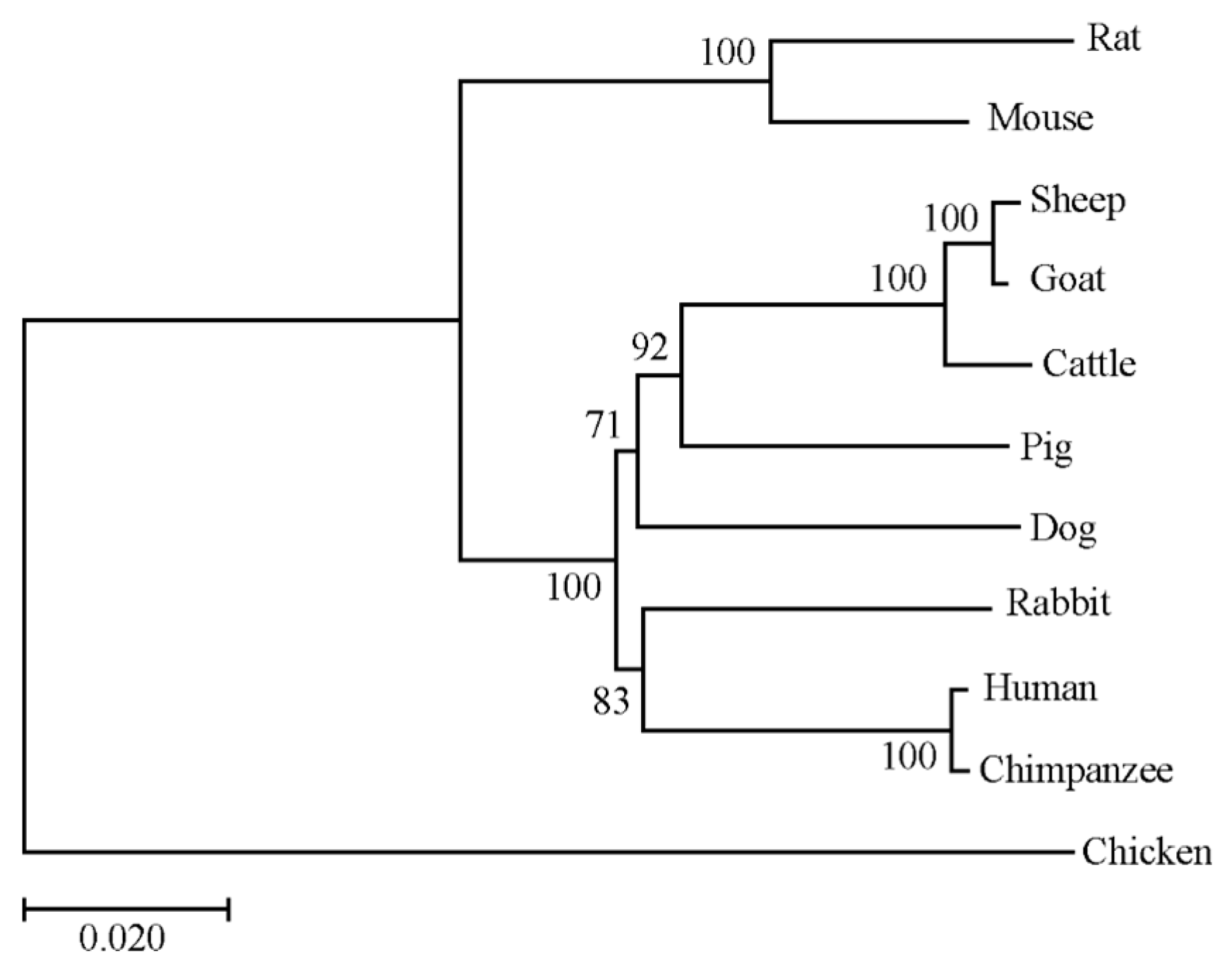
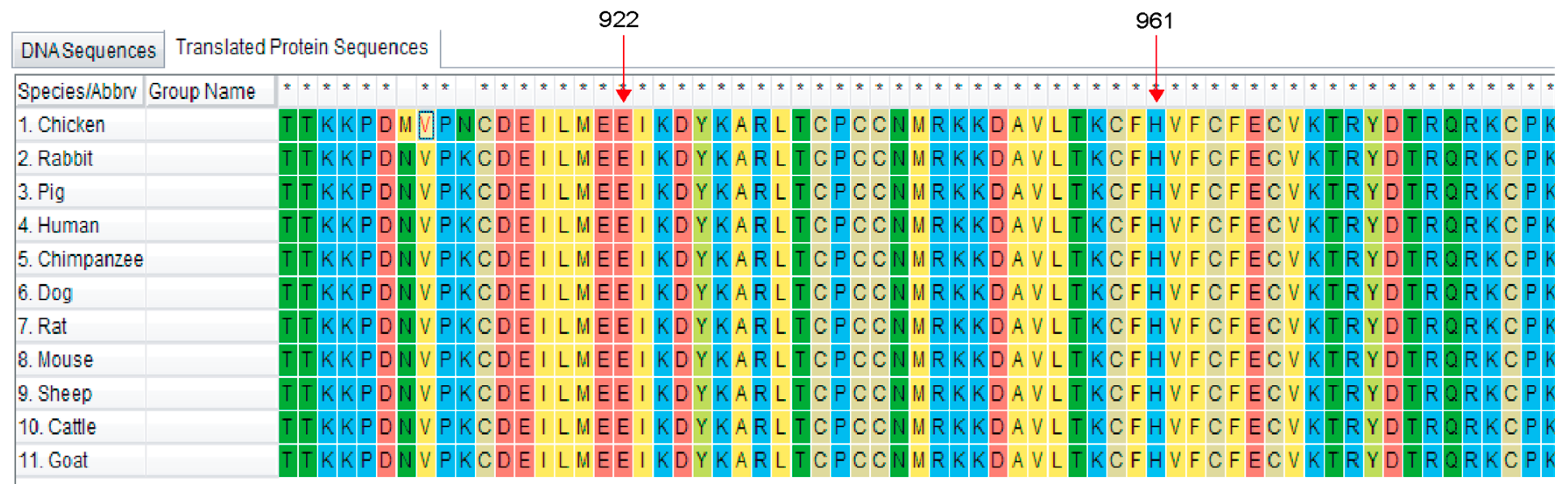
3.2. Bioinformatic Analysis of the RNF20 Gene
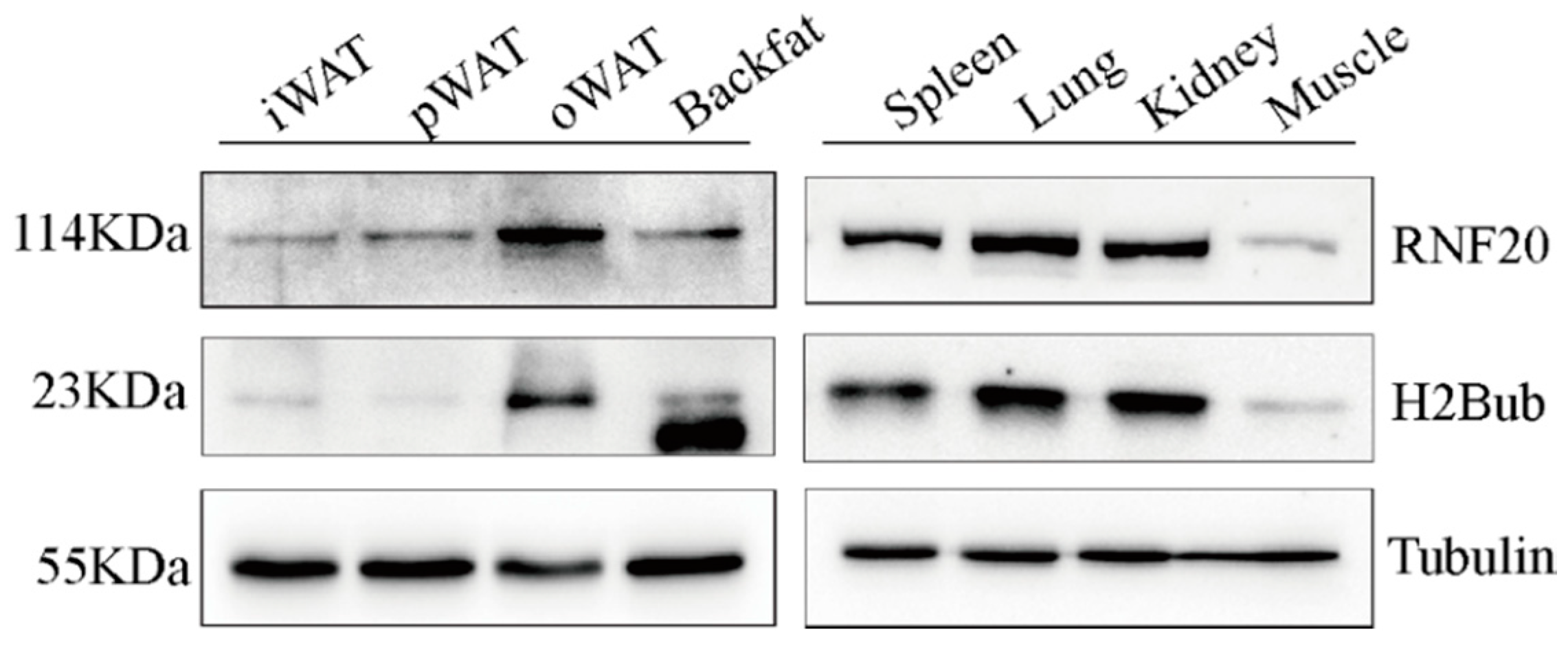
3.3. RNF20 Is Ubiquitously Expressed in Porcine Tissues
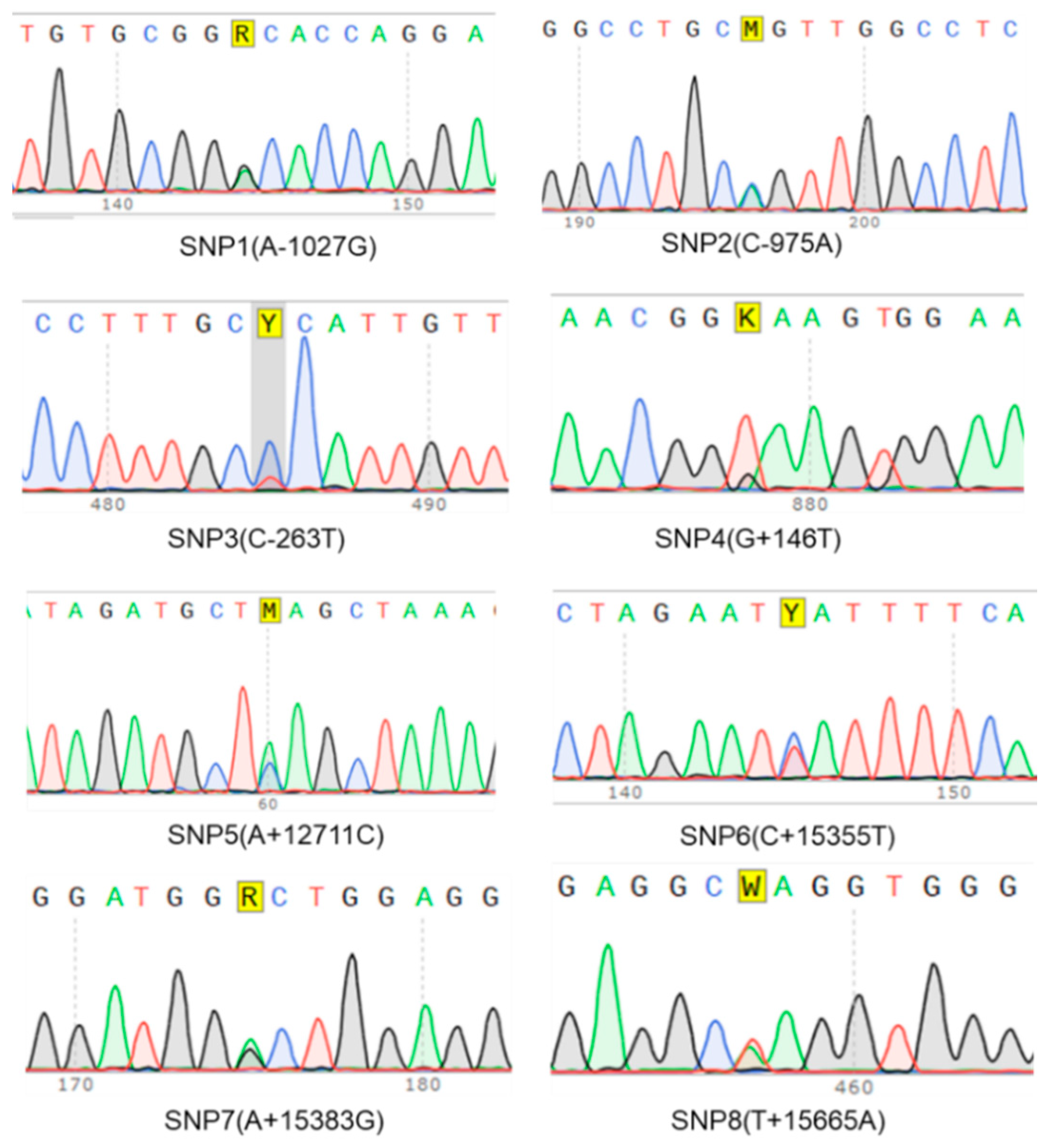
3.4. Potential SNPs Identification
3.5. Genotype and Allele Distribution of the RNF20 Gene
3.6. Association Analysis of SNP1 with Backfat Thickness
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Hwang, W.W.; Venkatasubrahmanyam, S.; Ianculescu, A.G.; Tong, A.; Boone, C.; Madhani, H.D. A conserved RING finger protein required for histone H2B monoubiquitination and cell size control. Mol. Cell 2003, 11, 261–266. [Google Scholar] [CrossRef]
- Trujillo, K.M.; Osley, M.A. A role for H2B ubiquitylation in DNA replication. Mol. Cell 2012, 48, 734–746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wright, D.E.; Kao, C.F. (Ubi) quitin’ the h2bit: Recent insights into the roles of H2B ubiquitylation in DNA replication and transcription. Epigenetics 2015, 10, 122–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fleming, A.B.; Kao, C.F.; Hillyer, C.; Pikaart, M.; Osley, M.A. H2B ubiquitylation plays a role in nucleosome dynamics during transcription elongation. Mol. Cell 2008, 31, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Yan, Q. Histone ubiquitination and deubiquitination in transcription, DNA damage response, and cancer. Front. Oncol. 2012, 2, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uckelmann, M.; Sixma, T.K. Histone ubiquitination in the DNA damage response. DNA Repair 2017, 56, 92–101. [Google Scholar] [CrossRef]
- Chernikova, S.B.; Razorenova, O.V.; Higgins, J.P.; Sishc, B.J.; Nicolau, M.; Dorth, J.A.; Chernikova, D.A.; Kwok, S.; Brooks, J.D.; Bailey, S.M.; et al. Deficiency in mammalian histone H2B ubiquitin ligase Bre1 (Rnf20/Rnf40) leads to replication stress and chromosomal instability. Cancer Res. 2012, 72, 2111–2119. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Chen, S.; Wang, S.; Soares, F.; Fischer, M.; Meng, F.; Du, Z.; Lin, C.; Meyer, C.; DeCaprio, J.A.; et al. Transcriptional landscape of the human cell cycle. Proc. Natl. Acad. Sci. USA 2017, 114, 3473–3478. [Google Scholar] [CrossRef] [Green Version]
- Ishino, Y.; Hayashi, Y.; Naruse, M.; Tomita, K.; Sanbo, M.; Fuchigami, T.; Fujiki, R.; Hirose, K.; Toyooka, Y.; Fujimori, T.; et al. Bre1a, a histone H2B ubiquitin ligase, regulates the cell cycle and differentiation of neural precursor cells. J. Neurosci. 2014, 34, 3067–3078. [Google Scholar] [CrossRef] [Green Version]
- Fuchs, G.; Shema, E.; Vesterman, R.; Kotler, E.; Wolchinsky, Z.; Wilder, S.; Golomb, L.; Pribluda, A.; Zhang, F.; Haj-Yahya, M.; et al. RNF20 and USP44 regulate stem cell differentiation by modulating H2B monoubiquitylation. Mol. Cell 2012, 46, 662–673. [Google Scholar] [CrossRef] [Green Version]
- Karpiuk, O.; Najafova, Z.; Kramer, F.; Hennion, M.; Galonska, C.; Konig, A.; Snaidero, N.; Vogel, T.; Shchebet, A.; Begus-Nahrmann, Y.; et al. The histone H2B monoubiquitination regulatory pathway is required for differentiation of multipotent stem cells. Mol. Cell 2012, 46, 705–713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shema, E.; Tirosh, I.; Aylon, Y.; Huang, J.; Ye, C.; Moskovits, N.; Raver-Shapira, N.; Minsky, N.; Pirngruber, J.; Tarcic, G.; et al. The histone H2B-specific ubiquitin ligase RNF20/hBRE1 acts as a putative tumor suppressor through selective regulation of gene expression. Genes Dev. 2008, 22, 2664–2676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.H.; Jeon, Y.G.; Lee, K.H.; Lee, H.W.; Park, J.; Jang, H.; Kang, M.; Lee, H.S.; Cho, H.J.; Nam, D.H.; et al. RNF20 Suppresses Tumorigenesis by Inhibiting the SREBP1c-PTTG1 Axis in Kidney Cancer. Mol. Cell Biol. 2017, 37, e00265–e00271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kosinsky, R.L.; Chua, R.L.; Qui, M.; Saul, D.; Mehlich, D.; Strobel, P.; Schildhaus, H.U.; Wegwitz, F.; Faubion, W.A.; Johnsen, S.A. Loss of RNF40 Decreases NF-kappaB Activity in Colorectal Cancer Cells and Reduces Colitis Burden in Mice. J. Crohns. Colitis. 2019, 13, 362–373. [Google Scholar] [CrossRef] [PubMed]
- Foglizzo, M.; Middleton, A.J.; Day, C.L. Structure and Function of the RING Domains of RNF20 and RNF40, Dimeric E3 Ligases that Monoubiquitylate Histone H2B. J. Mol. Biol. 2016, 428, 4073–4086. [Google Scholar] [CrossRef]
- Kim, J.; Hake, S.B.; Roeder, R.G. The human homolog of yeast BRE1 functions as a transcriptional coactivator through direct activator interactions. Mol. Cell 2005, 20, 759–770. [Google Scholar] [CrossRef]
- Lee, J.H.; Lee, G.Y.; Jang, H.; Choe, S.S.; Koo, S.H.; Kim, J.B. Ring finger protein20 regulates hepatic lipid metabolism through protein kinase A-dependent sterol regulatory element binding protein1c degradation. Hepatology 2014, 60, 844–857. [Google Scholar] [CrossRef] [Green Version]
- Jeon, Y.G.; Lee, J.H.; Ji, Y.; Sohn, J.H.; Lee, D.; Kim, D.W.; Yoon, S.G.; Shin, K.C.; Park, J.; Seong, J.K.; et al. RNF20 Functions as a Transcriptional Coactivator for PPARgamma by Promoting NCoR1 Degradation in Adipocytes. Diabetes 2020, 69, 20–34. [Google Scholar] [CrossRef] [Green Version]
- Wood, J.D.; Enser, M.; Fisher, A.V.; Nute, G.R.; Sheard, P.R.; Richardson, R.I.; Hughes, S.I.; Whittington, F.M. Fat deposition, fatty acid composition and meat quality: A review. Meat Sci. 2008, 78, 343–358. [Google Scholar] [CrossRef]
- Chen, J.N.; Jiang, Y.Z.; Cen, W.M.; Xing, S.H.; Zhu, L.; Tang, G.Q.; Li, M.Z.; Jiang, A.A.; Lou, P.E.; Wen, A.X.; et al. Distribution of H-FABP and ACSL4 gene polymorphisms and their associations with intramuscular fat content and backfat thickness in different pig populations. Genet. Mol. Res. 2014, 13, 6759–6772. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, J.; Gong, H.; Cui, L.; Zhang, W.; Ma, J.; Chen, C.; Ai, H.; Xiao, S.; Huang, L.; et al. Genetic correlation of fatty acid composition with growth, carcass, fat deposition and meat quality traits based on GWAS data in six pig populations. Meat Sci. 2019, 150, 47–55. [Google Scholar] [CrossRef]
- Wang, L.; Cao, C.; Wang, F.; Zhao, J.; Li, W. H2B ubiquitination: Conserved molecular mechanism, diverse physiologic functions of the E3 ligase during meiosis. Nucleus 2017, 8, 461–468. [Google Scholar] [CrossRef] [Green Version]
- Ren, P.; Sheng, Z.; Wang, Y.; Yi, X.; Zhou, Q.; Zhou, J.; Xiang, S.; Hu, X.; Zhang, J. RNF20 promotes the polyubiquitination and proteasome-dependent degradation of AP-2alpha protein. Acta Biochim. Biophys. Sin. 2014, 46, 136–140. [Google Scholar] [CrossRef] [Green Version]
- Coelho, M.; Oliveira, T.; Fernandes, R. Biochemistry of adipose tissue: An endocrine organ. Arch. Med. Sci. 2013, 9, 191–200. [Google Scholar] [CrossRef] [Green Version]
- Fontana, L.; Eagon, J.C.; Trujillo, M.E.; Scherer, P.E.; Klein, S. Visceral fat adipokine secretion is associated with systemic inflammation in obese humans. Diabetes 2007, 56, 1010–1013. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Song, Z.; Li, G.; Tu, H.; Liu, W.; Liu, Y.; Wang, P.; Wang, Y.; Cui, X.; Liu, C.; et al. H2B ubiquitination regulates meiotic recombination by promoting chromatin relaxation. Nucleic Acids Res. 2016, 44, 9681–9697. [Google Scholar] [CrossRef] [Green Version]
- Tarcic, O.; Pateras, I.S.; Cooks, T.; Shema, E.; Kanterman, J.; Ashkenazi, H.; Boocholez, H.; Hubert, A.; Rotkopf, R.; Baniyash, M.; et al. RNF20 Links Histone H2B Ubiquitylation with Inflammation and Inflammation-Associated Cancer. Cell Rep. 2016, 14, 1462–1476. [Google Scholar] [CrossRef] [Green Version]
- Vethantham, V.; Yang, Y.; Bowman, C.; Asp, P.; Lee, J.H.; Skalnik, D.G.; Dynlacht, B.D. Dynamic loss of H2B ubiquitylation without corresponding changes in H3K4 trimethylation during myogenic differentiation. Mol. Cell Biol. 2012, 32, 1044–1055. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Primer Sequences | Temp. (°C) | Product Sizes (bp) | Amplification Region |
|---|---|---|---|---|
| RNF20-1 | F: TTTTCCTCTCCCTGACTCCTC | 59.8 | 1427 | Exon1/2 |
| R: TGTTACTCCAGAAGGCTTCCA | ||||
| RNF20-2 | F: GTCAAGATTCCTCCCAGCTTC | 60.5 | 1180 | Exon3/4/5 |
| R: TGGCAGCTATAGTTCCGATCA | ||||
| RNF20-3 | F: TAGCATGTCATTGCTTCGTTG | 60.0 | 286 | Exon6 |
| R: GACGAGCTTCAAAGCATTCAG | ||||
| RNF20-4 | F: TGAACACCTCTCTTTGGGATG | 59.1 | 960 | Exon7/8/9 |
| R: TGTGTGTGAGCTAATCAGCAA | ||||
| RNF20-5 | F: TAGCATCCAGGGCACAGATAC | 60.1 | 389 | Exon10 |
| R: GCCCACCATGTAGCAGAGTAA | ||||
| RNF20-6 | F: TGTTGATTGGCTCCTTCTCTG | 59.9 | 1275 | Exon11/12/13 |
| R: GAACCAGACCACATGATAGCC | ||||
| RNF20-7 | F: CTGATGCCCTTTGTTTTCTCA | 59.4 | 1189 | Exon14/15 |
| R: TCCTCTGATTCCAGAAAGCTC | ||||
| RNF20-8 | F: TTGGGACTAGCAGATGCAGA | 59.8 | 397 | Exon16 |
| R: TTCCAAAATTCTGTTGAAGAG | ||||
| RNF20-9 | F: AACAGATTTTTAGGCCTGTGG | 60.0 | 591 | Exon17/18 |
| R: TAGGTGGTTTCTGAACTGTGA | ||||
| RNF20-10 | F: TGGGGAAGTGTGTAATGGGTA | 60.2 | 600 | Exon19/20 |
| R: TAGCCAGCTCGTCGTCTTCT |
| Breeds | Number | Genotype Frequency (Number) | Allele Frequency | |||
|---|---|---|---|---|---|---|
| GG | AG | AA | G | A | ||
| Yorkshire | 64 | 0.52 (33) | 0.42 (27) | 0.063 (4) | 0.73 | 0.27 |
| Landrace | 165 | 0.88 (145) | 0.12 (20) | 0.00 | 0.94 | 0.06 |
| Duroc | 37 | 0.60 (22) | 0.40 (15) | 0.00 | 0.80 | 0.20 |
| Min pig | 30 | 1.00 (30) | 0.00 (0) | 0.00 | 1 | 0.00 |
| Breeds | Genotype (Number) | BFT (mm) | p-Value |
|---|---|---|---|
| Yorkshire | GG (33) | 13.87 ± 1.47 | GG-AG 0.7709 |
| AG (27) | 12.71 ± 0.57 | AG-AA 0.5337 | |
| AA (4) | 13.19 ± 0.51 | AA-GG 0.6194 | |
| Landrace | GG (145) | 12.89 ± 0.25 | GG-AG 0.8668 |
| AG (20) | 12.93 ± 0.69 | ||
| Duroc | GG (22) | 12.73 ± 0.76 | GG-AG 0.7606 |
| AG (15) | 11.75 ± 0.92 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Y.; Yang, S.; Wang, Y.; Tao, C. Molecular Characterization, Expression Profiling, and SNP Analysis of the Porcine RNF20 Gene. Animals 2020, 10, 888. https://doi.org/10.3390/ani10050888
Zhao Y, Yang S, Wang Y, Tao C. Molecular Characterization, Expression Profiling, and SNP Analysis of the Porcine RNF20 Gene. Animals. 2020; 10(5):888. https://doi.org/10.3390/ani10050888
Chicago/Turabian StyleZhao, Ying, Shulin Yang, Yanfang Wang, and Cong Tao. 2020. "Molecular Characterization, Expression Profiling, and SNP Analysis of the Porcine RNF20 Gene" Animals 10, no. 5: 888. https://doi.org/10.3390/ani10050888