Mitochondrial DNA Survey Reveals the Lack of Accuracy in Maremmano Horse Studbook Records

 , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Samples
2.2. Pedigree Analysis
2.3. Reconstruction of Sire’s and Dam’s Lines
2.4. Mitochondrial DNA Control Region Analyses
3. Results
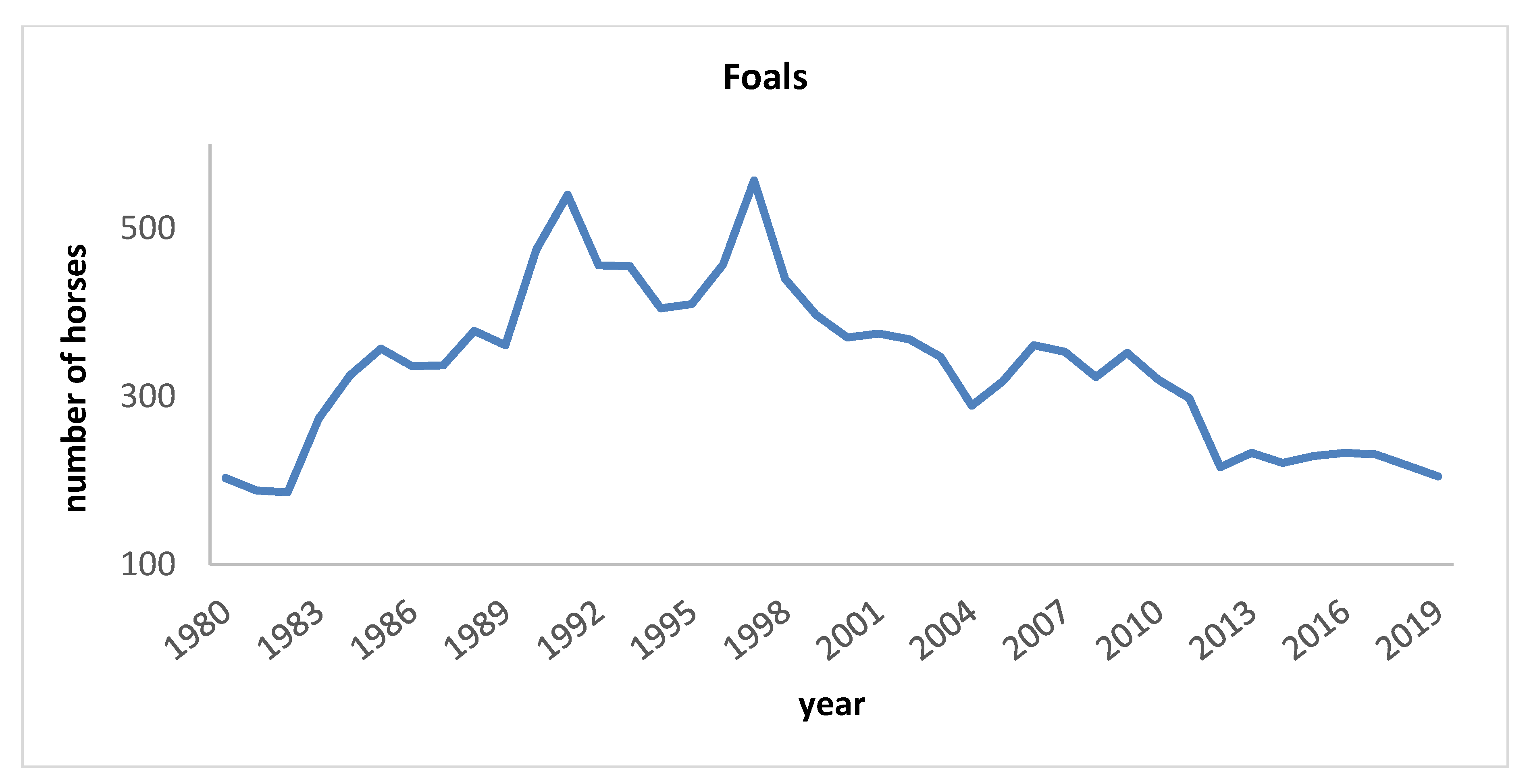
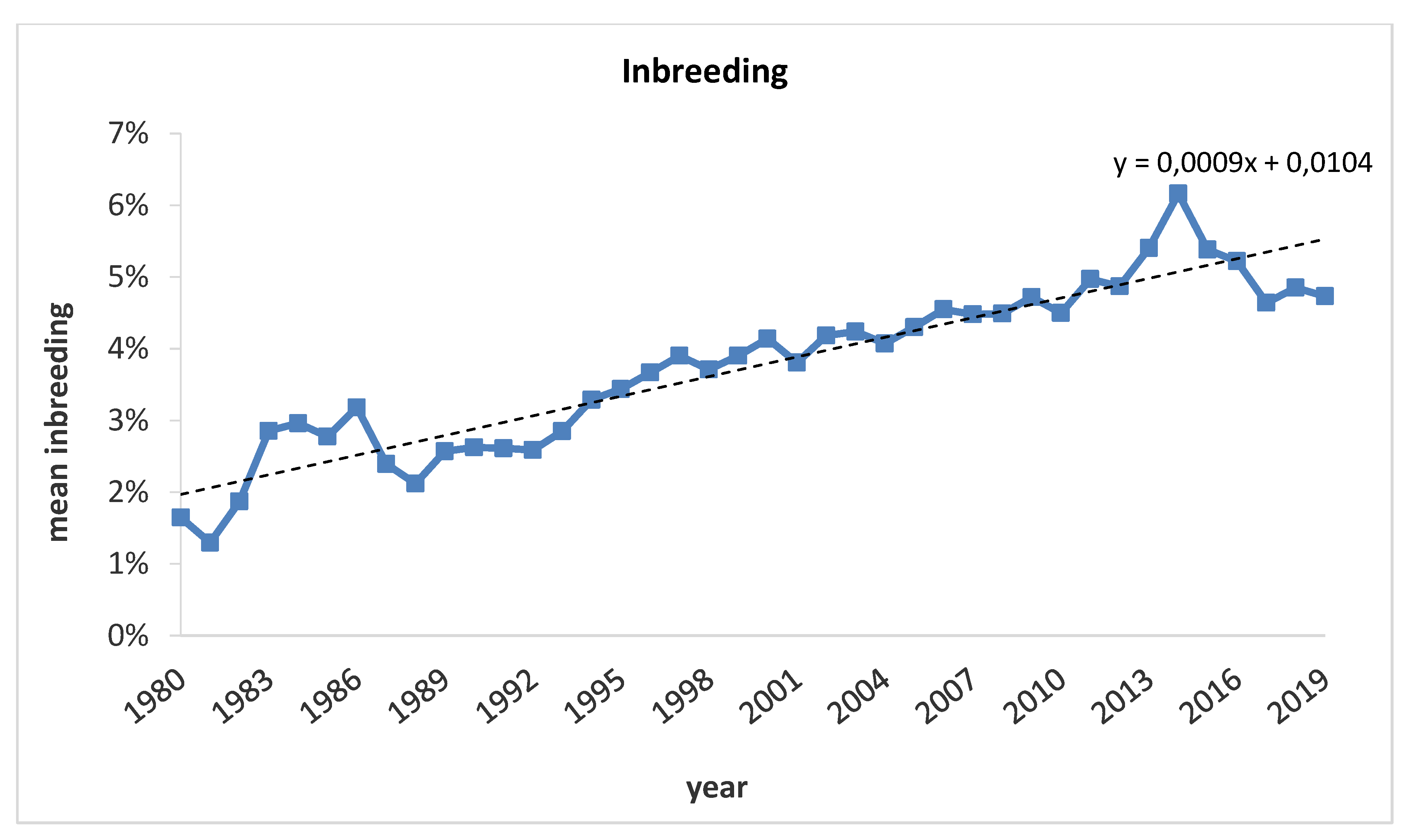
3.1. Pedigree Analysis
3.2. Reconstruction of Sire’s and Dam’s Lines
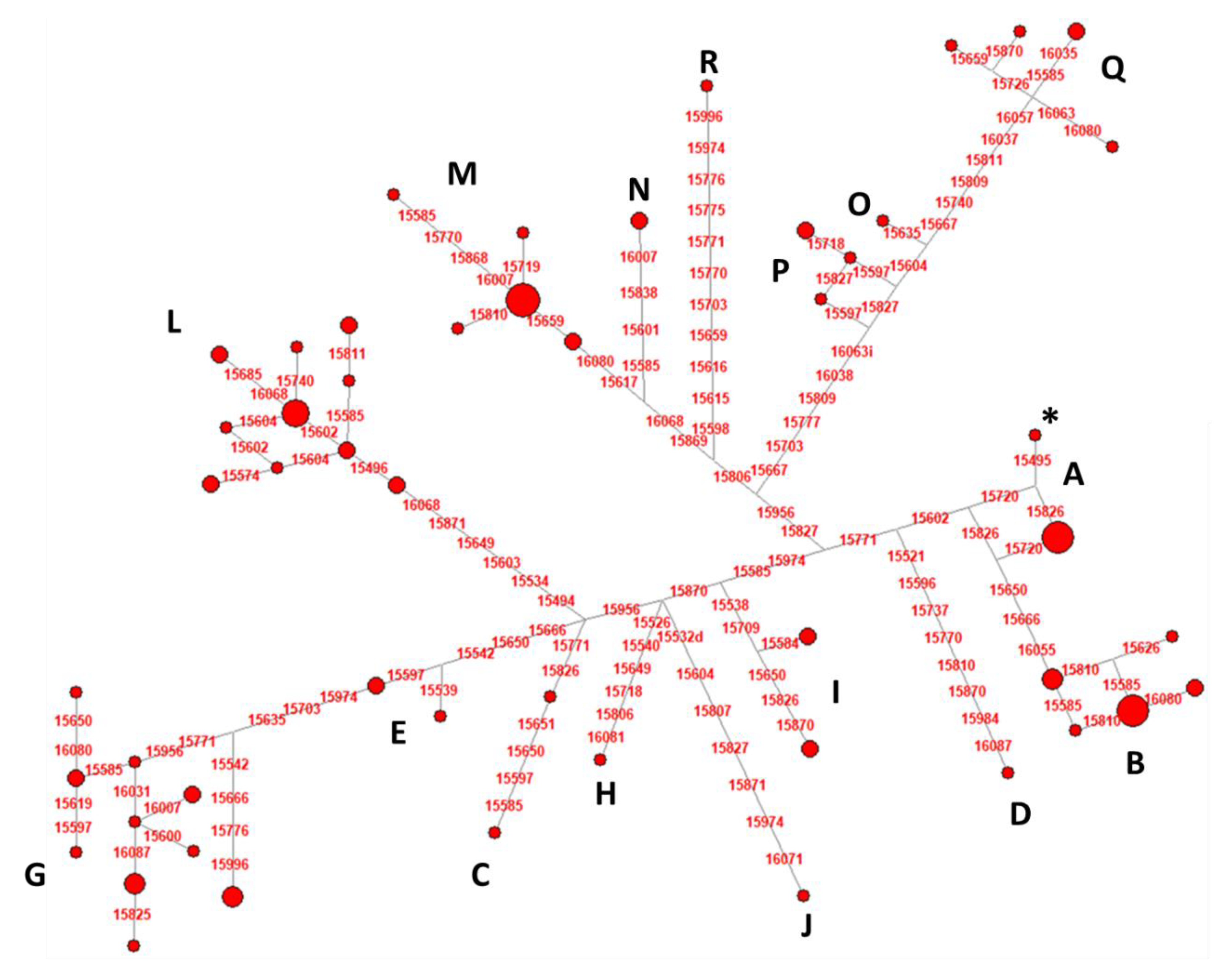
3.3. Mitochondrial DNA Control Region Analyses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Oldenbroek, K.; van der Waaij, L. Textbook Animal Breeding: Animal Breeding and Genetics for BSc Students; Centre for Genetic Resources and Animal Breeding and Genomics Group, Wageningen University and Research Centre: Wageningen, Netherlands, 2014. [Google Scholar]
- Giontella, A.; Pieramati, C.; Silvestrelli, M.; Sarti, F.M. Analysis of founders and performance test effects on an autochthonous horse population through pedigree analysis: Structure, genetic variability and inbreeding. Animal 2019, 13, 15–24. [Google Scholar] [CrossRef]
- Giontella, A.; Sarti, F.M.; Cardinali, I.; Giovannini, S.; Cherchi, R.; Lancioni, H.; Silvestrelli, M.; Pieramati, C. Genetic variability and population structure in Sardinian Anglo-Arab horse. Animals. under review.
- Vostrá-Vydrová, H.; Vostrý, L.; Hofmanová, B.; Krupa, E.; Zavadilová, L. Pedigree analysis of the endangered Old Kladruber horse population. Livest. Sci. 2016, 185, 17–23. [Google Scholar] [CrossRef]
- Ablondi, M.; Vasini, M.; Beretti, V.; Superchi, P.; Sabbioni, A. Exploring genetic diversity in an Italian horse native breed to develop strategies for preservation and management. J. Anim. Breed. Genet. 2018, 135, 450–459. [Google Scholar] [CrossRef]
- Fabbri, M.C.; Gonçalves de Rezende, M.P.; Dadousis, C.; Biffani, S.; Negrini, R.; Souza Carneiro, P.L.; Bozzi, R. Population Structure and Genetic Diversity of Italian Beef Breeds as a Tool for Planning Conservation and Selection Strategies. Animals (Basel) 2019, 9, 880. [Google Scholar] [CrossRef] [Green Version]
- Bodo, I.; Lawrence, A.; Langlois, B. Conservation Genetics of Endangered Horse Breeds; Wageningen Academic Publishers: Wageningen, Netherlands, 2005; pp. 155–162. [Google Scholar]
- Bömcke, E.; Gengler, N.; Cothran, E.G. Genetic variability in the Skyros pony and its relationship with other Greek and foreign horse breeds. Genet. Mol. Biol. 2011, 34, 68–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasler, H.; Flury, C.; Menet, S.; Haase, B.; Leeb, T.; Simianer, H.; Poncet, P.A.; Rieder, S. Genetic diversity in an indigenous horse breed: Implications for mating strategies and the control of future inbreeding. J. Anim. Breed. Genet. 2011, 128, 394–406. [Google Scholar] [CrossRef] [PubMed]
- Álvarez, I.; Fernández, I.; Lorenzo, L.; Payeras, L.; Cuervo, M.; Goyache, F. Founder and present maternal diversity in two endangered Spanish horse breeds assessed via pedigree and mitochondrial DNA information. J. Anim. Breed. Genet. 2012, 129, 271–279. [Google Scholar] [CrossRef]
- Głażewska, I.; Gralak, B.; Naczk, A.M.; Prusak, B. Genetic diversity and population structure of Polish Arabian horses assessed through breeding and microsatellite data. Anim. Sci. J. 2012, 89, 735–742. [Google Scholar] [CrossRef]
- Bowling, A.T.; Del Valle, A.; Bowling, M. A pedigree-Based study of mitochondrial D-loop DNA sequence variation among Arabian horses. Anim. Genet. 2000, 31, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, E.W.; Bradley, D.G.; Al-Barody, M.; Ertugrul, O.; Splan, R.K.; Zakharov, I.; Cunningham, E.P. History and integrity of thoroughbred dam lines revealed in equine mtDNA variation. Anim Genet. 2002, 33, 287–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kavar, T.; Bremm, G.; Habe, F.; Solkner, J.; Dovc, P. History of Lipizzan horse maternal lines as revealed by mtDNA analysis. Genet. Sel. Evol. 2002, 34, 635–648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Głażewska, I.; Wysocka, A.; Gralak, B.; Sell, J. A new view on dam lines in Polish Arabian horses based on mtDNA analysis. Genet. Sel. Evol. 2007, 39, 609–619. [Google Scholar] [CrossRef]
- Čačić, M.; Čubrić Čurik, V.; Baban, M.; Barać, Z.; Čurik, I. Use of mitochondrial DNA analyses in verification of the Lipizzan horse pedigree. Agric. Conspec. Sci. 2011, 76, 365–368. [Google Scholar]
- Bower, M.A.; McGivney, B.A.; Campana, M.G.; Gu, J.; Andersson, L.S.; Barrett, E.; Davis, C.R.; Mikko, S.; Stock, F.; Voronkova, V.; et al. The genetic origin and history of speed in the Thoroughbred racehorse. Nat. Commun. 2012, 3, 643. [Google Scholar] [CrossRef] [PubMed]
- Jansen, T.; Forster, P.; Levine, M.A.; Oelke, H.; Hurles, M.; Renfrew, C.; Weber, J.; Olek, K. Mitochondrial DNA and the origins of the domestic horse. Proc. Natl. Acad. Sci. USA 2002, 99, 10905–10910. [Google Scholar] [CrossRef] [Green Version]
- McGahern, A.; Bower, M.A.; Edwards, C.J.; Brophy, P.O.; Sulimova, G.; Zakharov, I.; Vizuete-Forster, M.; Levine, M.; Li, S.; MacHugh, D.E.; et al. Evidence for biogeographic patterning of mitochondrial DNA sequences in Eastern horse populations. Anim. Genet. 2006, 37, 494–497. [Google Scholar] [CrossRef]
- Lei, C.Z.; Su, R.; Bower, M.A.; Edwards, C.J.; Wang, X.B.; Weining, S.; Liu, L.; Xie, W.M.; Li, F.; Liu, R.Y.; et al. Multiple maternal origins of native modern and ancient horse populations in China. Anim. Genet. 2009, 40, 933–944. [Google Scholar] [CrossRef]
- Achilli, A.; Olivieri, A.; Soares, P.; Lancioni, H.; Hooshiar Kashani, B.; Perego, U.A.; Nergadze, S.G.; Carossa, V.; Santagostino, M.; Capomaccio, S.; et al. Mitochondrial genomes from modern horses reveal the major haplogroups that underwent domestication. Proc. Natl. Acad. Sci. USA 2012, 109, 2449–2454. [Google Scholar] [CrossRef] [Green Version]
- Khanshour, A.M.; Cothran, E.G. Maternal phylogenetic relationships and genetic variation among Arabian horse populations using whole mitochondrial DNA D-loop sequencing. BMC Genet. 2013, 14, 83. [Google Scholar] [CrossRef] [Green Version]
- Bonavolontà, G.; Silvestrelli, M. Il Maremmano; Bramante edizioni Equestri: Milano, Italy, 1989. [Google Scholar]
- Pieramati, C.; Sarti, F.M.; Silvestrelli, M.; Giontella, A. Optimization of the performance test length for the Sella Italiano stallion. Ital. J. Anim. Sci. 2013, 12, e67. [Google Scholar] [CrossRef]
- Cappelli, K.; Capomaccio, S.; Giontella, A.; Nardone, A.; Silvestrelli, M. Genetic and phenotypic characterization of the Maremmano horse stud in Castelporziano. Rend. Fis.Acc. Lincei 2015, 26, 545–552. [Google Scholar] [CrossRef]
- Petersen, J.L.; Mickelson, J.R.; Cothran, E.G.; Andersson, L.S.; Axelsson, J.; Bailey, E.; Bannasch, D.; Binns, M.M.; Borges, A.S.; Brama, P.; et al. Diversity in the Modern Horse Illustrated from Genome-Wide SNP Data. PLoS Genet. 2013, 8, e54997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardinali, I.; Lancioni, H.; Giontella, A.; Capodiferro, M.R.; Capomaccio, S.; Buttazzoni, L.; Biggio, G.P.; Cherchi, R.; Albertini, E.; Olivieri, A.; et al. An Overview of Ten Italian Horse Breeds through Mitochondrial DNA. PLoS ONE 2016, 11, e0153004. [Google Scholar] [CrossRef] [Green Version]
- Kinghorn, B.P. Pedigree Viewer-A graphical utility for browsing pedigreed data sets. In Proceedings of the 5th World Congress on Genetics Applied to Livestock Production, Guelph, ON, Canada, 7–12 August 1994; Volume 22, pp. 85–86. [Google Scholar]
- Gutierrez, J.P.; Goyache, F. A note on ENDOG: A computer program for analysing pedigree information. J. Anim. Breed. Genet. 2005, 122, 357–360. [Google Scholar] [CrossRef] [Green Version]
- Nei, M. Molecular Evolutionary Genetics; Columbia University Press: New York, USA, 1987. [Google Scholar]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giontella, A.; Cardinali, I.; Lancioni, H.; Giovannini, S.; Pieramati, C.; Silvestrelli, M.; Sarti, F.M. Mitochondrial DNA Survey Reveals the Lack of Accuracy in Maremmano Horse Studbook Records. Animals 2020, 10, 839. https://doi.org/10.3390/ani10050839
Giontella A, Cardinali I, Lancioni H, Giovannini S, Pieramati C, Silvestrelli M, Sarti FM. Mitochondrial DNA Survey Reveals the Lack of Accuracy in Maremmano Horse Studbook Records. Animals. 2020; 10(5):839. https://doi.org/10.3390/ani10050839
Chicago/Turabian StyleGiontella, Andrea, Irene Cardinali, Hovirag Lancioni, Samira Giovannini, Camillo Pieramati, Maurizio Silvestrelli, and Francesca Maria Sarti. 2020. "Mitochondrial DNA Survey Reveals the Lack of Accuracy in Maremmano Horse Studbook Records" Animals 10, no. 5: 839. https://doi.org/10.3390/ani10050839