Dietary Supplemented Curcumin Improves Meat Quality and Antioxidant Status of Intrauterine Growth Retardation Growing Pigs via Nrf2 Signal Pathway

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Care
2.2. Animals and Study Design
2.3. Growth Performance Analysis
2.4. Meat Quality Analysis
2.5. Assay of Antioxidant Capacity in Leg Muscle
2.6. Mineral Concentration in Leg Muscle
2.7. Determination of mRNA Expression
2.8. Western Blotting Anlysis
2.9. Statistical Analysis
3. Results
3.1. Growth Performance
3.2. Meat Quality
3.3. Antioxidant Enzyme Activity, MDA and Protein Carbonyls (PC)
3.4. Mineral Concentration in Leg Muscles
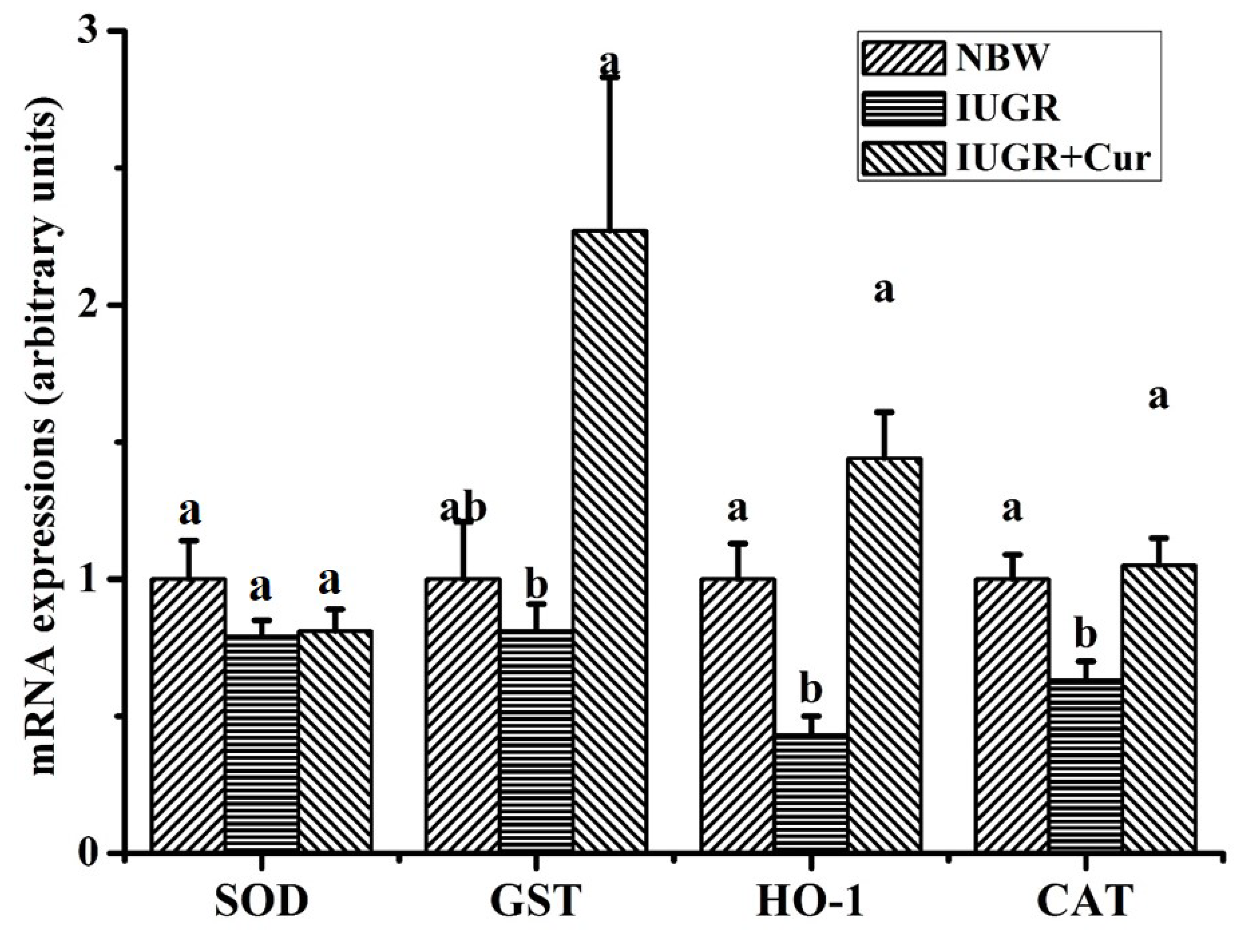
3.5. Gene Expression
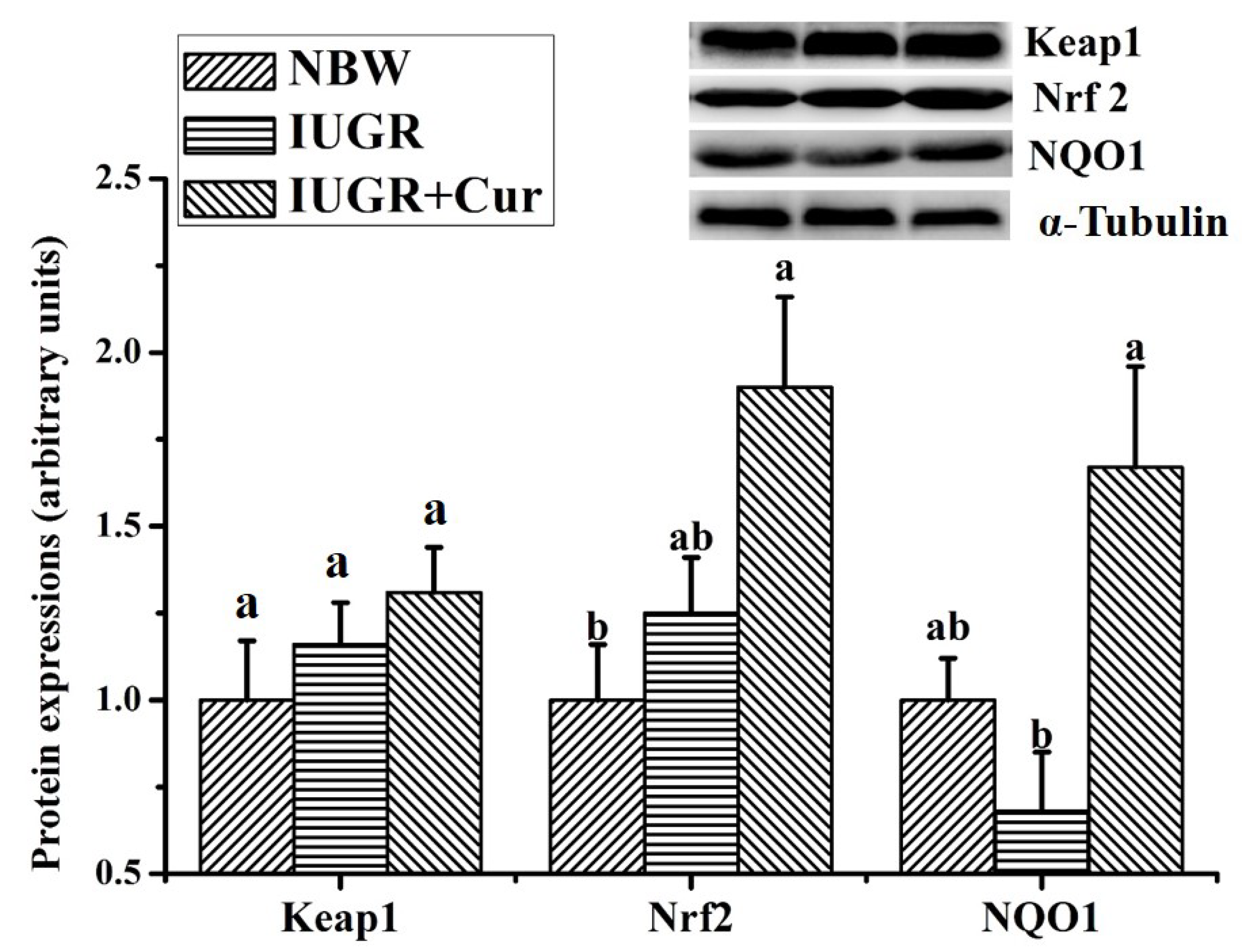
3.6. Protein Expressions of Keap1/Nrf2 Signal Pathway in Leg Muscle
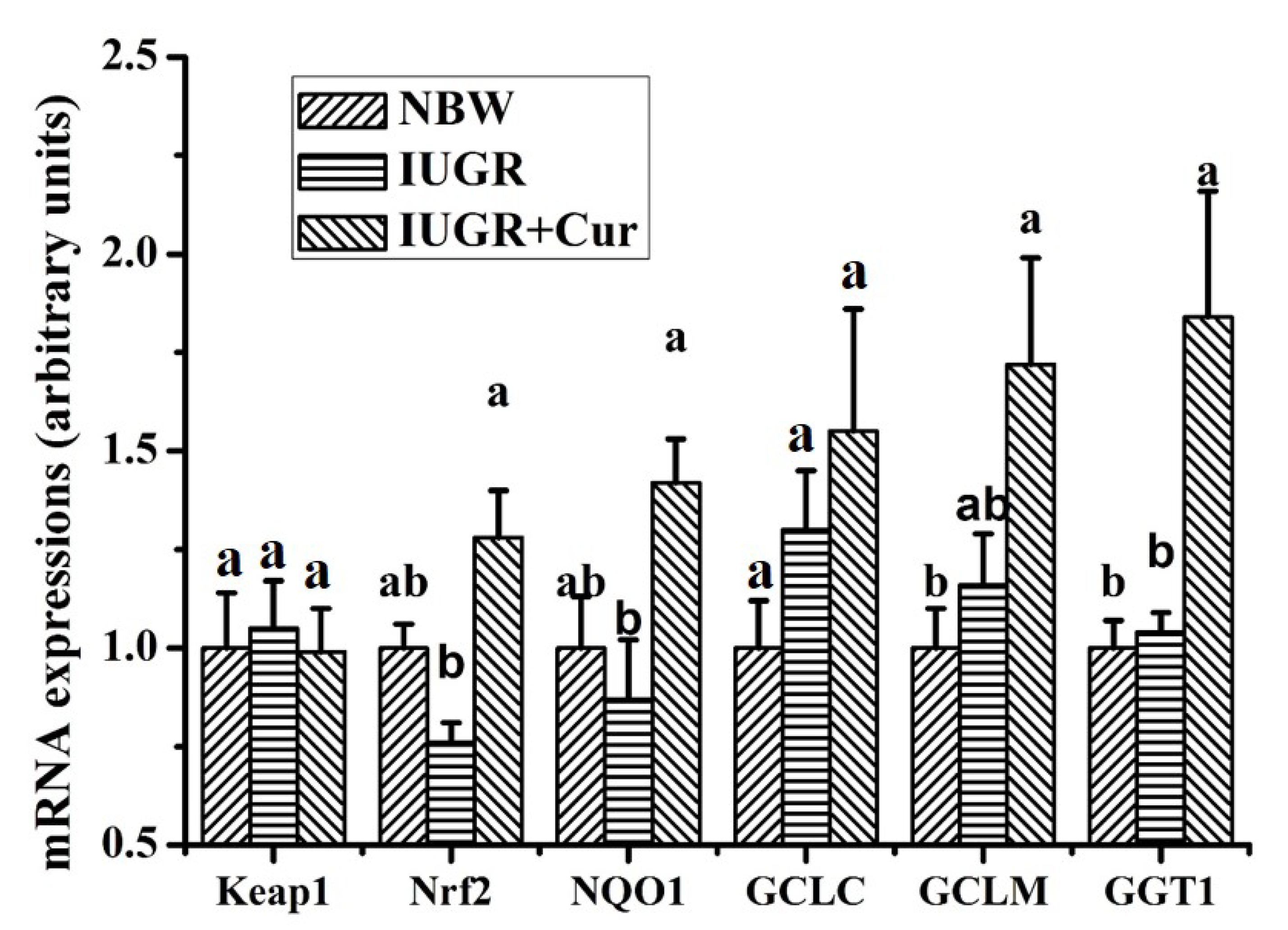
3.7. Gene Expressions of the Keap1/Nrf2 Signal Pathway in Leg Muscles
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wu, G.; Bazer, F.W.; Wallace, J.M.; Spencer, T.E. Board-invited review: Intrauterine growth retardation: Implications for the animal sciences. J. Anim. Sci. 2006, 84, 2316–2337. [Google Scholar] [CrossRef] [PubMed]
- Srividhya, S.; Kyle, P.M. Aetiology and pathogenesis of IUGR. Best Pract. Res. Clin. Obstet. Gynaecol. 2009, 23, 765–777. [Google Scholar]
- Barker, D.J.P. A new model for the origins of chronic disease. Med. Health Care Philos. 2001, 4, 31–35. [Google Scholar] [CrossRef]
- Langley-Evans, S. Nutrition in early life and the programming of adult disease: A review. J. Hum. Nutr. Diet. 2015, 28, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Li, Y.; Wang, T. Antioxidant capacity and concentration of redox-active trace mineral in fully weaned intra-uterine growth retardation piglets. J. Anim. Sci. Biotechnol. 2015, 6, 48. [Google Scholar] [CrossRef] [Green Version]
- Falowo, A.B.; Fayemi, P.O.; Muchenje, V. Natural antioxidants against lipid–protein oxidative deterioration in meat and meat products: A review. Food Res. Int. 2014, 64, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Luo, J.Q.; Yu, B.; Zheng, P.; Huang, Z.Q.; Mao, X.B.; He, J.; Yu, J.; Chen, J.; Chen, D. Dietary resveratrol supplementation improves meat quality of finishing pigs through changing muscle fiber characteristics and antioxidative status. Meat Sci. 2015, 102, 15–21. [Google Scholar] [CrossRef]
- Fasseas, M.K.; Mountzouris, K.C.; Tarantilis, P.A.; Polissiou, M.; Zervas, G. Antioxidant activity in meat treated with oregano and sage essential oils. Food Chem. 2008, 106, 1188–1194. [Google Scholar] [CrossRef]
- Xu, D.P.; Li, Y.; Meng, X.; Zhou, T.; Zhou, Y.; Zheng, J.; Zhang, J.J.; Li, H.B. Natural antioxidants in foods and medicinal plants: Extraction, assessment and resources. Int. J. Mol. Sci. 2017, 18, 96. [Google Scholar] [CrossRef]
- Hewlings, S.; Kalman, D. Curcumin: A review of its’ effects on human health. Foods 2017, 6, 92. [Google Scholar] [CrossRef]
- Aggarwal, B.B.; Harikumar, K.B. Potential therapeutic effects of curcumin, the anti-inflammatory agent, against neurodegenerative, cardiovascular, pulmonary, metabolic, autoimmune and neoplastic diseases. Int. J. Biochem. Cell Biol. 2009, 41, 40–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panahi, Y.; Hosseini, M.S.; Khalili, N.; Naimi, E.; Simental-Mendía, L.E.; Majeed, M.; Sahebkar, A. Effects of curcumin on serum cytokine concentrations in subjects with metabolic syndrome: A post-hoc analysis of a randomized controlled trial. Biomed. Pharmacother. 2016, 82, 578–582. [Google Scholar] [CrossRef] [PubMed]
- Sahin, K.; Orhan, C.; Tuzcu, Z.; Tuzcu, M.; Sahin, N. Curcumin ameloriates heat stress via inhibition of oxidative stress and modulation of Nrf2/HO-1 pathway in quail. Food Chem. Toxicol. 2012, 50, 4035–4041. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Hu, Z.; Lu, C.; Bai, K.; Zhang, L.; Wang, T. Effect of various levels of dietary curcumin on meat quality and antioxidant profile of breast muscle in broilers. J. Agric. Food Chem. 2015, 63, 3880–3886. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Niu, Y.; Wang, F.; Wang, C.; Cui, T.; Bai, K.; Zhang, J.; Zhong, X.; Zhang, L.; Wang, T. Dietary curcumin supplementation attenuates inflammation, hepatic injury and oxidative damage in a rat model of intra-uterine growth retardation. British J. Nutr. 2018, 120, 537–548. [Google Scholar] [CrossRef] [Green Version]
- Farombi, E.O.; Shrotriya, S.; Na, H.K.; Kim, S.H.; Surh, Y.J. Curcumin attenuates dimethylnitrosamine-induced liver injury in rats through Nrf2-mediated induction of heme oxygenase-1. Food Chem. Toxicol. 2008, 46, 1279–1287. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zheng, X.C.; Siyal, F.A.; Babazadeh, D.; Ayasan, T.; Cheng, K.; He, J.H.; Xu, W.; Zhang, L.L.; Zhong, X.; et al. Altered tissue mineralization, increased hepatic lipid and inhibited autophagy in intrauterine growth retardation piglets. S. Afr. J. Anim. Sci. 2018, 48, 56–66. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Su, W.P.; Ying, Z.X.; Chen, Y.P.; Zhou, L.; Li, Y.; Zhang, J.; Zhang, L.; Wang, T. N-acetylcysteine attenuates intrauterine growth retardation-induced hepatic damage in suckling piglets by improving glutathione synthesis and cellular homeostasis. Eur. J. Nutr. 2018, 57, 327–338. [Google Scholar] [CrossRef] [PubMed]
- Ying, Z.X.; Ge, X.K.; Zhang, H.; Su, W.P.; Li, Y.; Zhou, L.; Zhang, L.; Wang, T. Effects of dietary methionine restriction on postnatal growth, insulin sensitivity, and glucose metabolism in intrauterine growth retardation pigs at 49 and 105 d of age. J. Anim. Sci. 2019, 97, 610–619. [Google Scholar] [CrossRef] [PubMed]
- Lu, N.; Qiu, J.Y.; Ying, Z.X.; Zheng, Y.M.; Wang, T.; Zhong, X. Effects of varying levels of dietary curcumin on growth performance, digestibility and serum parameters of weaned piglets. J. Domestic. Anim. Ecol. 2017, 38, 30–35. [Google Scholar]
- Demirbaş, A. Proximate and heavy metal composition in chicken meat and tissues. Food Chem. 1999, 67, 27–31. [Google Scholar] [CrossRef]
- Wang, C.; Lu, J.; Le Zhou, J.L.; Xu, J.; Li, W.; Zhang, L.; Zhong, X.; Wang, T. Effects of long-term exposure to zinc oxide nanoparticles on development, zinc metabolism and biodistribution of minerals (Zn, Fe, Cu, Mn) in mice. PLoS ONE 2016, 11, e0164434. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(T)(-Delta Delta C) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Bai, K.; He, J.; Su, W.; Dong, L.; Zhang, L.; Wang, T. Leucine improves growth performance of intrauterine growth retardation piglets by modifying gene and protein expression related to protein synthesis. Nutrition 2016, 32, 114–121. [Google Scholar] [CrossRef] [PubMed]
- Shahkhalili, Y.; Moulin, J.; Zbinden, I.; Aprikian, O.; Macé, K. Comparison of two models of intrauterine growth restriction for early catch-up growth and later development of glucose intolerance and obesity in rats. Am. J. Physiol. 2010, 298, 141–146. [Google Scholar] [CrossRef]
- Beltrand, J.; Nicolescu, R.; Kaguelidou, F.; Verkauskiene, R.; Sibony, O.; Chevenne, D.; Claris, O.; Levy-Marchal, C. Catch-up growth following fetal growth restriction promotes rapid restoration of fat mass but without metabolic consequences at one year of age. PLoS ONE 2009, 4, e5343. [Google Scholar] [CrossRef]
- Attig, L.; Djiane, J.; Gertler, A.; Rampin, O.; Larcher, T.; Boukthir, S.; Anton, P.M.; Madec, J.Y.; Gourdou, I.; Abdennebi-Najar, L. Study of hypothalamic leptin receptor expression in low-birth-weight piglets and effects of leptin supplementation on neonatal growth and development. Am. J. Physiol. Endocrinol. Metab. 2008, 295, E1117. [Google Scholar] [CrossRef]
- Poore, K.R.; Fowden, A.L. The effects of birth weight and postnatal growth patterns on fat depth and plasma leptin concentrations in juvenile and adult pigs. J. Physiol. 2004, 558, 295–304. [Google Scholar] [CrossRef]
- Desai, M.; Gayle, D.; Babu, J.; Ross, M.G. Programmed obesity in intrauterine growth-restricted newborns: Modulation by newborn nutrition. Am. J. Physiol. Reg. Integr. Comp. Physiol. 2005, 288, R91–R96. [Google Scholar] [CrossRef]
- Fernandez-Feijoo, C.D.; Carrasco, C.C.; Francisco, N.V.; Romero, J.C.; Lorenzo, J.R.F.; Jiménez-Chillaron, J.C.; Camprubi, M.C. Influence of catch up growth on spatial learning and memory in a mouse model of intrauterine growth restriction. PLoS ONE 2017, 12, 15. [Google Scholar] [CrossRef] [Green Version]
- De Blasio, M.J.; Gatford, K.L.; Robinson, J.S.; Owens, J.A. Placental restriction of fetal growth reduces size at birth and alters postnatal growth, feeding activity, and adiposity in the young lamb. Am. J. Physiol. Reg. Integr. Comp. Physiol. 2007, 292, R875–R886. [Google Scholar] [CrossRef]
- Xun, W.J.; Shi, L.G.; Zhou, H.L.; Hou, G.Y.; Cao, T.; Zhao, C.P. Effects of curcumin on growth performance, jejunal mucosal membrane integrity, morphology and immune status in weaned piglets challenged with enterotoxigenic Escherichia coli. Int. Immunopharmacol. 2015, 27, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Wu, X.Y.; Zhou, X.Q.; Feng, L.; Liu, Y.; Jiang, W.D.; Wu, P.; Zhao, Y. Effects of dietary curcumin supplementation on growth performance, intestinal digestive enzyme activities and antioxidant capacity of crucian carp Carassius auratus. Aquaculture 2016, 463, 174–180. [Google Scholar] [CrossRef]
- Nissen, P.M.; Oksbjerg, N. Birth weight and postnatal dietary protein level affect performance, muscle metabolism and meat quality in pigs. Animal 2011, 5, 1382–1389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alvarenga, A.L.N.; Chiarini-Garcia, H.; Cardeal, P.C.; Moreira, L.P.; Foxcroft, G.R.; Fontes, D.O.; Almeida, F.R.C.L. Intra-uterine growth retardation affects birthweight and postnatal development in pigs, impairing muscle accretion, duodenal mucosa morphology and carcass traits. Reprod. Fertil. Develop. 2013, 25, 387–395. [Google Scholar] [CrossRef] [PubMed]
- Mancini, S.; Paci, G.; Pisseri, F.; Preziuso, G. Effect of turmeric (Curcuma longa L.) powder as dietary antioxidant supplementation on pig meat quality. J. Food Process. Preserv. 2017, 41, e12878. [Google Scholar] [CrossRef]
- Kohen, R.; Nyska, A. Invited review: Oxidation of biological systems: Oxidative stress phenomena, antioxidants, redox reactions, and methods for their quantification. Toxicol. Pathol. 2002, 30, 620–650. [Google Scholar] [CrossRef] [Green Version]
- Emerit, J.; Edeas, M.; Bricaire, F. Neurodegenerative diseases and oxidative stress. Biomed. Pharmacother. 2004, 58, 39–46. [Google Scholar] [CrossRef]
- Caperna, T.J.; Shannon, A.E.; Blomberg, L.A.; Garrett, W.M.; Ramsay, T.G. Identification of protein carbonyls in serum of the fetal and neonatal pig. Comp. Biochem. Physiol. Part B 2010, 156, 189–196. [Google Scholar] [CrossRef] [Green Version]
- Del Rio, D.; Stewart, A.J.; Pellegrini, N. A review of recent studies on malondialdehyde as toxic molecule and biological marker of oxidative stress. Nutr. Metab. Cardiovasc. Dis. 2005, 15, 316–328. [Google Scholar] [CrossRef]
- Scandalios, J. Oxidative stress: Molecular perception and transduction of signals triggering antioxidant gene defenses. Braz. J. Med. Biol. Res. 2005, 38, 995–1014. [Google Scholar] [CrossRef] [PubMed]
- Zadrożna, M.; Gawlik, M.; Nowak, B.; Marcinek, A.; Mrowiec, H.; Walas, S.; Wietecha-Posłuszny, R.; Zagrodzki, P. Antioxidants activities and concentration of selenium, zinc and copper in preterm and IUGR human placentas. J. Trace Elem. Med. Biol. 2009, 23, 144–148. [Google Scholar] [CrossRef] [PubMed]
- Ferencz, A.; Orvos, H.; Hermesz, E. Major differences in the levels of redox status and antioxidant defence markers in the erythrocytes of pre- and full-term neonates with intrauterine growth restriction. Reprod. Toxicol. 2015, 53, 10–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Xu, L.; Zhang, L.; Ying, Z.; Su, W.; Wang, T. Curcumin attenuates d-galactosamine/lipopolysaccharide–induced liver injury and mitochondrial dysfunction in mice. J. Nutr. 2014, 144, 1211–1218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sykiotis, G.P.; Bohmann, D. Keap1/Nrf2 Signaling Regulates Oxidative Stress Tolerance and Lifespan in Drosophila. Dev. Cell 2008, 14, 76–85. [Google Scholar] [CrossRef] [Green Version]
- Mi, S.S.; Hye, Y.J.; Hwan, K.S. Role of the Nrf2-ARE pathway in liver diseases. Oxidative Med. Cell. Longev. 2013, 2013, 763257. [Google Scholar]
- Michael, M.; Ken, I.; Masayuki, Y.; Hayes, D.J. Keap1-dependent proteasomal degradation of transcription factor Nrf2 contributes to the negative regulation of antioxidant response element-driven gene expression. J. Biol. Chem. 2003, 278, 21592–21600. [Google Scholar]
- Sykiotis, G.P.; Habeos, I.G.; Samuelson, A.V.; Dirk, B. The role of the antioxidant and longevity-promoting Nrf2 pathway in metabolic regulation. Curr. Opin. Clin. Nutr. 2011, 14, 41–48. [Google Scholar] [CrossRef] [Green Version]
- Hansen, M.; Porosk, R.; Mahlapuu, R.; Kairane, C.; Kilk, K.; Soomets, U. GSH synthetic analogue O-methyl-L-tyrosinylglutathione regulates Nrf2-mediated expression of GCLc and GCLm. J. Chem. 2019. [Google Scholar] [CrossRef]
- Receno, C.N.; Liang, C.; Korol, D.L.; DeRuisseau, K.C. Prolonged dietary curcumin supplementation augments Nrf2 nuclear translocation and attenuates oxidative stress in skeletal muscle of aging rats. Faseb. J. 2018, 32, 856. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Gene | Accession No. | Sequence (5′-3′) | Size (bp) |
|---|---|---|---|
| GAPDH | NM_001206359.1 | F-CGTCCCTGAGACACGATGGT | 194 |
| R-GCCTTGACTGTGCCGTGGAAT | |||
| Keap1 | XM_021076667.1 | F: CGTGGAGACAGAAACGTGGA | 239 |
| R: CAATCTGCTTCCGACAGGGT | |||
| Nrf2 | NM_001114671.1 | F: GACAAACCGCCTCAACTCAG | 183 |
| R: GTCTCCACGTCGTAGCGTTC | |||
| CAT | XM_021081498.1 | F:AGCTTTGCCCTTGCACAAAC | 119 |
| R:ACATCCTGAACAAGAAGGGGC | |||
| GST | NM_214389.2 | F:TTCAATGGCCGAGGCAGAAT | 216 |
| R: GAGGTTGTACTTGGTGGCGA | |||
| SOD | NM_001190422.1 | F: CATTCCATCATTGGCCGCAC | 118 |
| R: TTACACCACAGGCCAAACGA | |||
| HO-1 | NM_001004027.1 | F: CAAGCAGAAAATCCTCGAAG | 241 |
| R: GCTGAGTGTCAGGACCCATC | |||
| NQO1 | NM_001159613.1 | F: GATCATACTGGCCCACTCCG | 200 |
| R: GAGCAGTCTCGGCAGGATAC | |||
| GCLC | XM_003482164.4 | F:GGCGACGAGGTGGAATACAT | 123 |
| R: GTTTGGGTTTGTCCTTTCCCC | |||
| GCLM | XM_001926378.4 | F: GCATCTACAGCCTTACTGGGA | 180 |
| R: GTTAAATCGGGCGGCATCAC | |||
| GGT1 | NM_214030.1 | F:ATCACACCAGGAAAACAGCCA | 118 |
| R: CGGTAGATGTGGTGATCTGTGT |
| Item 1 | NBW | IUGR | IUGR + Cur | p |
|---|---|---|---|---|
| Body weight(Kg) | ||||
| 0 d | 1.49 ± 0.02 a | 0.75 ± 0.01 b | 0.76 ± 0.01 b | <0.01 |
| 26 d | 7.52 ± 0.35 a | 5.76 ± 0.20 b | 6.05 ± 0.10 b | <0.01 |
| 56 d | 17.03 ± 0.42 a | 13.39 ± 0.46 b | 13.96 ± 0.29 b | <0.01 |
| 115 d | 57.53 ± 1.31 | 53.46 ± 1.80 | 53.52 ± 1.53 | 0.12 |
| ADG(Kg) | ||||
| 0-26 d | 0.25 ± 0.01 a | 0.20 ± 0.01 b | 0.20 ± 0.01 b | <0.01 |
| 26-56 d | 0.30 ± 0.01 a | 0.25 ± 0.02 b | 0.26 ± 0.01 b | 0.02 |
| 56-115 d | 0.69 ± 0.02 | 0.68 ± 0.03 | 0.67 ± 0.03 | 0.91 |
| ADFI(Kg/d) | ||||
| 56-115 d | 2.35 ± 0.02 a | 2.31 ± 0.02 a | 2.17 ± 0.05 b | <0.01 |
| F:G | ||||
| 56-115 d | 3.43 ± 0.08 | 3.41 ± 0.08 | 3.25 ± 0.12 | 0.40 |
| Item | NBW | IUGR | IUGR+Cur | p |
|---|---|---|---|---|
| pH | ||||
| pH45min | 6.28 ± 0.07 | 6.39 ± 0.08 | 6.33 ± 0.11 | 0.70 |
| pH24h | 6.26 ± 0.01 | 6.21 ± 0.04 | 6.25 ± 0.03 | 0.42 |
| Drip loss | ||||
| 24 h | 0.18 ± 0.01a | 0.20 ± 0.01a | 0.14 ± 0.01b | <0.01 |
| 48 h | 0.22 ±0.01a | 0.25 ± 0.01a | 0.17 ± 0.01b | <0.01 |
| Meat color | ||||
| L* | 45.10 ± 1.58 | 44.90 ± 0.78 | 41.66 ± 0.79 | 0.08 |
| a* | 9.75 ± 0.30b | 9.90 ± 0.28ab | 11.25 ± 0.49a | 0.02 |
| b* | 17.50 ± 0.62 | 17.70 ± 0.67 | 17.35 ± 0.77 | 0.94 |
| Item 1 | NBW | IUGR | IUGR + Cur | p |
|---|---|---|---|---|
| MDA(nmol/mgprot) | 3.08 ± 0.26 b | 4.65 ± 0.32 a | 3.42 ± 0.19 b | <0.01 |
| PC (nmol/mgprot) | 3.19 ± 0.16 b | 3.85 ± 0.17 a | 3.43 ± 0.15 ab | 0.03 |
| GSH(mg/gprot) | 0.87 ± 0.07 | 0.94 ± 0.04 | 1.03 ± 0.11 | 0.41 |
| GPX (U/mgprot) | 16.65 ± 1.67 a | 13.65 ± 0.53 b | 15.50 ± 0.82 ab | 0.02 |
| CAT(U/mgprot) | 18.12 ± 1.06 a | 14.18 ± 0.32 b | 18.21 ± 0.97 a | <0.01 |
| T-AOC(U/mgprot) | 0.28 ± 0.01 | 0.28 ± 0.01 | 0.29 ± 0.01 | 0.94 |
| SOD(U/mgprot) | 17.38 ± 0.34 ab | 16.69 ± 0.23 b | 17.83 ± 0.28 a | 0.04 |
| TSH(mmol/gprot) | 0.13 ± 0.01 | 0.12 ± 0.01 | 0.12 ± 0.01 | 0.41 |
| POD(U/mgprot) | 0.27 ± 0.02 a | 0.19 ± 0.02 b | 0.29 ± 0.03 a | 0.02 |
| Item | NBW | IUGR | IUGR + Cur | p |
|---|---|---|---|---|
| Zn (mg/kg) | 15.71 ± 1.84 | 11.50 ± 0.91 | 13.42 ± 1.85 | 0.21 |
| Fe (mg/kg) | 100.47 ± 10.11 | 92.50 ± 9.65 | 89.70 ± 9.26 | 0.72 |
| Cu (mg/kg) | 1.04 ± 0.09 | 1.10 ± 0.09 | 1.13 ± 0.05 | 0.72 |
| Mn (mg/kg) | 0.70 ± 0.06 | 0.73 ± 0.02 | 0.75 ± 0.06 | 0.80 |
| Se (mg/kg) | 0.57 ± 0.04 | 0.59 ± 0.04 | 0.58 ± 0.02 | 0.87 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, L.; Zhang, J.; Yan, E.; He, J.; Zhong, X.; Zhang, L.; Wang, C.; Wang, T. Dietary Supplemented Curcumin Improves Meat Quality and Antioxidant Status of Intrauterine Growth Retardation Growing Pigs via Nrf2 Signal Pathway. Animals 2020, 10, 539. https://doi.org/10.3390/ani10030539
Zhang L, Zhang J, Yan E, He J, Zhong X, Zhang L, Wang C, Wang T. Dietary Supplemented Curcumin Improves Meat Quality and Antioxidant Status of Intrauterine Growth Retardation Growing Pigs via Nrf2 Signal Pathway. Animals. 2020; 10(3):539. https://doi.org/10.3390/ani10030539
Chicago/Turabian StyleZhang, Ligen, Jiaqi Zhang, Enfa Yan, Jintian He, Xiang Zhong, Lili Zhang, Chao Wang, and Tian Wang. 2020. "Dietary Supplemented Curcumin Improves Meat Quality and Antioxidant Status of Intrauterine Growth Retardation Growing Pigs via Nrf2 Signal Pathway" Animals 10, no. 3: 539. https://doi.org/10.3390/ani10030539