Mobility and Invasion Related Gene Expression Patterns in Equine Sarcoid

 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results
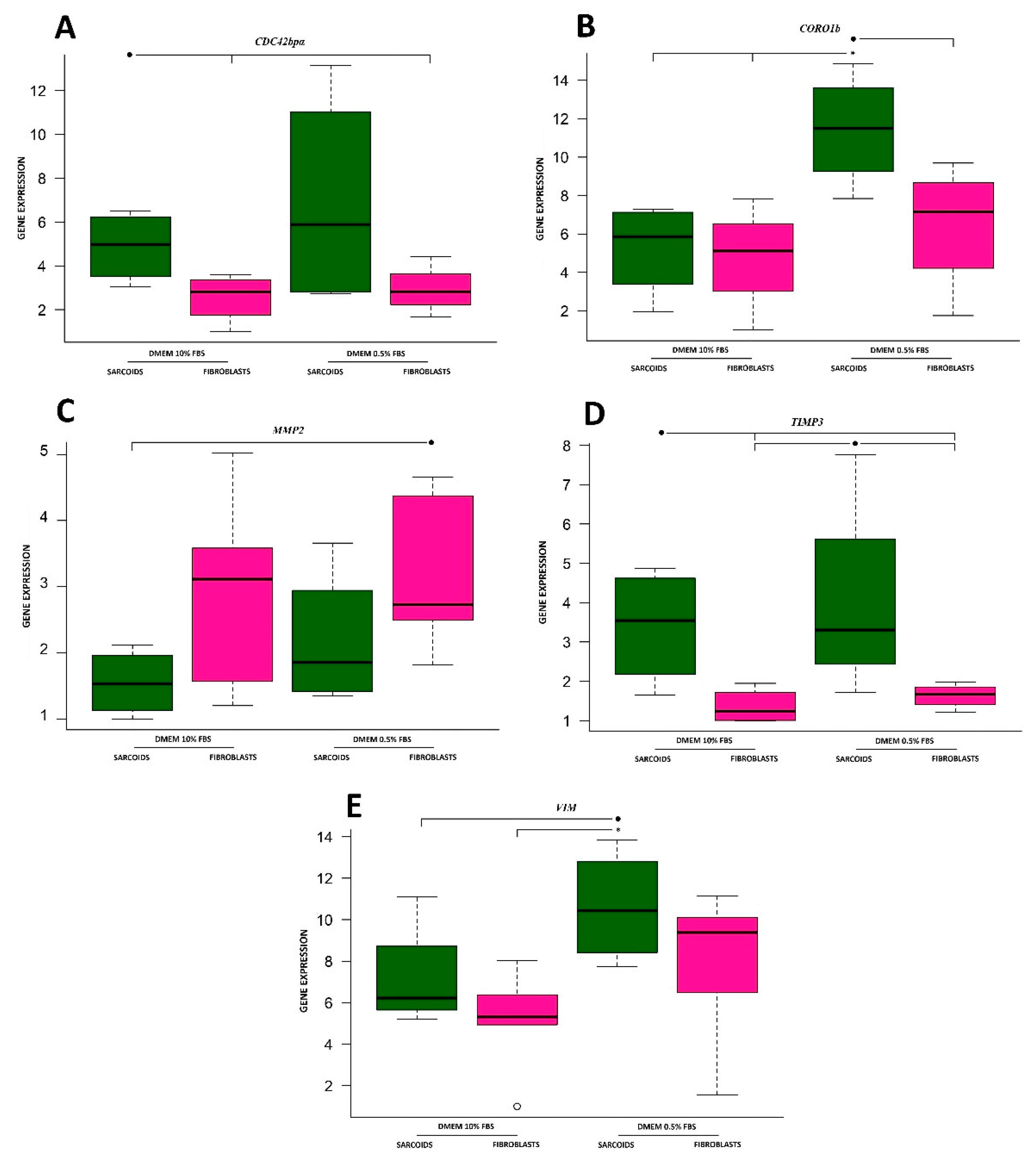
3.1. Cell Division Control 42 Binding Protein Alpha
3.2. Coronin
3.3. Matrix Metalloproteinase 2
3.4. Tissue Inhibitor of Metalloproteinases 3
3.5. Vimentin
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Knottenbelt, D.C. A Suggested Clinical Classification for the Equine Sarcoid. Clin. Tech. Equine Pr. 2005, 4, 278–295. [Google Scholar] [CrossRef]
- Haralambus, R.; Burgstaller, J.; Klukowska-Rötzler, J.; Steinborn, R.; Buchinger, S.; Gerber, V.; Brandt, S. Intralesional bovine papillomavirus DNA loads reflect severity of equine sarcoid disease. Equine Veter-J. 2010, 42, 327–331. [Google Scholar] [CrossRef] [PubMed]
- Christen, G.; Gerber, V.; Dolf, G.; Burger, D.; Koch, C. Inheritance of equine sarcoid disease in Franches-Montagnes horses. Veter-J. 2014, 199, 68–71. [Google Scholar] [CrossRef] [PubMed]
- Knottenbelt, D.C.; Matthews, J. A Positive Step Forwards in the Diagnosis of Equine Sarcoid. Veter-J. 2001, 161, 224–226. [Google Scholar] [CrossRef] [PubMed]
- Taylor, S.D.; Toth, B.; Baseler, L.J.; Charney, V.A.; Miller, M.A. Lack of Correlation Between Papillomaviral DNA in Surgical Margins and Recurrence of Equine Sarcoids. J. Equine Veter-Sci. 2014, 34, 722–725. [Google Scholar] [CrossRef]
- Nasir, L.; Reid, S. Bovine papillomaviral gene expression in equine sarcoid tumours. Virus Res. 1999, 61, 171–175. [Google Scholar] [CrossRef]
- Yuan, Z.; Gallagher, A.; Gault, E.A.; Campo, M.S.; Nasir, L. Bovine papillomavirus infection in equine sarcoids and in bovine bladder cancers. Veter-J. 2007, 174, 599–604. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.; Gault, E.; Gobeil, P.; Nixon, C.; Campo, M.; Nasir, L. Establishment and characterization of equine fibroblast cell lines transformed in vivo and in vitro by BPV-1: Model systems for equine sarcoids. Virology 2008, 373, 352–361. [Google Scholar] [CrossRef] [Green Version]
- Bogaert, L.; Martens, A.; Kast, W.M.; Van Marck, E.; De Cock, H. Bovine papillomavirus DNA can be detected in keratinocytes of equine sarcoid tumors. Veter-Microbiol. 2010, 146, 269–275. [Google Scholar] [CrossRef]
- Gaynor, A.M.; Zhu, K.W.; Affolter, V.K.; Cruz, F.N.D.; Pesavento, P. Localization of Bovine Papillomavirus Nucleic Acid in Equine Sarcoids. Veter-Pathol. 2015, 53, 567–573. [Google Scholar] [CrossRef]
- Rector, A.; Van Ranst, M. Animal papillomaviruses. Virology 2013, 445, 213–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bravo, I.G.; Félez-Sánchez, M. Papillomaviruses. Evol. Med. Public Health 2015, 2015, 32–51. [Google Scholar] [CrossRef] [PubMed]
- Semik-Gurgul, E.; Gurgul, A.; Ząbek, T.; Koch, C.; Mählmann, K.; Ropka-Molik, K.; Bugno-Poniewierska, M. Transcriptome analysis of equine sarcoids. Veter-Comp. Oncol. 2016, 15, 1370–1381. [Google Scholar] [CrossRef] [PubMed]
- Pawlina-Tyszko, K.; Gurgul, A.; Szmatoła, T.; Koch, C.; Mählmann, K.; Witkowski, M.; Poniewierska, B.; Pawlina-Tyszko, K. Comprehensive characteristics of microRNA expression profile of equine sarcoids. Biochimie 2017, 137, 20–28. [Google Scholar] [CrossRef]
- Pawlina-Tyszko, K.; Gurgul, A.; Szmatoła, T.; Ropka-Molik, K.; Semik-Gurgul, E.; Klukowska-Rötzler, J.; Koch, C.; Mählmann, K.; Poniewierska, B. Genomic landscape of copy number variation and copy neutral loss of heterozygosity events in equine sarcoids reveals increased instability of the sarcoid genome. Biochimie 2017, 140, 122–132. [Google Scholar] [CrossRef]
- Unger, L.; Jagannathan, V.; Pacholewska, A.; Leeb, T.; Gerber, V. Differences in miRNA differential expression in whole blood between horses with sarcoid regression and progression. J. Veter-Intern. Med. 2018, 33, 241–250. [Google Scholar] [CrossRef]
- Unger, L.; Gerber, V.; Pacholewska, A.; Leeb, T.; Jagannathan, V. MicroRNA fingerprints in serum and whole blood of sarcoid-affected horses as potential non-invasive diagnostic biomarkers. Veter-Comp. Oncol. 2018, 17, 107–117. [Google Scholar] [CrossRef]
- Taylor, S.; Haldorson, G. A review of equine sarcoid. Equine Veter-Educ. 2012, 25, 210–216. [Google Scholar] [CrossRef] [Green Version]
- Hanahan, D.; Weinberg, R.A. TheHallmarks of Cancer. Cell 2000, 100, 57–70. [Google Scholar] [CrossRef] [Green Version]
- Lambert, A.W.; Pattabiraman, D.; Weinberg, R.A. Emerging Biological Principles of Metastasis. Cell 2017, 168, 670–691. [Google Scholar] [CrossRef] [Green Version]
- Tomasek, J.J.; Haaksma, C.J.; Eddy, R.J.; Vaughan, M. Fibroblast contraction occurs on release of tension in attached collagen lattices: Dependency on an organized actin cytoskeleton and serum. Anat. Rec. Adv. Integr. Anat. Evol. Boil. 1992, 232, 359–368. [Google Scholar] [CrossRef] [PubMed]
- Teifke, J.P.; Kidney, B.A.; Löhr, C.; Yager, J.A. Detection of papillomavirus-DNA in mesenchymal tumour cells and not in the hyperplastic epithelium of feline sarcoids. Veter-Dermatol. 2003, 14, 47–56. [Google Scholar] [CrossRef]
- Primer3. Available online: http://bioinfo.ut.ee/primer3-0.4.0 (accessed on 17 July 2017).
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, 45. [Google Scholar] [CrossRef] [PubMed]
- Anonymous. The R Project for Statistical Computing. Available online: http://www.r-project.org/ (accessed on 13 February 2012).
- Fife, C.M.; A McCarroll, J.; Kavallaris, M. Movers and shakers: Cell cytoskeleton in cancer metastasis. Br. J. Pharmacol. 2014, 171, 5507–5523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stengel, K.; Zheng, Y. Cdc42 in oncogenic transformation, invasion, and tumorigenesis. Cell. Signal. 2011, 23, 1415–1423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Y.; Kenney, S.R.; Muller, C.Y.; Adams, S.; Rutledge, T.; Romero, E.; Murray-Krezan, C.; Prekeris, R.; Sklar, L.A.; Hudson, L.G.; et al. R-Ketorolac Targets Cdc42 and Rac1 and Alters Ovarian Cancer Cell Behaviors Critical for Invasion and Metastasis. Mol. Cancer Ther. 2015, 14, 2215–2227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abreu-Velez, A.M.; Howard, M.S. Collagen IV in Normal Skin and in Pathological Processes. N. Am. J. Med. Sci. 2012, 4, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Vihinen, P.; Kähäri, V.-M. Matrix metalloproteinases in cancer: Prognostic markers and therapeutic targets. Int. J. Cancer 2002, 99, 157–166. [Google Scholar] [CrossRef]
- Yuan, Z.; Gobeil, P.A.; Campo, M.S.; Nasir, L. Equine sarcoid fibroblasts over-express matrix metalloproteinases and are invasive. Virology 2010, 396, 143–151. [Google Scholar] [CrossRef]
- Anania, M.C.; Sensi, M.; Radaelli, E.; Miranda, C.; Vizioli, M.G.; Pagliardini, S.; Favini, E.; Cleris, L.; Supino, R.; Formelli, F.; et al. TIMP3 regulates migration, invasion and in vivo tumorigenicity of thyroid tumor cells. Oncogene 2011, 30, 3011–3023. [Google Scholar] [CrossRef] [Green Version]
- Adissu, H.A.; McKerlie, C.; Di Grappa, M.; Waterhouse, P.; Xu, Q.; Fang, H.; Khokha, R.; Wood, G.A. Timp3 loss accelerates tumour invasion and increases prostate inflammation in a mouse model of prostate cancer. Prostate 2015, 75, 1831–1843. [Google Scholar] [CrossRef]
- Williams, H.C.; Martin, A.S.; Adamo, C.M.; Seidel-Rogol, B.; Pounkova, L.; Datla, S.R.; Lassègue, B.; Bear, J.E.; Griendling, K. Role of coronin 1B in PDGF-induced migration of vascular smooth muscle cells. Circ. Res. 2012, 111, 56–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, G.-Y.; Park, J.-H.; Kim, H.; Lim, H.-J.; Park, H.-Y. Coronin 1B serine 2 phosphorylation by p38α is critical for vascular endothelial growth factor-induced migration of human umbilical vein endothelial cells. Cell. Signal. 2016, 28, 1817–1825. [Google Scholar] [CrossRef] [PubMed]
- Priya, R.; Wee, K.; Budnar, S.; Gomez, G.A.; Yap, A.S.; Michael, M. Coronin 1B supports RhoA signaling at cell-cell junctions through Myosin II. Cell Cycle 2016, 15, 3033–3041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dave, J.M.; Bayless, K.J. Vimentin as an Integral Regulator of Cell Adhesion and Endothelial Sprouting. Microcirc. 2014, 21, 333–344. [Google Scholar] [CrossRef]
- Battaglia, R.A.; Delic, S.; Herrmann, H.; Snider, N.T. Vimentin on the move: New developments in cell migration. F1000Research 2018, 7, 1796. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Gene | Primer Sequence | Product Length (bp) |
|---|---|---|
| CDC42bpα | F:GCTCCATTCAAACGACCACA | 176 |
| R:AAGGATTTGCTGGCCACATC | ||
| CORO1b | F:AGATCGCCCGGTTCTACAAA | 179 |
| R:CAGGGAAATGAGGATGGGGT | ||
| MMP2 | F:TCCCACTTTGATGACGACGA | 182 |
| R:AAGTTGTAGGTGGTGGAGCA | ||
| TIMP3 | F:AAGATGCCCCATGTGCAGTA | 213 |
| R:TGCAGTTACAACCCAGGTGA | ||
| VIM | F:ACAAGTCCAAGTTTGCCGAC | 262 |
| R:CGCGCCATTTCTTTCCTTCAT | ||
| GAPDH | F:TCACCAGGGCTGCTTTTAAC | 156 |
| R:GCCTTTCCGTTGATGACAAG |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Podstawski, P.; Witarski, W.; Szmatoła, T.; Bugno-Poniewierska, M.; Ropka-Molik, K. Mobility and Invasion Related Gene Expression Patterns in Equine Sarcoid. Animals 2020, 10, 880. https://doi.org/10.3390/ani10050880
Podstawski P, Witarski W, Szmatoła T, Bugno-Poniewierska M, Ropka-Molik K. Mobility and Invasion Related Gene Expression Patterns in Equine Sarcoid. Animals. 2020; 10(5):880. https://doi.org/10.3390/ani10050880
Chicago/Turabian StylePodstawski, Przemysław, Wojciech Witarski, Tomasz Szmatoła, Monika Bugno-Poniewierska, and Katarzyna Ropka-Molik. 2020. "Mobility and Invasion Related Gene Expression Patterns in Equine Sarcoid" Animals 10, no. 5: 880. https://doi.org/10.3390/ani10050880