Influence of Housing Systems on Physical, Emotional, and Cognitive Functions with Aging in DBA/2CrSlc Mice


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Housing Systems and Cages (IVC type vs. Chamber Type)
2.3. Animals and Housing Conditions
2.4. Food Consumption and Body Weight
2.5. Measurement of Rectal Core Temperature
2.6. Changes in Physical Performances with Aging (Tests 1–5)
2.6.1. Balancing Ability on an Acryl Rod (Test 1)
2.6.2. Horizontal Bar Test (Test 2)
- Score 5, crossing on the bar with two forefeet and two hindfeet;
- Score 4, not crossing the bar, but clinging with two forefeet and two hindfeet;
- Score 3, clinging with two forefeet and one hindfoot;
- Score 2, hanging with two forefeet for more than 10 s and falling off the bar;
- Score 1, hanging with two forefeet for under 10 s and falling off the bar.
2.6.3. Wire Hanging Test (Test 3)
2.6.4. Footprint Test (Test 4)
2.6.5. Locomotor Activities (Test 5)
2.7. Changes in Anxiety-like Behaviors with Aging (Test 6 and Test 7)
2.7.1. Black and White Box Test (Test 6)
2.7.2. Open Field Test (Test 7)
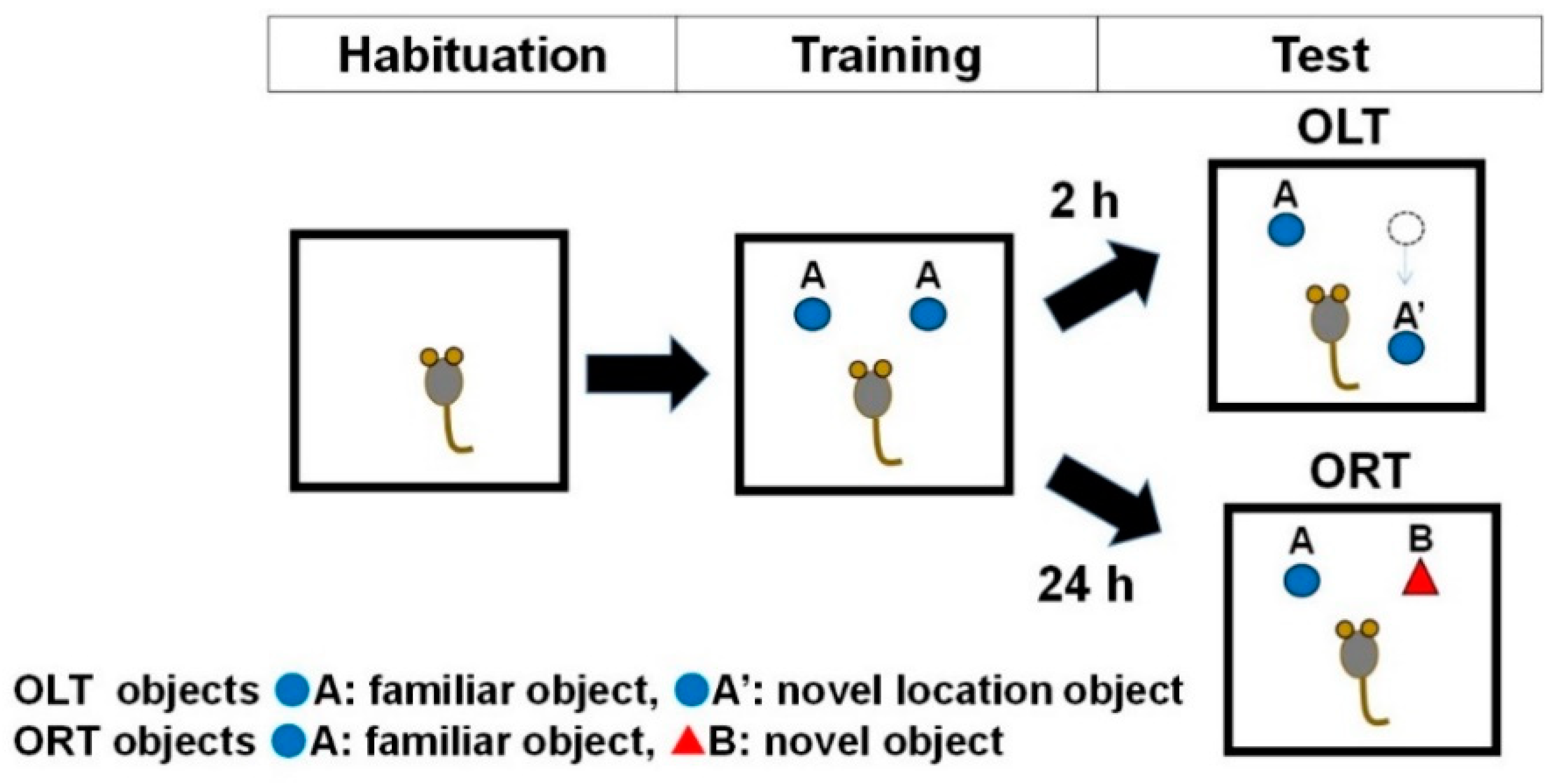
2.8. Changes in Spatial Recognition (Test 8, Short-Term Location Memory) and Object Recognition (Test 9, Long-Term Object Memory) with Aging
2.8.1. Paradigm of ORT and OLT (Test 8 and Test 9)
2.8.2. Data analysis of ORT and OLT (Test 8 and Test 9)
2.9. Changes in Intestinal Microbiome
2.10. Degree of Visceroptosis
2.11. Statistical Analyses
3. Results
3.1. Food Consumption and Body Weight with Aging
3.2. Changes in Rectal Core Temperature with Aging
3.3. Changes in Physical Performances in chamber and IVC groups with Aging (Tests 1–5)
3.3.1. Balancing Ability on an Acryl Rod (Test 1)
3.3.2. Horizontal Bar Test (Test 2)
3.3.3. Wire Hanging Test (Test 3)
3.3.4. Footprint Test (Test 4)
3.3.5. Locomotor Activities (Test 5)
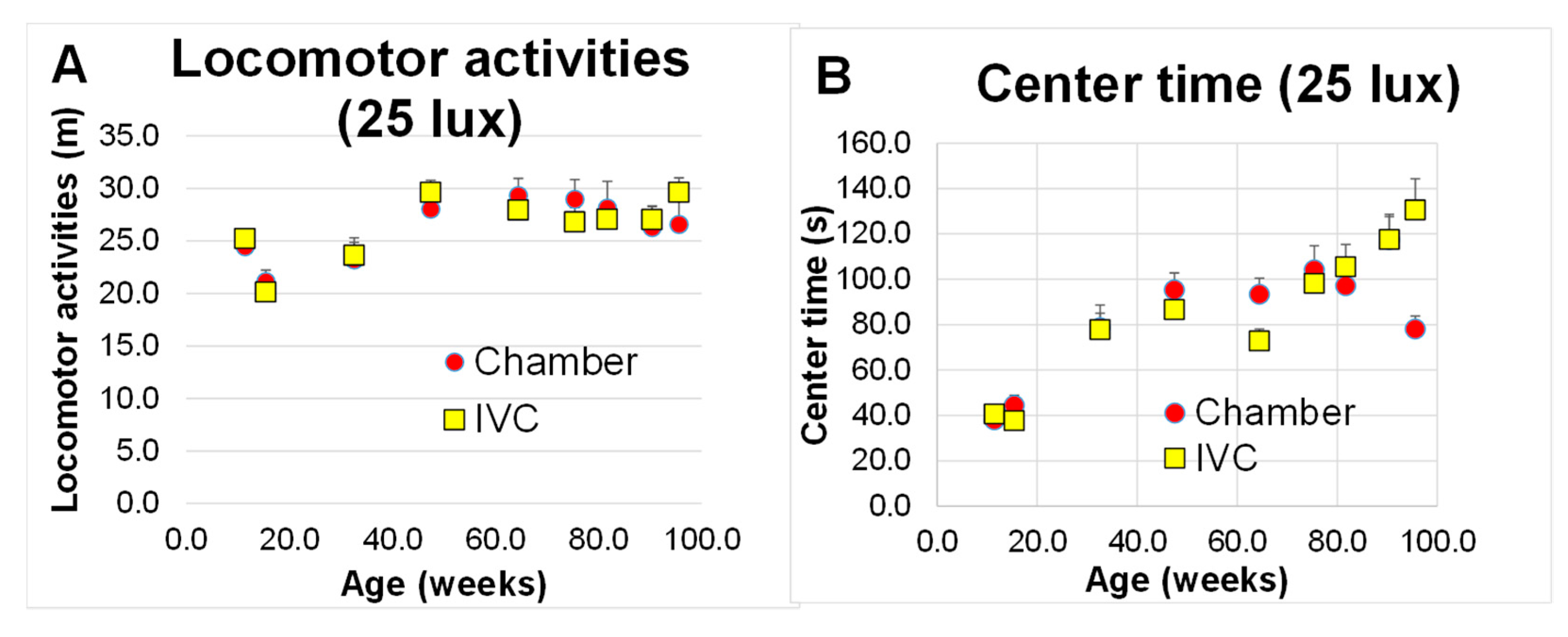
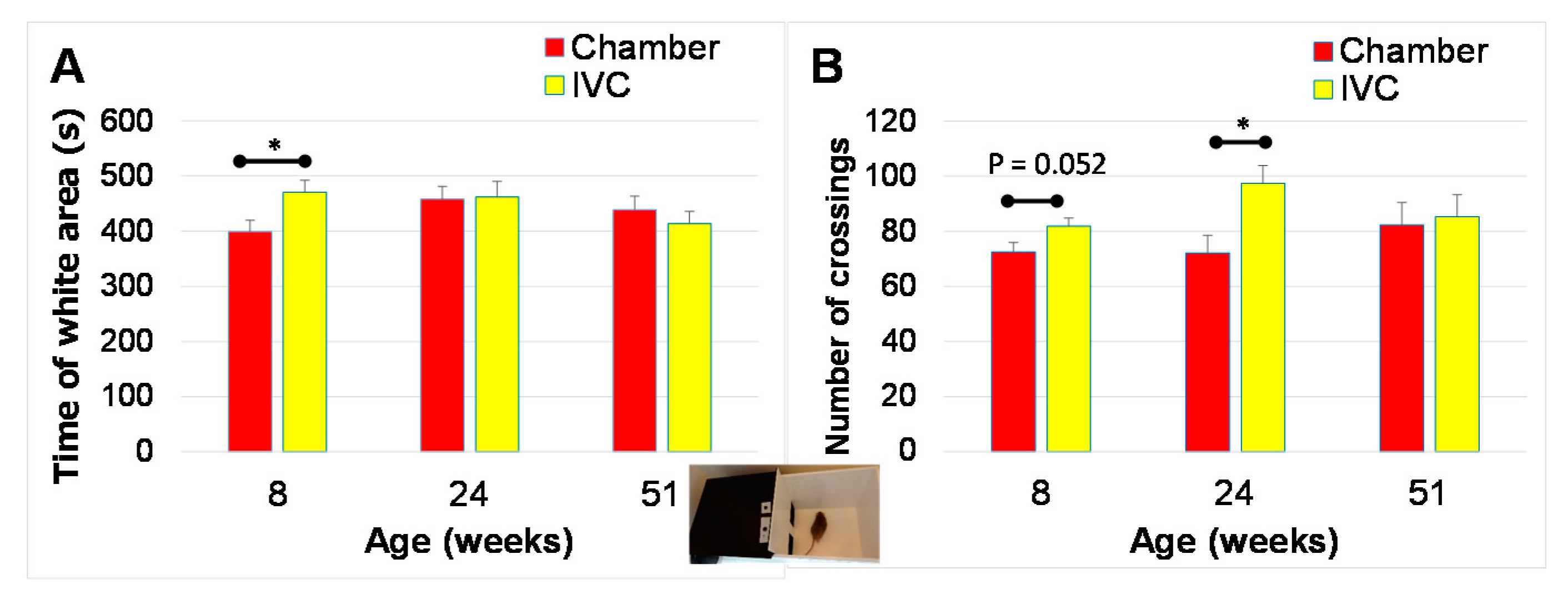
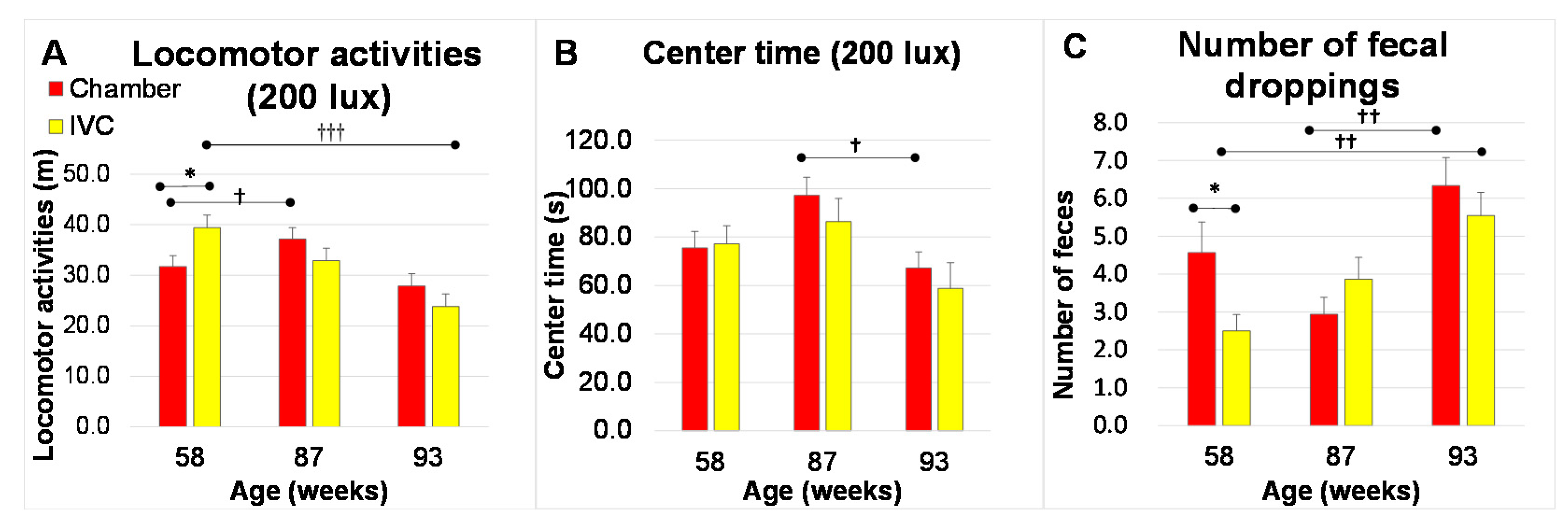
3.4. Changes in Anxiety-like Behaviors with Aging (Test 6 and Test 7)
3.4.1. Black and White Box Test (Test 6)
3.4.2. Open Field Test (Test 7)
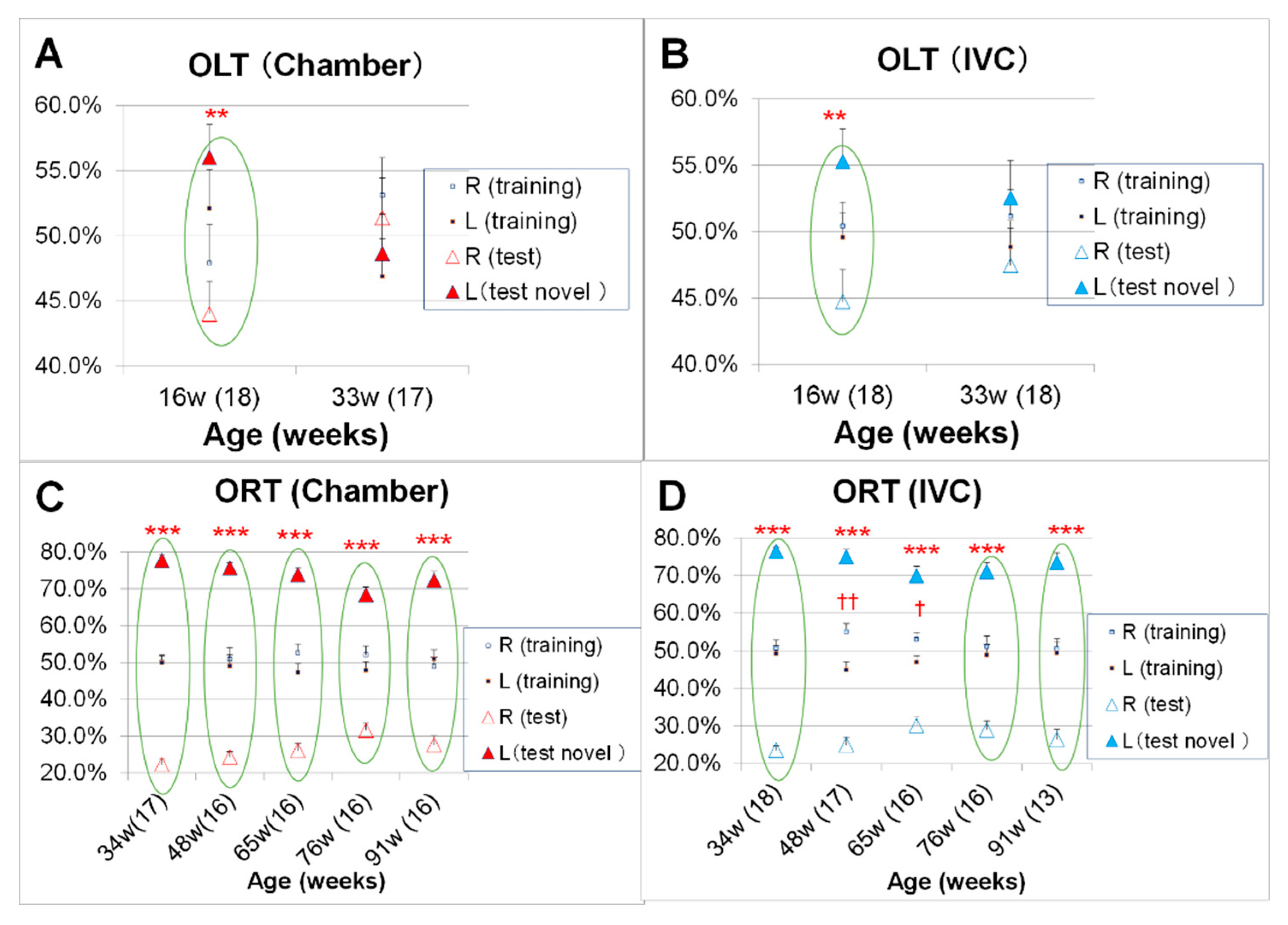
3.5. Changes in Spatial Recognition (Test 8, Short-Term Location Memory) and Object Recognition (Test 9, Long-Term Object Memory) with Aging
3.6. Changes in Intestinal Microbiome with Aging
3.7. Changes in Other Behaviors and Body Features with Aging
3.7.1. Champing Behavior to Slice Papers of Bedding in Cage and Dishcloth Gourd for Enrichment
3.7.2. Changes in the Bulge of the Rump at the Age of 94 W
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tractenberg, S.G.; Levandowski, M.L.; Azeredo, L.A.; Orso, R.; Roithmann, L.G.; Hoffmann, E.S.; Brenhouse, H.; Grassi-Oliveira, R. An overview of maternal separation effects on behavioural outcomes in mice: Evidence from a four-stage methodological systematic review. Neurosci. Biobehav. Rev. 2016, 68, 489–503. [Google Scholar] [CrossRef] [PubMed]
- Niwa, M.; Jaaro-Peled, H.; Tankou, S.; Seshadri, S.; Hikida, T.; Matsumoto, Y.; Cascella, N.G.; Kano, S.; Ozaki, N.; Nabeshima, T.; et al. Adolescent stress-induced epigenetic control of dopaminergic neurons via glucocorticoids. Science 2013, 339, 335–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, A.; Chung, J.; Zhang, S.; Zhang, H.; Grossman, Y.; Aleyasin, H.; Flanigan, M.E.; Pfau, M.L.; Menard, C.; Dumitriu, D.; et al. Establishment of a repeated social defeat stress model in female mice. Sci. Rep. 2017, 7, 12838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iñiguez, S.D.; Riggs, L.M.; Nieto, S.J.; Dayrit, G.; Zamora, N.Z.; Shawhan, K.L.; Cruz, C.; Warren, B.L. Social defeat stress induces a depression-like phenotype in adolescent male c57BL/6 mice. Stress 2014, 17, 247–255. [Google Scholar] [CrossRef] [Green Version]
- Koseki, T.; Mouri, A.; Mamiya, T.; Aoyama, Y.; Toriumi, K.; Suzuki, S.; Nakajima, A.; Yamada, T.; Nagai, T.; Nabeshima, T. Exposure to enriched environments during adolescence prevents abnormal behaviours associated with histone deacetylation in phencyclidine-treated mice. Int. J. Neuropsychopharmacol. 2012, 15, 1489–1501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bayne, K. Environmental enrichment and mouse models: Current perspectives. Animal Model. Exp. Med. 2018, 1, 82–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailoo, J.D.; Murphy, E.; Boada-Saña, M.; Varholick, J.A.; Hintze, S.; Baussière, C.; Hahn, K.C.; Göpfert, C.; Palme, R.; Voelkl, B.; et al. Effects of cage enrichment on behavior, welfare and outcome variability in female mice. Front. Behav. Neurosci. 2017, 12, 232. [Google Scholar] [CrossRef]
- Martinez-Cue, C.; Baamonde, C.; Lumbreras, M.; Paz, J.; Davisson, M.T.; Schmidt, C.; Dierssen, M.; Flórez, J. Differential effects of environmental enrichment on behavior and learning of male and female Ts65Dn mice, a model for Down syndrome. Behav. Brain Res. 2002, 134, 185–200. [Google Scholar] [CrossRef]
- Nilsson, M.; Perfilieva, E.; Johansson, U.; Orwar, O.; Eriksson, P.S. Enriched environment increases neurogenesis in the adult rat dentate gyrus and improves spatial memory. J. Neurobiol. 1999, 39, 569–578. [Google Scholar] [CrossRef]
- Yamashita, Y.; Kawai, N.; Ueno, O.; Matsumoto, Y.; Oohashi, T.; Honda, M. Induction of prolonged natural lifespans in mice exposed to acoustic environmental enrichment. Sci. Rep. 2018, 8, 7909. [Google Scholar] [CrossRef]
- Braden, G.C.; Rasmussen, S.; Monette, S.; Tolwani, R.J. Effects of rearing configuration on maternal and weanling behavior in laboratory mice. J. Am. Assoc. Lab. Anim. Sci. 2017, 56, 369–376. [Google Scholar]
- Zidar, J.; Weber, E.M.; Ewaldsson, B.; Tjäder, S.; Lilja, J.; Mount, J.; Svensson, C.; Svensk, E.; Udén, E.; Törnqvist, E. Group and single housing of male mice: Collected experiences from research facilities in Sweden. Animals 2019, 9, 1010. [Google Scholar] [CrossRef] [Green Version]
- Freymann, J.; Tsai, P.; Stelzer, H.; Hackbarth, H. The amount of cage bedding preferred by female BALB/c and C57BL/6 mice. Lab. Anim. (NY) 2015, 44, 17–22. [Google Scholar] [CrossRef]
- Spangenberg, E.; Wallenbeck, A.; Eklöf, A.-C.; Carlstedt-Duke, J.; Tjäder, S. Housing rearing mice in three different IVC systems: Maternal performance and pup development. Lab. Anim. 2014, 48, 193–206. [Google Scholar] [CrossRef] [Green Version]
- Yadav, S.; Yadav, I.; Pratap, K.; Tiwari, P.K.; Singh, V.P. Reproductive performance of genetically engineered mice housed in different housing systems. Lab. Anim. Res. 2017, 33, 68–75. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.; Brielmeier, M. Environmental samples make soiled bedding sentinels dispensable for hygienic monitoring of IVC-reared mouse colonies. Lab. Anim. 2018, 52, 233–239. [Google Scholar] [CrossRef]
- Tsai, P.P.; Oppermann, D.; Stelzer, H.D.; Mähler, M.; Hackbarth, H. The effects of different rack systems on the rearing performance of DBA/2 mice. Lab. Anim. 2003, 37, 44–53. [Google Scholar] [CrossRef] [PubMed]
- Feistenauer, S.; Sander, I.; Schmidt, J.; Zahradnik, E.; Raulf, M.; Brielmeier, M. Influence of 5 different caging types and the use of cage-changing stations on mouse allergen exposure. J. Am. Assoc. Lab. Anim. Sci. 2014, 53, 356–363. [Google Scholar] [PubMed]
- Shimizu, C.; Oki, Y.; Mitani, Y.; Nakamura, T.; Nabeshima, T. Factors affecting ethanol-induced conditioned place preference and locomotor sensitization in mice. Biol. Pharm. Bull. 2015, 38, 1935–1945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimizu, C.; Mitani, Y.; Tsuchiya, Y.; Nabeshima, T. Effects of oral calcium dosage and timing on ethanol-induced sensitization of locomotion in DBA/2 mice. Biol. Pharm. Bull. 2018, 41, 1049–1061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mantsch, J.R.; Weyer, A.; Vranjkovic, O.; Beyer, C.E.; Baker, D.A.; Caretta, H. Involvement of noradrenergic neurotransmission in the stress- but not cocaine-induced reinstatement of extinguished cocaine-induced conditioned place preference in mice: Role for β-2 adrenergic receptors. Neuropsychopharmacology 2010, 35, 2165–2178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zernig, G.; Pinheiro, B.S. Dyadic social interaction inhibits cocaine-conditioned place preference and the associated activation of the accumbens corridor. Behav. Pharmacol. 2015, 26, 580–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamada, K.; Nagai, T.; Nabeshima, T. Drug dependence, synaptic plasticity, and tissue plasminogen activator. J. Pharmacol. Sci. 2005, 97, 157–161. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, A.; Yamane, T.; Imaizumi, M.; Fushiki, T. Integration of orosensory and postingestive stimuli for the control of excessive fat intake in mice. Nutrition 2003, 19, 36–40. [Google Scholar] [CrossRef]
- Cason, A.M.; Smith, R.J.; Tahsili-Fahadan, P.; Moorman, D.E.; Sartor, G.C.; Aston-Jones, G. Role of orexin/hypocretin in reward-seeking and addiction: Implications for obesity. Physiol. Behav. 2010, 100, 419–428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toepper, M. Dissociating normal aging from Alzheimer’s disease: A view from cognitive neuroscience. J. Alzheimers Dis. 2017, 57, 331–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishibashi, H. Locomotive syndrome in Japan. Osteoporos. Sarcopenia 2018, 4, 86–94. [Google Scholar] [CrossRef]
- van Hagen, B.T.; van Goethem, N.P.; Lagatta, D.C.; Prickaerts, J. The object pattern separation (OPS) task: A behavioral paradigm derived from the object recognition task. Behav. Brain Res. 2015, 285, 44–52. [Google Scholar] [CrossRef]
- Li, S.; Wang, C.; Wang, W.; Dong, H.; Hou, P.; Tang, Y. Chronic mild stress impairs cognition in mice: From brain homeostasis to behavior. Life Sci. 2008, 82, 934–942. [Google Scholar] [CrossRef]
- Takeda, T.; Hosokawa, M.; Takeshita, S.; Irino, M.; Higuchi, K.; Matushita, T.; Tomita, Y.; Yasuhira, K.; Hamamoto, H.; Shimizu, K.; et al. A new murine model of accelerated senescence. Mech. Ageing Dev. 1981, 17, 183–194. [Google Scholar] [CrossRef]
- Akiguchi, I.; Pallàs, M.; Budka, H.; Akiyama, H.; Ueno, M.; Han, J.; Yagi, H.; Nishikawa, T.; Chiba, Y.; Sugiyama, H.; et al. SAMP8 mice as a neuropathological model of accelerated brain aging and dementia: Toshio Takeda’s legacy and future directions. Neuropathology 2017, 37, 293–305. [Google Scholar] [CrossRef] [Green Version]
- Shimizu, C.; Oki, Y.; Mitani, Y.; Tsuchiya, Y.; Nabeshima, T. Moderate-dose regular lifelong alcohol intake changes the intestinal flora, protects against aging, and keeps spatial memory in the senescence-accelerated mouse prone 8 (SAMP8) model. J. Pharm. Pharm. Sci. 2016, 19, 430–447. [Google Scholar] [CrossRef]
- Shimizu, C.; Wakita, Y.; Inoue, T.; Hiramitsu, M.; Okada, M.; Mitani, Y.; Segawa, S.; Tsuchiya, Y.; Nabeshima, T. Effects of lifelong intake of lemon polyphenols on aging and intestinal microbiome in the senescence-accelerated mouse prone 1 (SAMP1). Sci. Rep. 2019, 9, 3671. [Google Scholar] [CrossRef] [Green Version]
- Shimizu, C.; Wakita, Y.; Kihara, M.; Kobayashi, N.; Tsuchiya, Y.; Nabeshima, T. Association of lifelong intake of barley diet with healthy aging: Changes in physical and cognitive functions and intestinal microbiome in senescence-accelerated mouse-prone 8 (SAMP8). Nutrients 2019, 11, E1770. [Google Scholar] [CrossRef] [Green Version]
- Abreu-Vieira, G.; Xiao, C.; Gavrilova, O.; Reitman, M.L. Integration of body temperature into the analysis of energy expenditure in the mouse. Mol. Metab. 2015, 4, 461–470. [Google Scholar] [CrossRef]
- Ludvik, B. Weight loss by increasing energy consumption and thermogenesis. Acta Med. Austriaca 1998, 25, 136–137. [Google Scholar]
- Shiomi, T.; Mori, M. Maintenance of hypothermia in mice. Exp. Animals 1968, 17, 97–102. (In Japanese) [Google Scholar] [CrossRef] [Green Version]
- Tai, C.Y.; Nabeshima, T.; Sivam, S.; Ho, I.K. Effects of acute and continuous administration of pentobarbital on phencyclidine response. Res. Comm. Sub. Abuse 1981, 2, 239–252. [Google Scholar]
- Gleitz, H.F.C.; O’Leary, C.; Holley, R.J.; Bigger, B.W. Identification of age-dependent motor and neuropsychological behavioural abnormalities in a mouse model of Mucopolysaccharidosis Type II. PLoS ONE 2017, 12, e0172435. [Google Scholar] [CrossRef]
- Garneau, A.P.; Marcoux, A.A.; Noël, M.; Frenette-Cotton, R.; Drolet, M.C.; Couet, J.; Larivière, R.; Isenring, P. Ablation of potassium-chloride cotransporter type 3 (Kcc3) in mouse causes multiple cardiovascular defects and isosmotic polyuria. PLoS ONE 2016, 11, e0154398. [Google Scholar] [CrossRef]
- Blizzard, C.A.; Southam, K.A.; Dawkins, E.; Lewis, K.E.; King, A.E.; Clark, J.A.; Dickson, T.C. Identifying the primary site of pathogenesis in amyotrophic lateral sclerosis—vulnerability of lower motor neurons to proximal excitotoxicity. Dis. Model. Mech. 2015, 8, 215–224. [Google Scholar] [CrossRef] [Green Version]
- Sanchez, C. Acute stress enhances anxiolytic-like drug responses of mice tested in a black and white test box. Eur. Neuropsychopharmacol. 1997, 7, 283–288. [Google Scholar] [CrossRef]
- Kraeuter, A.K.; Guest, P.C.; Sarnyai, Z. The Open field test for measuring locomotor activity and anxiety-like behavior. Methods Mol. Biol. 2019, 1916, 99–103. [Google Scholar] [CrossRef]
- Murai, T.; Okuda, S.; Tanaka, T.; Ohta, H. Characteristics of object location memory in mice: Behavioral and pharmacological studies. Physiol. Behav. 2007, 90, 116–124. [Google Scholar] [CrossRef]
- Mitsuoka, T. Intestinal flora and aging. Nutr. Rev. 1992, 50, 438–446. [Google Scholar] [CrossRef]
- Allen, J.M.; Berg Miller, M.E.; Pence, B.D.; Whitlock, K.; Nehra, V.; Gaskins, H.R.; White, B.A.; Fryer, J.D.; Wood, J.A. Voluntary and forced exercise differentially alters the gut microbiome in C57BL/6J mice. J. Appl. Physiol.(1985) 2015, 18, 1059–1066. [Google Scholar] [CrossRef] [Green Version]
- Matsuki, T.; Watanabe, K.; Fujimoto, J.; Kado, Y.; Takada, T.; Matsumoto, K.; Tanaka, R. Quantitative PCR with 16S rRNA-gene-targeted species-specific primers for analysis of human intestinal bifidobacteria. Appl. Environ. Microbiol. 2004, 70, 167–173. [Google Scholar] [CrossRef] [Green Version]
- Lozupone, C.; Lladser, M.E.; Knights, D.; Stombaugh, J.; Knight, R. UniFrac: An effective distance metric for microbial community comparison. ISME J. 2011, 5, 169–172. [Google Scholar] [CrossRef] [Green Version]
- Jung, A.P.; Curtis, T.S.; Turner, M.J.; Lightfoot, J.T. Physical activity and food consumption in high- and low-active inbred mouse strains. Med. Sci. Sports. Exerc. 2010, 42, 1826–1833. [Google Scholar] [CrossRef] [Green Version]
- Talan, M. Body temperature of C57BL/6J mice with age. Exp. Gerontol. 1984, 19, 25–29. [Google Scholar] [CrossRef]
- Takao, K.; Shoji, H.; Hattori, S.; Miyakawa, T. Cohort removal induces changes in body temperature, pain sensitivity, and anxiety-like behavior. Front. Behav. Neurosci. 2016, 10, 99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuzuki, T.; Yoshihara, T.; Ichinoseki-Sekine, N.; Kakigi, R.; Takamine, Y.; Kobayashi, H.; Naito, H. Body temperature elevation during exercise is essential for activating the Akt signaling pathway in the skeletal muscle of type 2 diabetic rats. PLoS ONE 2018, 13, e0205456. [Google Scholar] [CrossRef] [PubMed]
- Tieland, M.; Trouwborst, I.; Clark, B.C. Skeletal muscle performance and ageing. J. Cachexia Sarcopenia Muscle 2018, 9, 3–19. [Google Scholar] [CrossRef]
- Ko, S.U.; Ling, S.M.; Winters, J.; Ferrucci, L. Age-related mechanical work expenditure during normal walking: The Baltimore longitudinal study of aging. J. Biomech. 2009, 42, 1834–1839. [Google Scholar] [CrossRef] [Green Version]
- Hunter, I.; Lee, K.; Ward, J.; Tracy, J. Self-optimization of stride length among experienced and inexperienced runners. Int. J. Exerc. Sci. 2017, 10, 446–453. [Google Scholar]
- Cai, M.; Yang, E.J. Hochu-Ekki-To improves motor function in an amyotrophic lateral sclerosis animal model. Nutrients 2019, 11, 2644. [Google Scholar] [CrossRef] [Green Version]
- Matsubara, K.; Okuda, M.; Shibata, S.; Miyaki, S.; Ohkubo, T.; Izu, H.; Fujii, T. The delaying effect of alpha-glycerophosphocholine on senescence, transthyretin deposition, and osteoarthritis in senescence-accelerated mouse prone 8 mice. Biosci. Biotechnol. Biochem. 2018, 82, 647–653. [Google Scholar] [CrossRef] [Green Version]
- Costall, B.; Jones, B.J.; Kelly, M.E.; Naylor, R.J.; Tomkins, D.M. Exploration of mice in a black and white test box: Validation as a model of anxiety. Pharmacol. Biochem. Behav. 1989, 32, 777–785. [Google Scholar] [CrossRef]
- Seibenhener, M.L.; Wooten, M.C. Use of the open field maze to measure locomotor and anxiety-like behavior in mice. J. Vis. Exp. 2015, 96, e52434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan-Vazquez, A.; Rye, N.; Ameri, M.; McSparron, B.; Smallwood, G.; Bickerdyke, J.; Rathbone, A.; Dajas-Bailador, F.; Toledo-Rodriguez, M. Impact of voluntary exercise and housing conditions on hippocampal glucocorticoid receptor, miR-124 and anxiety. Mol. Brain. 2015, 8, 40. [Google Scholar] [CrossRef] [Green Version]
- Dong, J.; Zhao, J.; Lin, Y.; Liang, H.; He, X.; Zheng, X.; Sui, M.; Zhuang, Z.; Yan, T. Exercise improves recognition memory and synaptic plasticity in the prefrontal cortex for rats modelling vascular dementia. Neurol. Res. 2018, 40, 68–77. [Google Scholar] [CrossRef] [PubMed]
- Groot, C.; Hooghiemstra, A.M.; Raijmakers, P.G.; van Berckel, B.N.; Scheltens, P.; Scherder, E.J.; van der Flier, W.M.; Ossenkoppele, R. The effect of physical activity on cognitive function in patients with dementia: A meta-analysis of randomized control trials. Ageing Res. Rev. 2016, 25, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Hyodo, K.; Dan, I.; Kyutoku, Y.; Suwabe, K.; Byun, K.; Ochi, G.; Kato, M.; Soya, H. The association between aerobic fitness and cognitive function in older men mediated by frontal lateralization. Neuroimage 2015, 125, 291–300. [Google Scholar] [CrossRef] [PubMed]
- Wakita, Y.; Shimomura, Y.; Kitada, Y.; Yamamoto, H.; Ohashi, Y.; Matsumoto, M. Taxonomic classification for microbiome analysis, which correlates well with the metabolite milieu of the gut. BMC Microbiol. 2018, 18, 188. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.; Zeng, B.; Zeng, L.; Du, X.; Li, B.; Huo, R.; Liu, L.; Wang, H.; Dong, M.; Pan, J.; et al. Gut microbiota regulates mouse behaviors through glucocorticoid receptor pathway genes in the hippocampus. Transl. Psychiatry 2018, 8, 187. [Google Scholar] [CrossRef]
- Felice, V.D.; O’Mahony, S.M. The microbiome and disorders of the central nervous system. Pharmacol. Biochem. Behav. 2017, 160, 1–13. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Repeated Measures Two-Way ANOVA | Two-Way ANOVA | ||
|---|---|---|---|
| Alive Mice at 96 Weeks of Age | All Mice | ||
| Balancing Ability on an Acryl Rod (Test 1) | age | F (11,253) = 20.0, p < 0.001 | F (11,365) = 16.6, p < 0.001 |
| group | F (123) = 1.63, p > 0.05 | F (1365) = 14.8, p < 0.001 | |
| age × group | F (11,253) =1.59, p > 0.05 | F (11,365) = 1.04, p > 0.05 | |
| Horizontal Bar Test (Test 2) | age | F (11,253) = 16.0, p < 0.001 | F (11,365) = 17.1, p < 0.001 |
| group | F (123) = 4.10, p = 0.054 | F (1365) = 30.5, p < 0.001 | |
| age × group | F (11,253) = 1.00, p > 0.05 | F (11,365) = 0.99, p > 0.05 | |
| Wire Hanging Test (Test 3) | age | F (11,253) = 11.0, p < 0.001 | F (11,365) = 10.5, p < 0.001 |
| group | F (123) = 26.7, p < 0.001 | F (1365) = 179.0, p < 0.001 | |
| age × group | F (11,253) = 1.00, p < 0.05 | F (11,365) = 2.16, p < 0.05 |
| Foot Print Test (Test 4) | Repeated Measures Two-Way ANOVA | Two-Way ANOVA | |
|---|---|---|---|
| Alive Mice at 93 Weeks of Age | All Mice | ||
| Stride of forefoot | age | F (5120) = 16.4, p < 0.001 | F (5182) = 18.0, p < 0.001 |
| group | F (124) = 16.4, p < 0.05 | F (1182) = 4.45, p < 0.05 | |
| age × group | F (5120) = 1.73, p > 0.05 | F (5182) = 1.85, p > 0.05 | |
| Sway of forefoot | age | F (5120) = 10.8, p < 0.001 | F (5182) = 12.2, p < 0.001 |
| group | F (124) = 4.15, p = 0.051 | F (1182) = 3.86, p = 0.051 | |
| age × group | F (5120) = 0.70, p > 0.05 | F (5182) = 1.56, p > 0.05 | |
| Stance of forefoot | age | F (5120) = 2.48, p < 0.05 | F (5182) = 3.29, p < 0.01 |
| group | F (124) = 0.245, p > 0.05 | F (1182) = 0.0007, p > 0.05 | |
| age × group | F (5120) = 1.30, p > 0.05 | F (5182) = 1.16, p > 0.05 | |
| Stride of hindfoot | age | F (5120) = 8.51, p < 0.001 | F (5182) = 9.06, p < 0.001 |
| group | F (124) = 14.4, p < 0.001 | F (1182) = 8.26, p < 0.01 | |
| age × group | F (5120) = 1.19, p > 0.05 | F (5182) = 1.08, p > 0.05 | |
| Sway of hindfoot | age | F (5120) = 2.70, p < 0.05 | F (5182) = 4.65, p < 0.001 |
| group | F (124) = 13.4, p < 0.01 | F (1182) = 10.9, p < 0.01 | |
| age × group | F (5120) = 2.01, p > 0.05 | F (5182) = 1.31, p > 0.05 | |
| Stance of hindfoot | age | F (5120) = 4.56, p < 0.01 | F (5182) = 3.39, p < 0.01 |
| group | F (124) = 0.85, p > 0.05 | F (1182) = 1.59, p > 0.05 | |
| age × group | F (5120) = 1.37, p > 0.05 | F (5182) = 1.50, p > 0.05 | |
| Locomotor Activity (Test 5) | Repeated Measures Two-Way ANOVA | Two-Way ANOVA | |
|---|---|---|---|
| Alive Mice at 96 Weeks of Age | All Mice | ||
| Locomotor activity | age | F(8168) = 8.75, p < 0.001 | F (8268) = 6.76, p < 0.001 |
| group | F (121) = 0.032, p > 0.05 | F (1268) = 0.083, p > 0.05 | |
| age × group | F (8168) =0.95, p > 0.05 | F (8268) = 0.554, p > 0.05 | |
| Center time | age | F (8168) = 29.0, p < 0.001 | F (8268) = 26.0, p < 0.001 |
| group | F (F(121) = 1.20, p > 0.05 | F (1268) = 0.544, p > 0.05 | |
| age × group | F (8168) =4.20, p < 0.01 | F (8268) = 2.83, p < 0.001 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shimizu, C.; Wakita, Y.; Tsuchiya, Y.; Nabeshima, T. Influence of Housing Systems on Physical, Emotional, and Cognitive Functions with Aging in DBA/2CrSlc Mice. Animals 2020, 10, 746. https://doi.org/10.3390/ani10040746
Shimizu C, Wakita Y, Tsuchiya Y, Nabeshima T. Influence of Housing Systems on Physical, Emotional, and Cognitive Functions with Aging in DBA/2CrSlc Mice. Animals. 2020; 10(4):746. https://doi.org/10.3390/ani10040746
Chicago/Turabian StyleShimizu, Chikako, Yoshihisa Wakita, Youichi Tsuchiya, and Toshitaka Nabeshima. 2020. "Influence of Housing Systems on Physical, Emotional, and Cognitive Functions with Aging in DBA/2CrSlc Mice" Animals 10, no. 4: 746. https://doi.org/10.3390/ani10040746