Running Head: Heat Affects Cholesterol and Bile Acid Alterations in Cholesterol and Bile Acids Metabolism in Large White Pigs during Short-Term Heat Exposure

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Study Design
2.2. Serum Lipid Profiles
2.3. Hepatic Lipid Analyses
2.4. Bile Acids Extraction and Quantification
2.5. Isolation of RNA and Quantification of mRNA Levels in the Liver
2.6. Statistical Analysis
3. Results
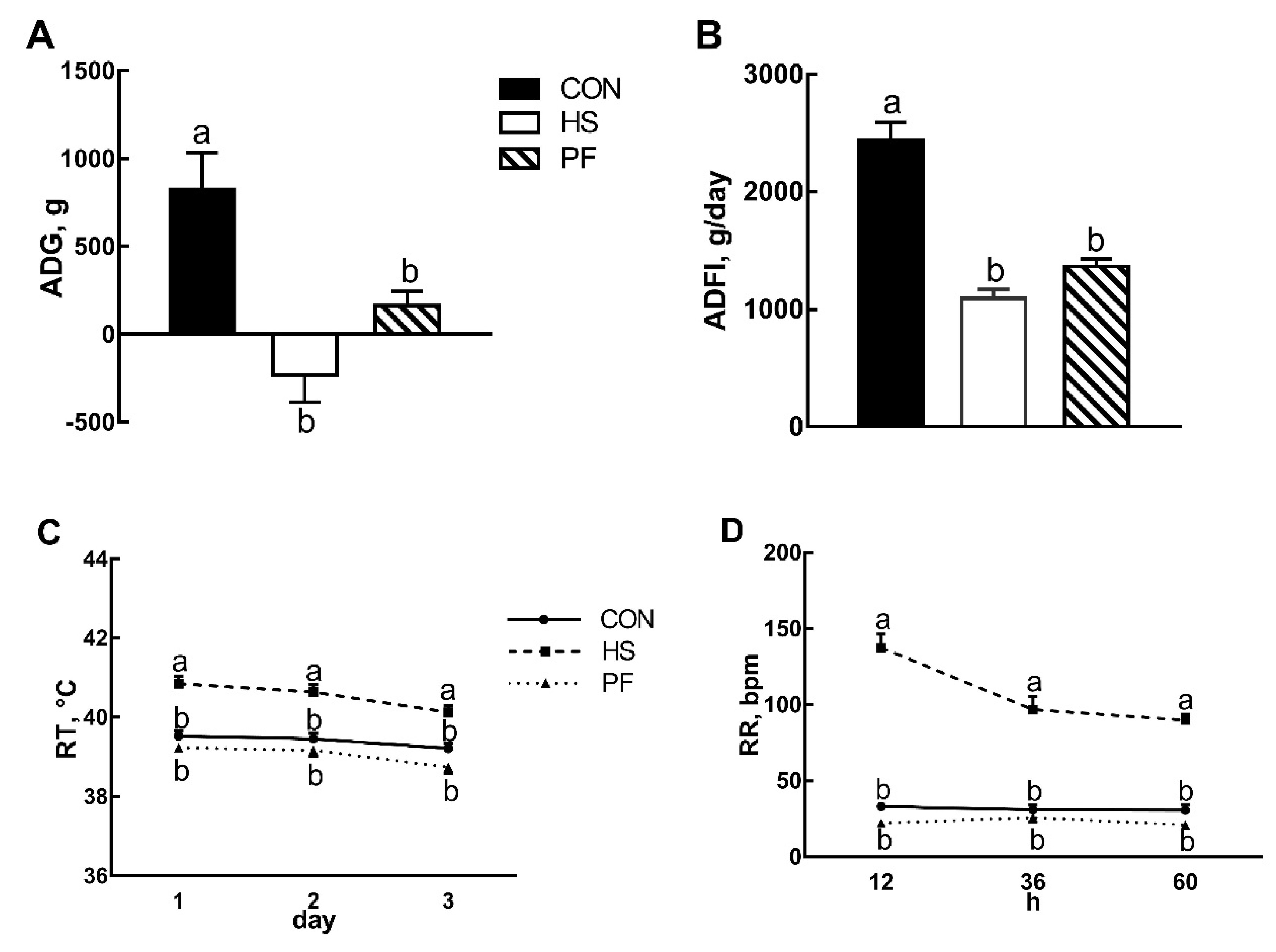
3.1. Pig Performances after Short Term Heat Stress
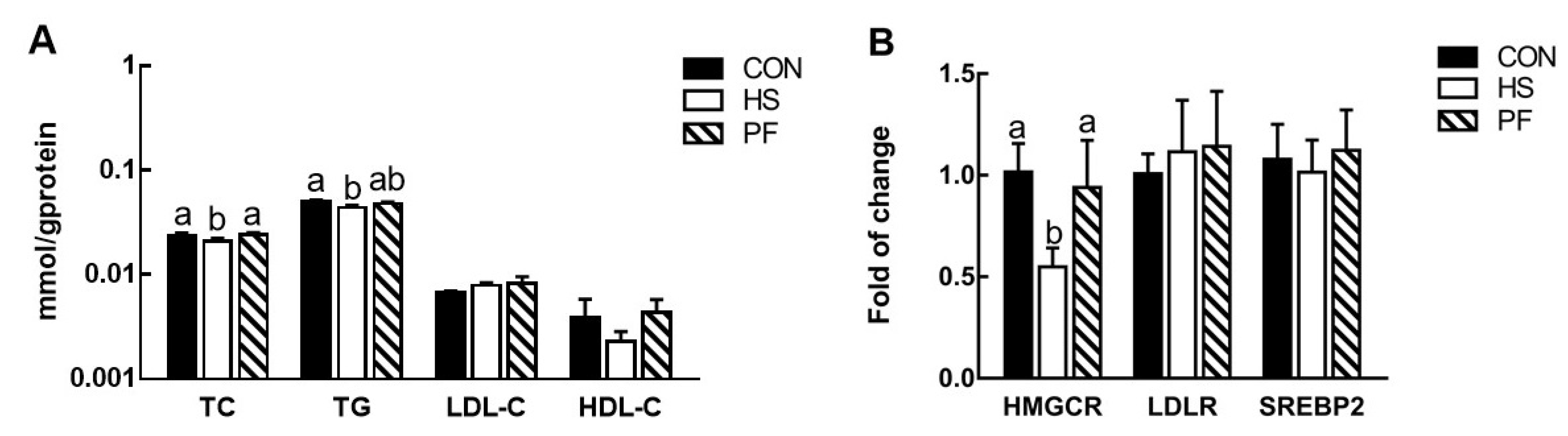
3.2. Serum Lipid Levels
3.3. Changes in Liver Cholesterol Biosynthesis
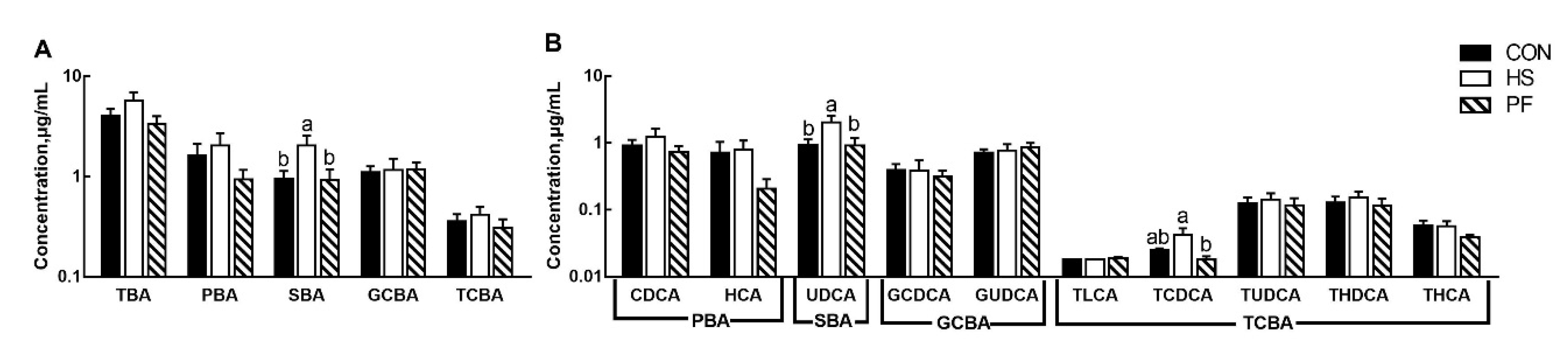
3.4. Serum BAs Composition after Short Term Heat Stress
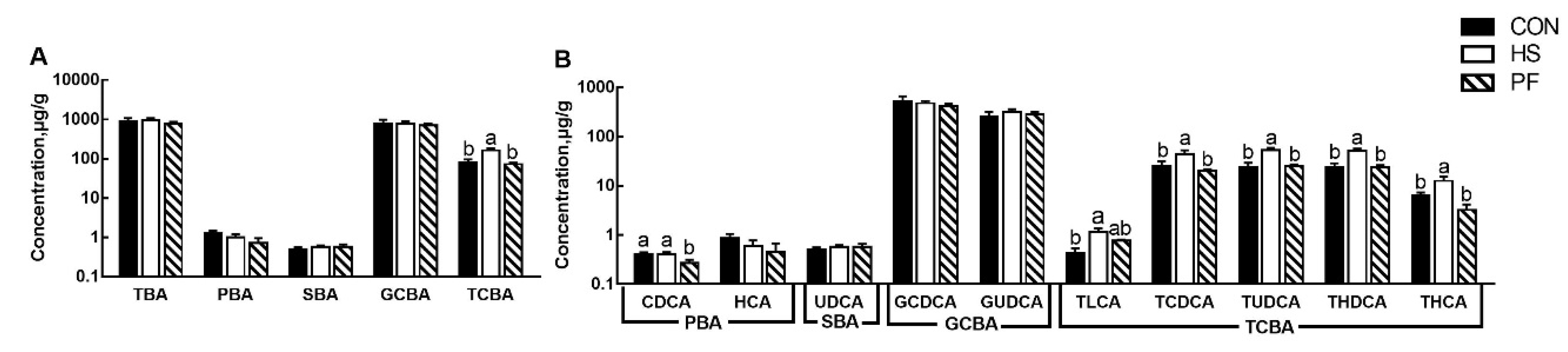
3.5. Hepatic BAs Composition after Short Term Heat Stress
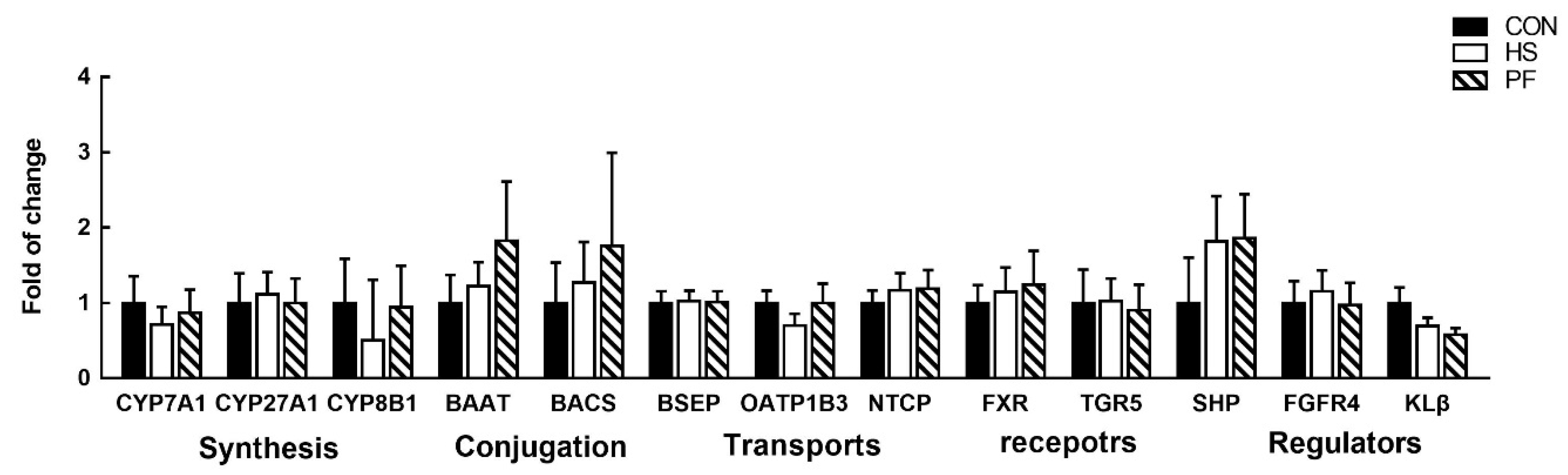
3.6. Bile Acids Synthesis, Transporters, and Regulation in the Liver
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- St-Pierre, N.; Cobanov, B.; Schnitkey, G. Economic losses from heat stress by US livestock industries1. J. Dairy Sci. 2003, 86, E52–E77. [Google Scholar] [CrossRef] [Green Version]
- Hyun, Y.; Ellis, M.; Riskowski, G.; Johnson, R. Growth performance of pigs subjected to multiple concurrent environmental stressors. J. Anim. Sci. 1998, 76, 721–727. [Google Scholar] [CrossRef]
- Renaudeau, D.; Anais, C.; Tel, L.; Gourdine, J. Effect of temperature on thermal acclimation in growing pigs estimated using a nonlinear function1. J. Anim. Sci. 2010, 88, 3715–3724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collin, A.; van Milgent, J.; Le Dividich, J. Modelling the effect of high, constant temperature on food intake in young growing pigs. Anim. Sci. 2001, 72, 519–527. [Google Scholar] [CrossRef]
- Baumgard, L.H.; Rhoads, R.P., Jr. Effects of heat stress on postabsorptive metabolism and energetics. Annu. Rev. Anim. Biosci. 2013, 1, 311–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pearce, S.; Gabler, N.; Ross, J.; Escobar, J.; Patience, J.; Rhoads, R.; Baumgard, L. The effects of heat stress and plane of nutrition on metabolism in growing pigs. J. Anim. Sci. 2013, 91, 2108–2118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qu, H.; Donkin, S.; Ajuwon, K. Heat stress enhances adipogenic differentiation of subcutaneous fat depot–derived porcine stromovascular cells. J. Anim. Sci. 2015, 93, 3832–3842. [Google Scholar] [CrossRef] [Green Version]
- Belhadj Slimen, I.; Najar, T.; Ghram, A.; Abdrrabba, M. Heat stress effects on livestock: Molecular, cellular and metabolic aspects, a review. J. Anim. Physiol. Anim. Nutr. 2016, 100, 401–412. [Google Scholar] [CrossRef] [Green Version]
- Kouba, M.; Hermier, D.; Le Dividich, J. Influence of a high ambient temperature on lipid metabolism in the growing pig. J. Anim. Sci. 2001, 79, 81–87. [Google Scholar] [CrossRef]
- Qu, H.; Ajuwon, K. Adipose tissue-specific responses reveal an important role of lipogenesis during heat stress adaptation in pigs. J. Anim. Sci. 2018, 96, 975–989. [Google Scholar] [CrossRef] [Green Version]
- Qu, H.; Ajuwon, K.M. Metabolomics of heat stress response in pig adipose tissue reveals alteration of phospholipid and fatty acid composition during heat stress. J. Anim. Sci. 2018, 96, 3184–3195. [Google Scholar] [CrossRef] [PubMed]
- Vigh, L.; Maresca, B.; Harwood, J.L. Does the membrane’s physical state control the expression of heat shock and other genes? Trends Biochem. Sci. 1998, 23, 369–374. [Google Scholar] [CrossRef]
- Leach, M.D.; Cowen, L.E. Membrane fluidity and temperature sensing are coupled via circuitry comprised of Ole1, Rsp5, and Hsf1 in Candida albicans. Eukaryot. Cell 2014, 13, 1077–1084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagy, E.; Balogi, Z.; Gombos, I.; Åkerfelt, M.; Björkbom, A.; Balogh, G.; Török, Z.; Maslyanko, A.; Fiszer-Kierzkowska, A.; Lisowska, K. Hyperfluidization-coupled membrane microdomain reorganization is linked to activation of the heat shock response in a murine melanoma cell line. Proc. Natl. Acad. Sci. USA 2007, 104, 7945–7950. [Google Scholar] [CrossRef] [Green Version]
- Balogh, G.; Péter, M.; Glatz, A.; Gombos, I.; Török, Z.; Horváth, I.; Harwood, J.L.; Vígh, L. Key role of lipids in heat stress management. FEBS Lett. 2013, 587, 1970–1980. [Google Scholar] [CrossRef] [Green Version]
- Sudhof, T.C.; Goldstein, J.L.; Brown, M.S.; Russell, D.W. The LDL receptor gene: A mosaic of exons shared with different proteins. Science 1985, 228, 815–822. [Google Scholar] [CrossRef] [Green Version]
- Lusis, A.; Heinzmann, C.; Sparkes, R.; Scott, J.; Knott, T.; Geller, R.; Sparkes, M.; Mohandas, T. Regional mapping of human chromosome 19: Organization of genes for plasma lipid transport (APOC1,-C2, and-E and LDLR) and the genes C3, PEPD, and GPI. Proc. Natl. Acad. Sci. USA 1986, 83, 3929–3933. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.B.; Kan, C.F.K.; Shende, V.; Dong, B.; Liu, J. A novel posttranscriptional mechanism for dietary cholesterol-mediated suppression of liver LDL receptor expression. J. Lipid Res. 2014, 55, 1397–1407. [Google Scholar] [CrossRef] [Green Version]
- Horton, J.D.; Goldstein, J.L.; Brown, M.S. SREBPs: Activators of the complete program of cholesterol and fatty acid synthesis in the liver. J. Clin. Investig. 2002, 109, 1125–1131. [Google Scholar] [CrossRef]
- Lu, T.T.; Makishima, M.; Repa, J.J.; Schoonjans, K.; Kerr, T.A.; Auwerx, J.; Mangelsdorf, D.J. Molecular basis for feedback regulation of bile acid synthesis by nuclear receptors. Mol. Cell 2000, 6, 507–515. [Google Scholar] [CrossRef]
- Dawson, P.A.; Karpen, S.J. Intestinal Transport and Metabolism of Bile Acids. J. Lipid Res. 2015, 56, 1085–1099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russell, D.W. The enzymes, regulation, and genetics of bile acid synthesis. Annu. Rev. Biochem. 2003, 72, 137–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collier, R.J.; Baumgard, L.H.; Zimbelman, R.B.; Xiao, Y. Heat stress: Physiology of acclimation and adaptation. Anim. Front. 2018, 9, 12–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wen, X.; Wu, W.; Fang, W.; Tang, S.; Xin, H.; Xie, J.; Zhang, H. Effects of long-term heat exposure on cholesterol metabolism and immune responses in growing pigs. Livest. Sci. 2019, 230, 103857. [Google Scholar] [CrossRef]
- Fang, W.; Wen, X.; Meng, Q.; Wu, W.; Everaert, N.; Xie, J.; Zhang, H. Alteration in bile acids profile in Large White pigs during chronic heat exposure. J. Therm. Biol. 2019, 84, 375–383. [Google Scholar] [CrossRef]
- Pearce, S.; Sanz-Fernandez, M.; Hollis, J.; Baumgard, L.H.; Gabler, N.K. Short-term exposure to heat stress attenuates appetite and intestinal integrity in growing pigs. J. Anim. Sci. 2014, 92, 5444–5454. [Google Scholar] [CrossRef] [Green Version]
- Seelenbinder, K.M.; Zhao, L.D.; Hanigan, M.D.; Hulver, M.W.; McMillan, R.P.; Baumgard, L.H.; Selsby, J.T.; Ross, J.W.; Gabler, N.K.; Rhoads, R.P. Effects of heat stress during porcine reproductive and respiratory syndrome virus infection on metabolic responses in growing pigs. J. Anim. Sci. 2018, 96, 1375–1387. [Google Scholar] [CrossRef]
- Konings, A. Membranes as targets for hyperthermic cell killing. In Preclinical Hyperthermia; Springer: Berlin/Heidelberg, Germany, 1988; pp. 9–21. [Google Scholar]
- Crockett, E.L. Cholesterol function in plasma membranes from ectotherms: Membrane-specific roles in adaptation to temperature. Am. Zool. 1998, 38, 291–304. [Google Scholar] [CrossRef] [Green Version]
- Cress, A.E.; Culver, P.S.; Moon, T.E.; Gerner, E.W. Correlation between amounts of cellular membrane components and sensitivity to hyperthermia in a variety of mammalian cell lines in culture. Cancer Res. 1982, 42, 1716–1721. [Google Scholar]
- Cress, A.E.; Gerner, E.W. Cholesterol levels inversely reflect the thermal sensitivity of mammalian cells in culture. Nature 1980, 283, 677. [Google Scholar] [CrossRef]
- Kirkegaard, T.; Roth, A.G.; Petersen, N.H.; Mahalka, A.K.; Olsen, O.D.; Moilanen, I.; Zylicz, A.; Knudsen, J.; Sandhoff, K.; Arenz, C. Hsp70 stabilizes lysosomes and reverts Niemann–Pick disease-associated lysosomal pathology. Nature 2010, 463, 549. [Google Scholar] [CrossRef] [PubMed]
- Feder, M.E.; Hofmann, G.E. Heat-shock proteins, molecular chaperones, and the stress response: Evolutionary and ecological physiology. Annu. Rev. Physiol. 1999, 61, 243–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ness, G.C.; Chambers, C.M. Feedback and hormonal regulation of hepatic 3-hydroxy-3-methylglutaryl coenzyme A reductase: The concept of cholesterol buffering capacity. Proc. Soc. Exp. Biol. Med. 2000, 224, 8–19. [Google Scholar] [CrossRef] [PubMed]
- Hardison, W.; Proffitt, J.H. Influence of hepatic taurine concentration on bile acid conjugation with taurine. Am. J. Physiol. Endocrinol. Metab. 1977, 232, E75. [Google Scholar] [CrossRef] [PubMed]
- Hardison, W.G. Hepatic taurine concentration and dietary taurine as regulators of bile acid conjugation with taurine. Gastroenterology 1978, 75, 71–75. [Google Scholar] [CrossRef]
- Sjövall, J. Dietary glycine and taurine on bile acid conjugation in man. Bile acids and steroids 75. Proc. Soc. Exp. Biol. Med. 1959, 100, 676–678. [Google Scholar] [CrossRef]
- Hahn, G. Dynamic responses of cattle to thermal heat loads. J. Anim. Sci. 1999, 77, 10–20. [Google Scholar] [CrossRef]
- Frosini, M.; Sesti, C.; Palmi, M.; Valoti, M.; Fusi, F.; Mantovani, P.; Bianchi, L.; Della Corte, L.; Sgaragli, G. The possible role of taurine and GABA as endogenous cryogens in the rabbit. In Taurine 4; Springer: Berlin/Heidelberg, Germany, 2002; pp. 335–344. [Google Scholar]
- El Idrissi, A.; Trenkner, E. Growth factors and taurine protect against excitotoxicity by stabilizing calcium homeostasis and energy metabolism. J. Neurosci. 1999, 19, 9459–9468. [Google Scholar] [CrossRef]
- Yang, J.; Zong, X.; Wu, G.; Lin, S.; Feng, Y.; Hu, J. Taurine increases testicular function in aged rats by inhibiting oxidative stress and apoptosis. Amino Acids 2015, 47, 1549–1558. [Google Scholar] [CrossRef]
- Surai, P.; Kochish, I.; Kidd, M. Taurine in poultry nutrition. Anim. Feed Sci. Technol. 2019, 260, 114339. [Google Scholar] [CrossRef]
- Serviddio, G.; Pereda, J.; Pallardó, F.V.; Carretero, J.; Borras, C.; Cutrin, J.; Vendemiale, G.; Poli, G.; Viña, J.; Sastre, J. Ursodeoxycholic acid protects against secondary biliary cirrhosis in rats by preventing mitochondrial oxidative stress. Hepatology 2004, 39, 711–720. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, C.; Fan, G.; Wong, P.Y.; Kren, B.T.; Steer, C.J. Ursodeoxycholic acid may inhibit deoxycholic acid-induced apoptosis by modulating mitochondrial transmembrane potential and reactive oxygen species production. Mol. Med. 1998, 4, 165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hofmann, A.F. The enterohepatic circulation of bile acids in mammals: Form and functions. Front Biosci. 2009, 14, 2584–2598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watt, S.M.; Simmonds, W.J. Effects of four taurine-conjugated bile acids on mucosal uptake and lymphatic absorption of cholesterol in the rat. J. Lipid Res. 1984, 25, 448–455. [Google Scholar]
- Murakami, S.; Fujita, M.; Nakamura, M.; Sakono, M.; Nishizono, S.; Sato, M.; Imaizumi, K.; Mori, M.; Fukuda, N. Taurine ameliorates cholesterol metabolism by stimulating bile acid production in high-cholesterol-fed rats. Clin. Exp. Pharmacol. Physiol. 2016, 43, 372–378. [Google Scholar] [CrossRef]
- Yu, J.; Yin, P.; Liu, F.; Cheng, G.; Guo, K.; Lu, A.; Zhu, X.; Luan, W.; Xu, J. Effect of heat stress on the porcine small intestine: A morphological and gene expression study. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2010, 156, 119–128. [Google Scholar] [CrossRef]
- Zhang, M.; Bi, L.; Fang, J.; Su, X.; Da, G.; Kuwamori, T.; Kagamimori, S. Beneficial effects of taurine on serum lipids in overweight or obese non-diabetic subjects. Amino Acids 2004, 26, 267–271. [Google Scholar] [CrossRef]
- Yeh, Y.-H.; Chen, M.-H.; Lee, Y.-T.; Hsieh, H.-S.; Hwang, D.-F. Effect of Taurine on Toxicity of Oxidized Cholesterol and Oxidized Fish Oil in Rats. J. Food Drug Anal. 2008, 16, 76–85. [Google Scholar]
- Lu, Z.; He, X.; Ma, B.; Zhang, L.; Li, J.; Jiang, Y.; Zhou, G.; Gao, F. Dietary taurine supplementation decreases fat synthesis by suppressing the liver X receptor α pathway and alleviates lipid accumulation in the liver of chronic heat-stressed broilers. J. Sci. Food Agric. 2019. [Google Scholar] [CrossRef]
- Morales, A.; Cota, S.; Ibarra, N.; Arce, N.; Htoo, J.; Cervantes, M. Effect of heat stress on the serum concentrations of free amino acids and some of their metabolites in growing pigs. J. Anim. Sci. 2016, 94, 2835–2842. [Google Scholar] [CrossRef]
- Gabler, N.K.; Koltes, D.; Schaumberger, S.; Murugesan, G.R.; Reisinger, N. Diurnal heat stress reduces pig intestinal integrity and increases endotoxin translocation. Transl. Anim. Sci. 2018, 2, 1–10. [Google Scholar] [CrossRef]
- Sun, X.; Zhang, H.; Sheikhahmadi, A.; Wang, Y.; Jiao, H.; Lin, H.; Song, Z. Effects of heat stress on the gene expression of nutrient transporters in the jejunum of broiler chickens (Gallus gallus domesticus). Int. J. Biometeorol. 2015, 59, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Xin, H.; Zhang, X.; Sun, D.; Zhang, C.; Hao, Y.; Gu, X. Chronic heat stress increases insulin-like growth factor-1 (IGF-1) but does not affect IGF-binding proteins in growing pigs. J. Therm. Biol. 2018, 77, 122–130. [Google Scholar] [CrossRef] [PubMed]
- Oded, S.; Ru, W.; Wang, T.J.; Catherine, R.; Lewis, G.D.; Vasan, R.S.; Carr, S.A.; Ravi, T.; Gerszten, R.E.; Mootha, V.K. Metabolic profiling of the human response to a glucose challenge reveals distinct axes of insulin sensitivity. Mol. Syst. Biol. 2008, 4, 214. [Google Scholar]
- Fu, Z.D.; Klaassen, C.D. Increased bile acids in enterohepatic circulation by short-term calorie restriction in male mice. Toxicol. Appl. Pharmacol. 2013, 273, 680–690. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Value, % |
|---|---|
| Corn | 62.55 |
| Soybean meal | 27.00 |
| Wheat bran | 5.00 |
| Soybean oil | 1.00 |
| Limestone | 1.50 |
| CaHPO4 | 1.50 |
| NaCl | 0.30 |
| L-Lysine-HCl | 0.15 |
| Vitamin and mineral premix (1) | 1.00 |
| Nutrients (2) | |
| DE (cal/kg) | 3230 |
| CP | 18.00 |
| Lys | 1.03 |
| Ca | 0.95 |
| P | 0.74 |
| Items | CON | HS | PF | SEM | p-Value |
|---|---|---|---|---|---|
| TC, mmol/L | 1.97 b | 2.26 a | 1.77 b | 0.19 | 0.009 |
| HDL-C, mmol/L | 0.57 | 0.52 | 0.50 | 0.10 | 0.302 |
| LDL-C, mmol/L | 1.12 b | 1.43 a | 0.99 b | 0.17 | 0.002 |
| TG, mmol/L | 0.41 b | 0.66 a | 0.34 b | 0.14 | 0.002 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fang, W.; Wen, X.; Meng, Q.; Liu, L.; Xie, J.; Zhang, H.; Everaert, N. Running Head: Heat Affects Cholesterol and Bile Acid Alterations in Cholesterol and Bile Acids Metabolism in Large White Pigs during Short-Term Heat Exposure. Animals 2020, 10, 359. https://doi.org/10.3390/ani10020359
Fang W, Wen X, Meng Q, Liu L, Xie J, Zhang H, Everaert N. Running Head: Heat Affects Cholesterol and Bile Acid Alterations in Cholesterol and Bile Acids Metabolism in Large White Pigs during Short-Term Heat Exposure. Animals. 2020; 10(2):359. https://doi.org/10.3390/ani10020359
Chicago/Turabian StyleFang, Wei, Xiaobin Wen, Qingshi Meng, Lei Liu, Jingjing Xie, Hongfu Zhang, and Nadia Everaert. 2020. "Running Head: Heat Affects Cholesterol and Bile Acid Alterations in Cholesterol and Bile Acids Metabolism in Large White Pigs during Short-Term Heat Exposure" Animals 10, no. 2: 359. https://doi.org/10.3390/ani10020359