Hepatic Inflammatory Response to Exogenous LPS Challenge is Exacerbated in Broilers with Fatty Liver Disease

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Treatment
2.2. Histopathology
2.3. Plasma Biochemical Indicators Measurement and Triglycerides Content in the Liver
2.4. Reverse Transcription and Quantitative Real-Time PCR
2.5. Statistical Analysis
3. Results
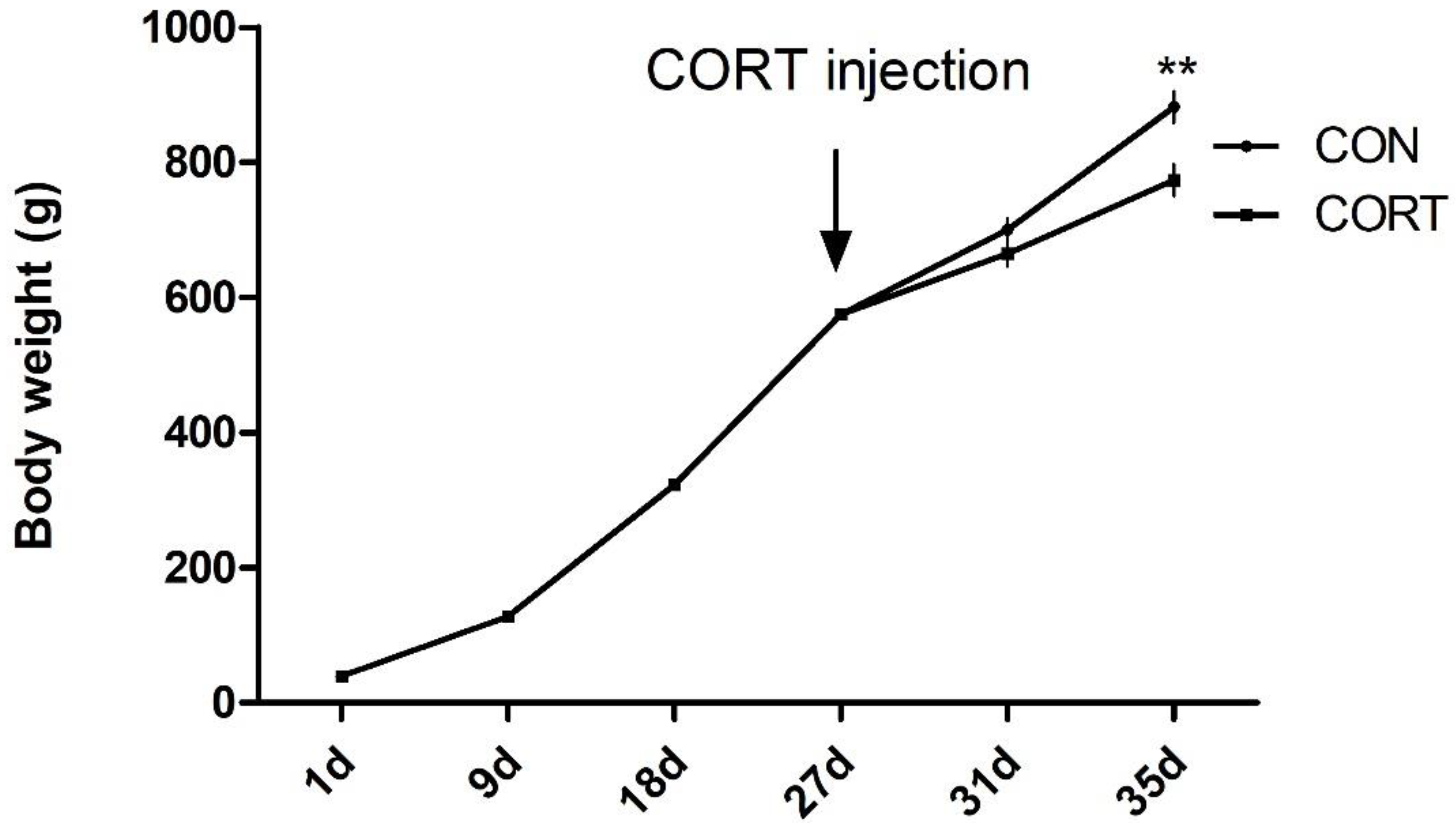
3.1. Growth Performance
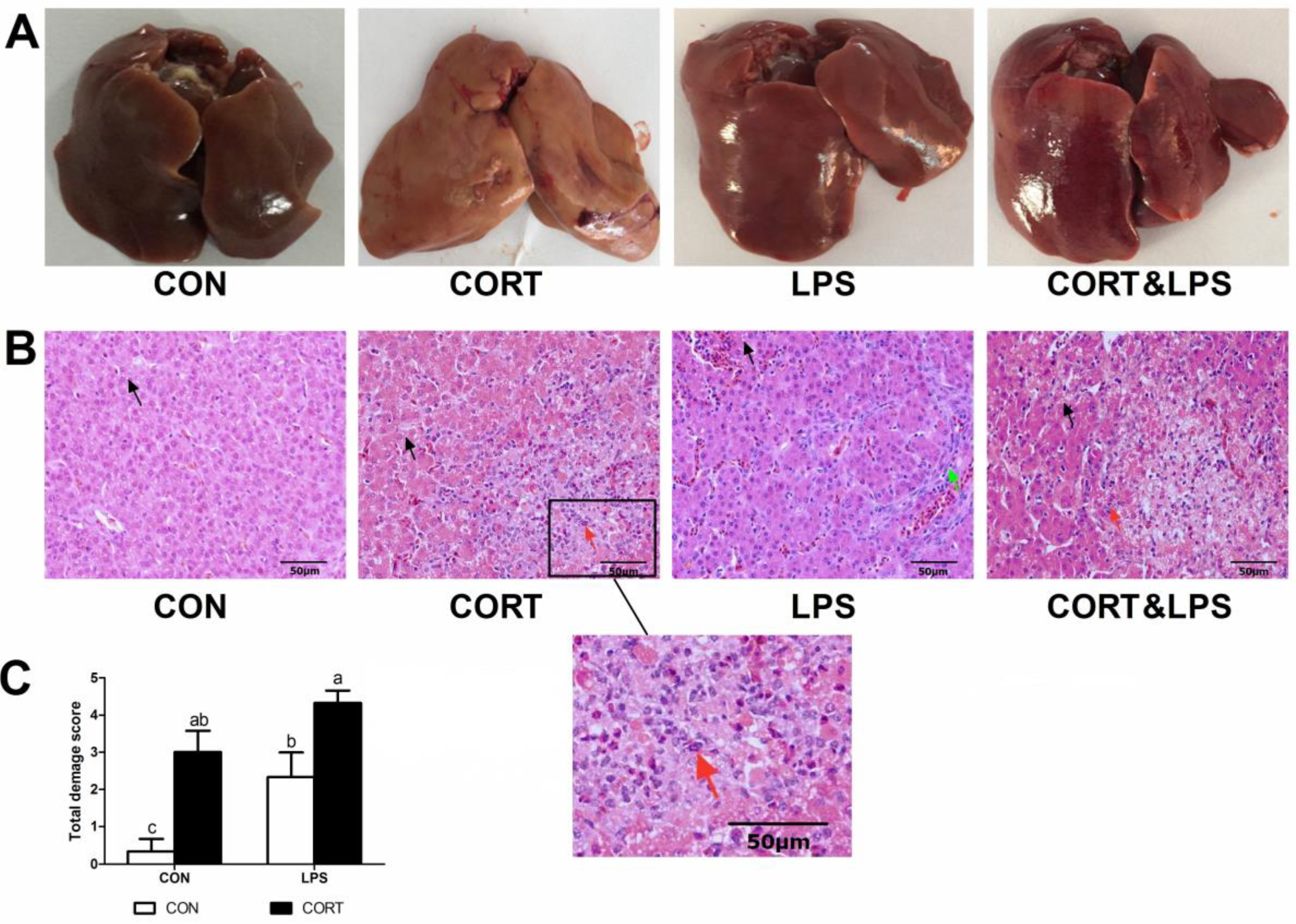
3.2. Hepatic Histological Analysis and Liver Damage Evaluation
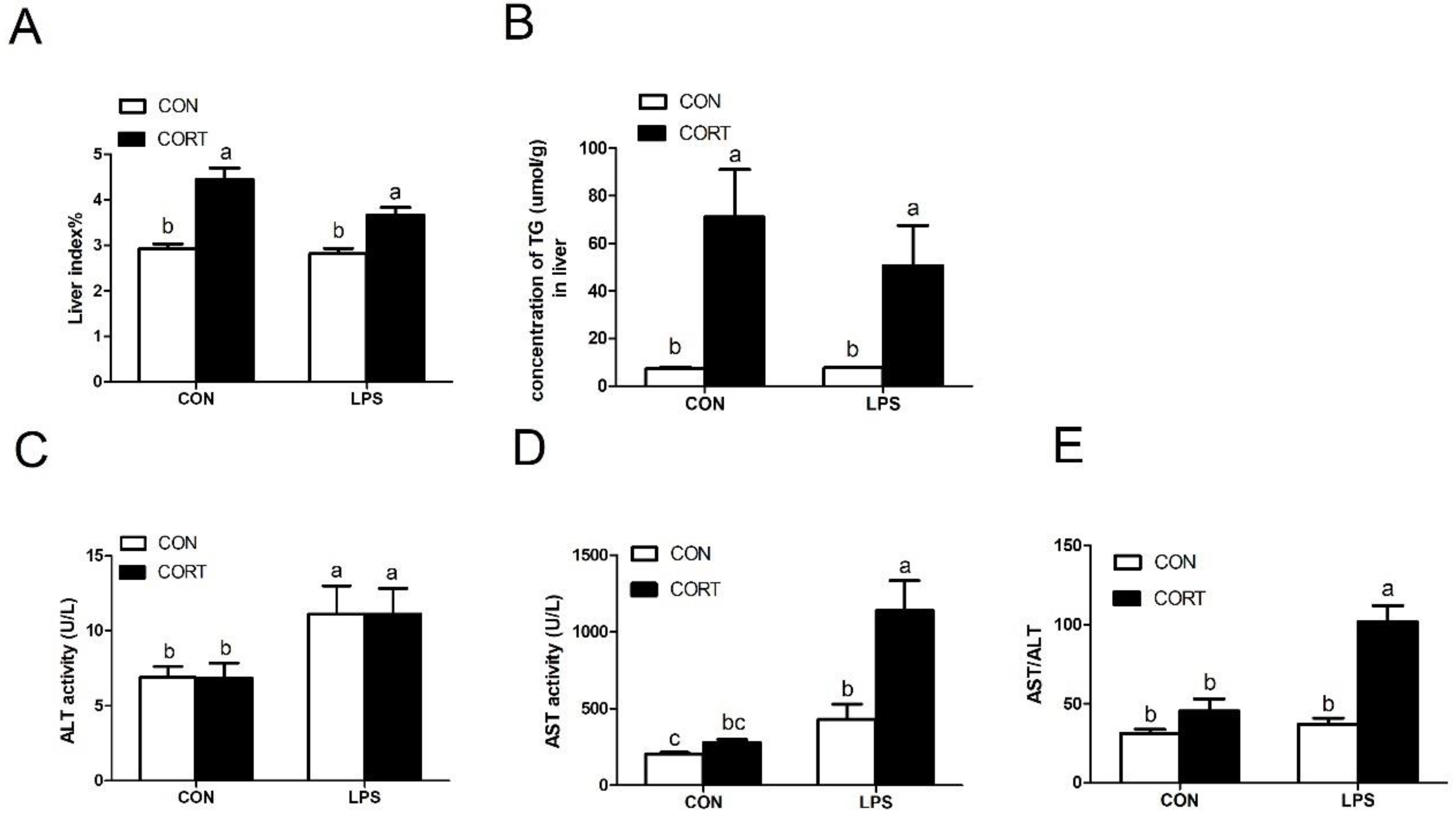
3.3. Plasma Biochemical Indicators
3.4. Liver Damage Index
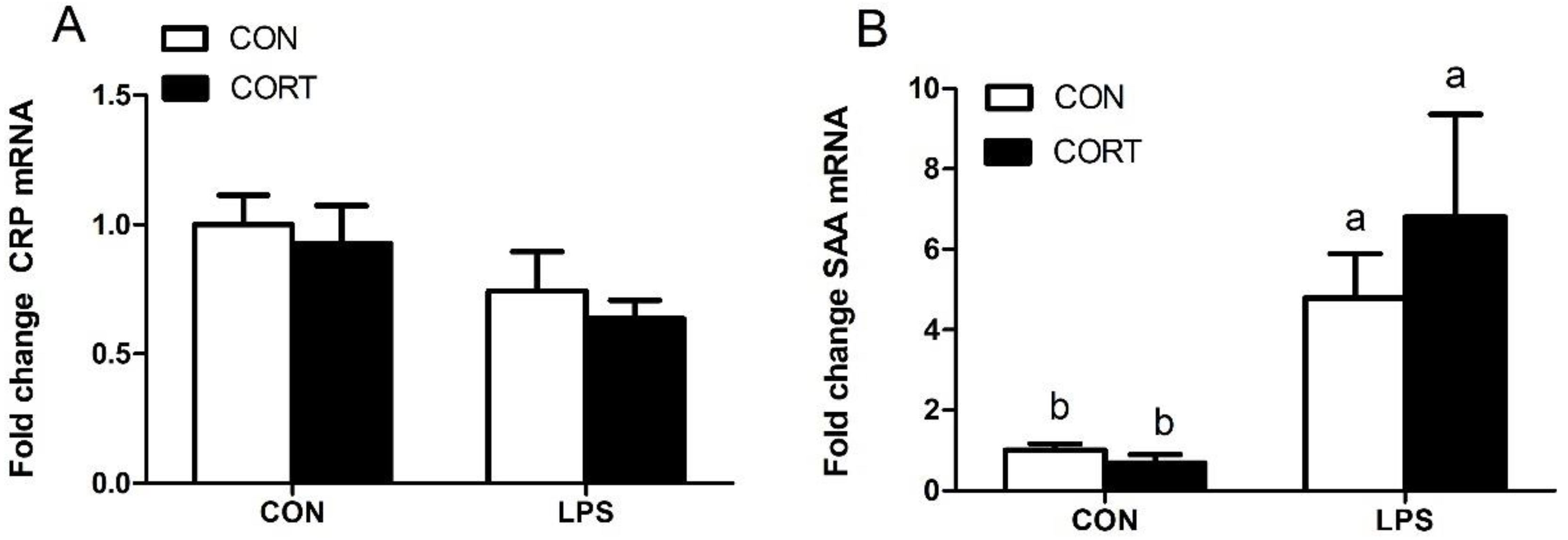
3.5. Hepatic Expression of Genes Encoding Acute-Phase Proteins
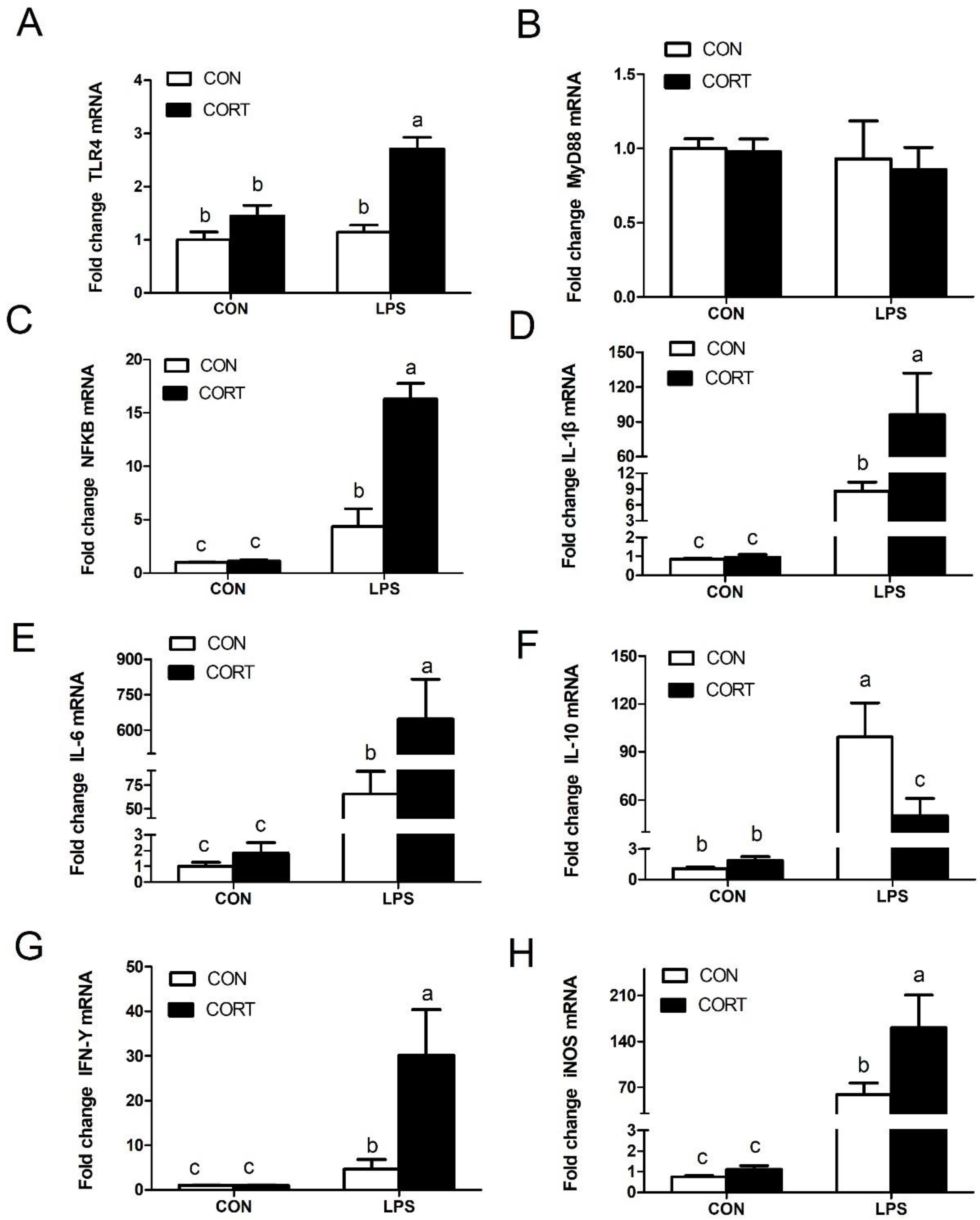
3.6. Liver Expression of Genes Related to Inflammation
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cohen, J.C.; Horton, J.D.; Hobbs, H.H. Human fatty liver disease: Old questions and new insights. Science 2011, 332, 1519–1523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boursier, J.; Mueller, O.; Barret, M.; Machado, M.; Fizanne, L.; Araujo-Perez, F.; Guy, C.D.; Seed, P.C.; Rawls, J.F.; David, L.A. The severity of nonalcoholic fatty liver disease is associated with gut dysbiosis and shift in the metabolic function of the gut microbiota. Hepatology 2016, 63, 764–775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Ding, Y.-L.; Zhang, J.-L.; Zhang, P.; Wang, J.-Q.; Li, Z.-H. Alpinetin improved high fat diet-induced non-alcoholic fatty liver disease (NAFLD) through improving oxidative stress, inflammatory response and lipid metabolism. Biomed. Pharmacother. 2018, 97, 1397–1408. [Google Scholar] [CrossRef] [PubMed]
- Cherian, G.; Holsonbake, T.B.; Goeger, M.P.; Bildfell, R. Dietary CLA alters yolk and tissue FA composition and hepatic histopathology of laying hens. Lipids 2002, 37, 751–757. [Google Scholar] [CrossRef]
- Paglialunga, S.; Dehn, C.A. Clinical assessment of hepatic de novo lipogenesis in non-alcoholic fatty liver disease. Lipids Health Dis. 2016, 15, 159. [Google Scholar] [CrossRef] [Green Version]
- Davis, J.; Cain, J.; Small, C.; Hales, D. Therapeutic effect of flax-based diets on fatty liver in aged laying hens. Poult. Sci. 2016, 95, 2624–2632. [Google Scholar] [CrossRef] [PubMed]
- Woods, C.P.; Hazlehurst, J.M.; Tomlinson, J.W. Glucocorticoids and non-alcoholic fatty liver disease. J. Steroid Biochem. Mol. Biol. 2015, 154, 94–103. [Google Scholar] [CrossRef]
- Hu, Y.; Sun, Q.; Hu, Y.; Hou, Z.; Zong, Y.; Omer, N.A.; Abobaker, H.; Zhao, R. Corticosterone-induced Lipogenesis activation and Lipophagy inhibition in chicken liver are alleviated by maternal Betaine supplementation. J. Nutr. 2018, 148, 316–325. [Google Scholar] [CrossRef] [Green Version]
- Jiang, K.; Jiao, H.; Song, Z.; Yuan, L.; Zhao, J.; Lin, H. Corticosterone administration and dietary glucose supplementation enhance fat accumulation in broiler chickens. Br. Poult. Sci. 2008, 49, 625–631. [Google Scholar] [CrossRef]
- Hu, Y.; Sun, Q.; Liu, J.; Jia, Y.; Cai, D.; Idriss, A.A.; Omer, N.A.; Zhao, R. In ovo injection of betaine alleviates corticosterone-induced fatty liver in chickens through epigenetic modifications. Sci. Rep. 2017, 7, 40251. [Google Scholar] [CrossRef]
- Chanie, M.; Negash, T.; Tilahun, S.B. Occurrence of concurrent infectious diseases in broiler chickens is a threat to commercial poultry farms in Central Ethiopia. Trop. Anim. Health Prod. 2009, 41, 1309. [Google Scholar] [CrossRef] [PubMed]
- Compare, D.; Coccoli, P.; Rocco, A.; Nardone, O.; De Maria, S.; Cartenì, M.; Nardone, G. Gut–liver axis: The impact of gut microbiota on non alcoholic fatty liver disease. Nutr. Metab. Cardiovas 2012, 22, 471–476. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Liu, L.; Sheikhahmadi, A.; Wang, Y.; Li, C.; Jiao, H.; Lin, H.; Song, Z. Effects of corticosterone and dietary energy on immune function of broiler chickens. PLoS ONE 2015, 10, e0119750. [Google Scholar] [CrossRef] [PubMed]
- Jirillo, E.; Caccavo, D.; Magrone, T.; Piccigallo, E.; Amati, L.; Lembo, A.; Kalis, C.; Gumenscheimer, M. The role of the liver in the response to LPS: Experimental and clinical findings. J. Endotoxin Res. 2002, 8, 319–327. [Google Scholar] [CrossRef]
- Triantafilou, M.; Triantafilou, K. Lipopolysaccharide recognition: CD14, TLRs and the LPS-activation cluster. Trends Immunol. 2002, 23, 301–304. [Google Scholar] [CrossRef]
- Jerala, R. Structural biology of the LPS recognition. Int. J. Med. Microbiol. 2007, 297, 353–363. [Google Scholar] [CrossRef]
- Verstrepen, L.; Bekaert, T.; Chau, T.-L.; Tavernier, J.; Chariot, A.; Beyaert, R. TLR-4, IL-1R and TNF-R signaling to NF-κB: Variations on a common theme. Cell. Mol. Life Sci. 2008, 65, 2964–2978. [Google Scholar] [CrossRef]
- Henao-Mejia, J.; Elinav, E.; Jin, C.; Hao, L.; Mehal, W.Z.; Strowig, T.; Thaiss, C.A.; Kau, A.L.; Eisenbarth, S.C.; Jurczak, M.J. Inflammasome-mediated dysbiosis regulates progression of NAFLD and obesity. Nature 2012, 482, 179–185. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.; Han, D.; Li, X.; Zhang, Y.; Zhao, Y. The impact of small doses of LPS on NASH in high sucrose and high fat diet induced rats. Eur. Rev. Med. Pharmacol. Sci. 2014, 18, 2742–2747. [Google Scholar]
- Sharifnia, T.; Antoun, J.; Verriere, T.G.; Suarez, G.; Wattacheril, J.; Wilson, K.T.; Peek, R.M., Jr.; Abumrad, N.N.; Flynn, C.R. Hepatic TLR4 signaling in obese NAFLD. Am. J. Physiol. Gastrointest. Liver Physiol. 2015, 309, G270–G278. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Ren, F.; Zhang, H.; Wen, T.; Piao, Z.; Zhou, L.; Zheng, S.; Zhang, J.; Chen, Y.; Han, Y. Inhibition of glycogen synthase kinase 3β ameliorates D-GalN/LPS-induced liver injury by reducing endoplasmic reticulum stress-triggered apoptosis. PLoS ONE 2012, 7, e45202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, L.; Dong, H.; Zhang, Z.; Liu, J.; Hu, Y.; Ni, Y.; Grossmann, R.; Zhao, R. Activation of epithelial proliferation induced by Eimeria acervulina infection in the duodenum may be associated with cholesterol metabolism. Oncotarget 2016, 7, 27627–27640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, Y.; Fukusato, T. Histopathology of nonalcoholic fatty liver disease/nonalcoholic steatohepatitis. World J. Gastroenterol. 2014, 20, 15539–15548. [Google Scholar] [CrossRef]
- Chalasani, N.; Wilson, L.; Kleiner, D.E.; Cummings, O.W.; Brunt, E.M.; Ünalp, A.; Network, N.C.R. Relationship of steatosis grade and zonal location to histological features of steatohepatitis in adult patients with non-alcoholic fatty liver disease. J. Hepatol. 2008, 48, 829–834. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Chen, X.; Huang, Z.; Chen, D.; He, J.; Zheng, P.; Chen, H.; Luo, J.; Luo, Y.; Yu, B. Effects of dietary apple polyphenols supplementation on hepatic fat deposition and antioxidant capacity in finishing pigs. Animals 2019, 9, 937. [Google Scholar] [CrossRef] [Green Version]
- Wu, T.; Yang, L.; Jiang, J.; Ni, Y.; Zhu, J.; Zheng, X.; Wang, Q.; Lu, X.; Fu, Z. Chronic glucocorticoid treatment induced circadian clock disorder leads to lipid metabolism and gut microbiota alterations in rats. Life Sci. 2018, 192, 173–182. [Google Scholar] [CrossRef]
- Lin, H.; Sui, S.; Jiao, H.; Buyse, J.; Decuypere, E. Impaired development of broiler chickens by stress mimicked by corticosterone exposure. Comp. Biochem. Phys. A 2006, 143, 400–405. [Google Scholar] [CrossRef]
- Zhang, H.; Chen, Y.; Chen, Y.; Li, Y.; Jia, P.; Ji, S.; Zhou, Y.; Wang, T. Dietary pterostilbene supplementation attenuates intestinal damage and immunological stress of broiler chickens challenged with lipopolysaccharide. J. Anim. Sci. 2019, 98. [Google Scholar] [CrossRef]
- Senior, J. Alanine aminotransferase: A clinical and regulatory tool for detecting liver injury—Past, present, and future. Clin. Pharmacol. Ther. 2012, 92, 332–339. [Google Scholar] [CrossRef]
- Chen, Z.; Yang, Y.; Mi, S.; Fan, Q.; Sun, X.; Deng, B.; Wu, G.; Li, Y.; Zhou, Q.; Ruan, Z. Hepatoprotective effect of chlorogenic acid against chronic liver injury in inflammatory rats. J. Funct. Foods 2019, 62, 103540. [Google Scholar] [CrossRef]
- Lim, J.W.; Dillon, J.; Miller, M. Proteomic and genomic studies of non-alcoholic fatty liver disease-clues in the pathogenesis. World J. Gastroenterol. 2014, 20, 8325–8340. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Liu, G.; Liang, X.; Wang, M.; Zhu, X.; Luo, Y.; Shang, Y.; Yang, J.; Zhou, P.; Gu, X.L. Effects of berberine on the growth performance, antioxidative capacity and immune response to lipopolysaccharide challenge in broilers. Anim. Sci. J. 2019, 90, 1229–1238. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wan, D.; Zhou, X.; Ruan, Z.; Zhang, T.; Wu, X.; Yin, Y. Effects of dynamic feeding low-and high-methionine diets on the variation of glucose and lipid metabolism-related genes in the liver of laying hens. Poult. Sci. 2019, 98, 2231–2240. [Google Scholar] [CrossRef] [PubMed]
- Valenti, L.; Fracanzani, A.L.; Fargion, S. The immunopathogenesis of alcoholic and nonalcoholic steatohepatitis: Two triggers for one disease? Semin. Immunopathol. 2009, 31, 359. [Google Scholar] [CrossRef]
- Bertola, A.; Bonnafous, S.; Anty, R.; Patouraux, S.; Saint-Paul, M.-C.; Iannelli, A.; Gugenheim, J.; Barr, J.; Mato, J.M.; Le Marchand-Brustel, Y. Hepatic expression patterns of inflammatory and immune response genes associated with obesity and NASH in morbidly obese patients. PLoS ONE 2010, 5, e13577. [Google Scholar] [CrossRef] [Green Version]
- Forn-Cuni, G.; Varela, M.; Fernandez-Rodriguez, C.M.; Figueras, A.; Novoa, B. Liver immune responses to inflammatory stimuli in a diet-induced obesity model of zebrafish. J. Endocrinol. 2015, 224, 159–170. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient % | Content |
|---|---|
| Corn | 58.80 |
| Soybean meal | 30.50 |
| Corn gluten meal | 5.00 |
| soybean oil | 1.42 |
| Calcium carbonate powder | 1.34 |
| Calcium hydrogen phosphate | 1.33 |
| Methionine | 0.18 |
| 65% Lysine | 0.45 |
| 93% Threonine | 0.05 |
| Sodium chloride | 0.30 |
| 50% Choline chloride | 0.30 |
| Chlortetracycline | 0.10 |
| Trace elements | 0.20 |
| Vitamins | 0.03 |
| Calculated Chemical Composition | |
| Metabolizable energy (MJ/kg) | 12.34 |
| Crude protein (g/kg) | 221.53 |
| Crude fat (g/kg) | 43.58 |
| Crude fiber (g/kg) | 27.90 |
| Calcium (g/kg) | 8.90 |
| Available P (g/kg) | 3.50 |
| Lysine (g/kg) | 12.91 |
| Methionine (g/kg) | 5.50 |
| Methionine + Cysteine (g/kg) | 9.17 |
| Target Genes | Genbank Accession | Primer Sequences (5′-3′) | PCR Products (bp) |
|---|---|---|---|
| TLR4 | NM_001030693 | F: AAACACCACCCTGGACTTGGAC | 188 |
| R: GTGCTGGAGTGAATTGGCAGTTA | |||
| MyD88 | NM_001030962 | F: CCTGGAAAGTGATGAATGTGA | 165 |
| R: TTGGTGCAAGGATTGGTGTA | |||
| NF-κB | NM_205134.1 | F: TCAACGCAGGACCTAAAGACAT | 162 |
| R: GCAGATAGCCAAGTTCAGGATG | |||
| INF-γ | X99774.1 | F: ACACTGACAAGTCAAAGCCGCACA | 129 |
| R: AGTCGTTCATCGGGAGCTTGGC | |||
| iNOS | D85422 | F: CCTGGAGGTCCTGGAAGAGT | 185 |
| R: CCTGGGTTTCAGAAGTGGC | |||
| IL-6 | AJ309541.1 | F: CGTGTGCGAGAACAGCATGGAGA | 130 |
| R: TCAGGCATTTCTCCTCGTCGAAGC | |||
| IL-1β | Y15006 | F: CATTCAGCGTCCAGGTA | 91 |
| R: TCTCGGCAGTTTCTTTT | |||
| IL-10 | NM_001004414.2 | F: AGCAGATCAAGGAGACGTTC | 103 |
| R: ATCAGCAGGTACTCCTCGAT | |||
| CRP | DQ374639.1 | F: TGTCACGGCCCAGGAAGA | 175 |
| R: TTTGGTGGCGTAGGAGAAGA | |||
| SAA | GU929209.1 | F: AGCGGTATCAAGTTTGTCAGG | 129 |
| R: TCCGAGCATCGTAATT | |||
| 18S | >AF173612.1 | F: GAAACGGCTACCACATCC | 168 |
| R: CACCAGACTTGCCCTCCA |
| Parameters | CON | CORT | LPS | LPS&CORT |
|---|---|---|---|---|
| TG (mmol/l) | 0.40 ± 0.03 b | 0.70 ± 0.10 a | 0.27 ± 0.02 c | 0.64 ± 0.14 a,b |
| TCHOL (mmol/l) | 2.67 ± 0.22 c | 5.66 ± 0.53 a | 2.24 ± 0.18 c | 4.23 ± 0.32 b |
| HDL (mmol/l) | 1.48 ± 0.12 c | 3.25 ± 0.27 a | 1.14 ± 0.12 c | 2.22 ± 0.17 b |
| LDL (mmol/l) | 0.28 ± 0.02 b | 0.53 ± 0.08 a | 0.39 ± 0.03 a,b | 0.44 ± 0.06 a |
| GLU (mmol/L) | 14.80 ± 0.31 | 14.80 ± 0.48 | 14.40 ± 0.64 | 15.81 ± 0.67 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mei, W.; Hao, Y.; Xie, H.; Ni, Y.; Zhao, R. Hepatic Inflammatory Response to Exogenous LPS Challenge is Exacerbated in Broilers with Fatty Liver Disease. Animals 2020, 10, 514. https://doi.org/10.3390/ani10030514
Mei W, Hao Y, Xie H, Ni Y, Zhao R. Hepatic Inflammatory Response to Exogenous LPS Challenge is Exacerbated in Broilers with Fatty Liver Disease. Animals. 2020; 10(3):514. https://doi.org/10.3390/ani10030514
Chicago/Turabian StyleMei, Wenqing, Yarong Hao, Huilin Xie, Yingdong Ni, and Ruqian Zhao. 2020. "Hepatic Inflammatory Response to Exogenous LPS Challenge is Exacerbated in Broilers with Fatty Liver Disease" Animals 10, no. 3: 514. https://doi.org/10.3390/ani10030514