Rumen-Protected Glucose Stimulates the Insulin-Like Growth Factor System and mTOR/AKT Pathway in the Endometrium of Early Postpartum Dairy Cows


Abstract
:Simple Summary
Abstract
1. Introduction
2. Methods and Materials
2.1. Animals and Experimental Procedures
2.2. Sample Collection
2.3. Immunofluorescence Double-Labeling
2.4. Insulin-Like Growth Factor 1 Assay
2.5. Total RNA Extraction and cDNA Synthesis
2.6. RT-PCR Analysis
2.7. Western Blot Analysis
2.8. Histological and Immunohistochemical Assessment
2.9. Statistical Analysis
3. Results
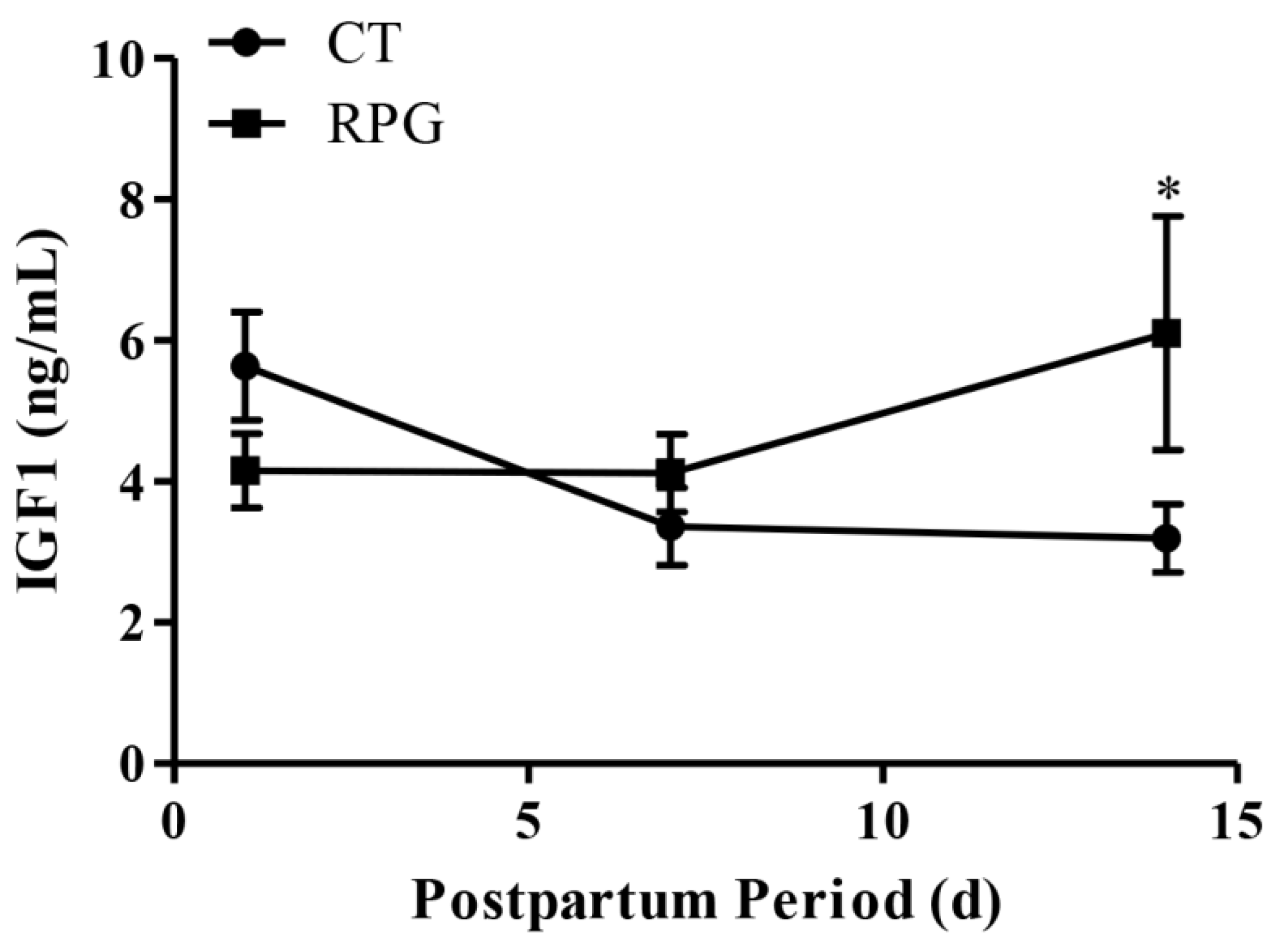
3.1. Body and Uterine Weight, Plasma IGF1 Concentration
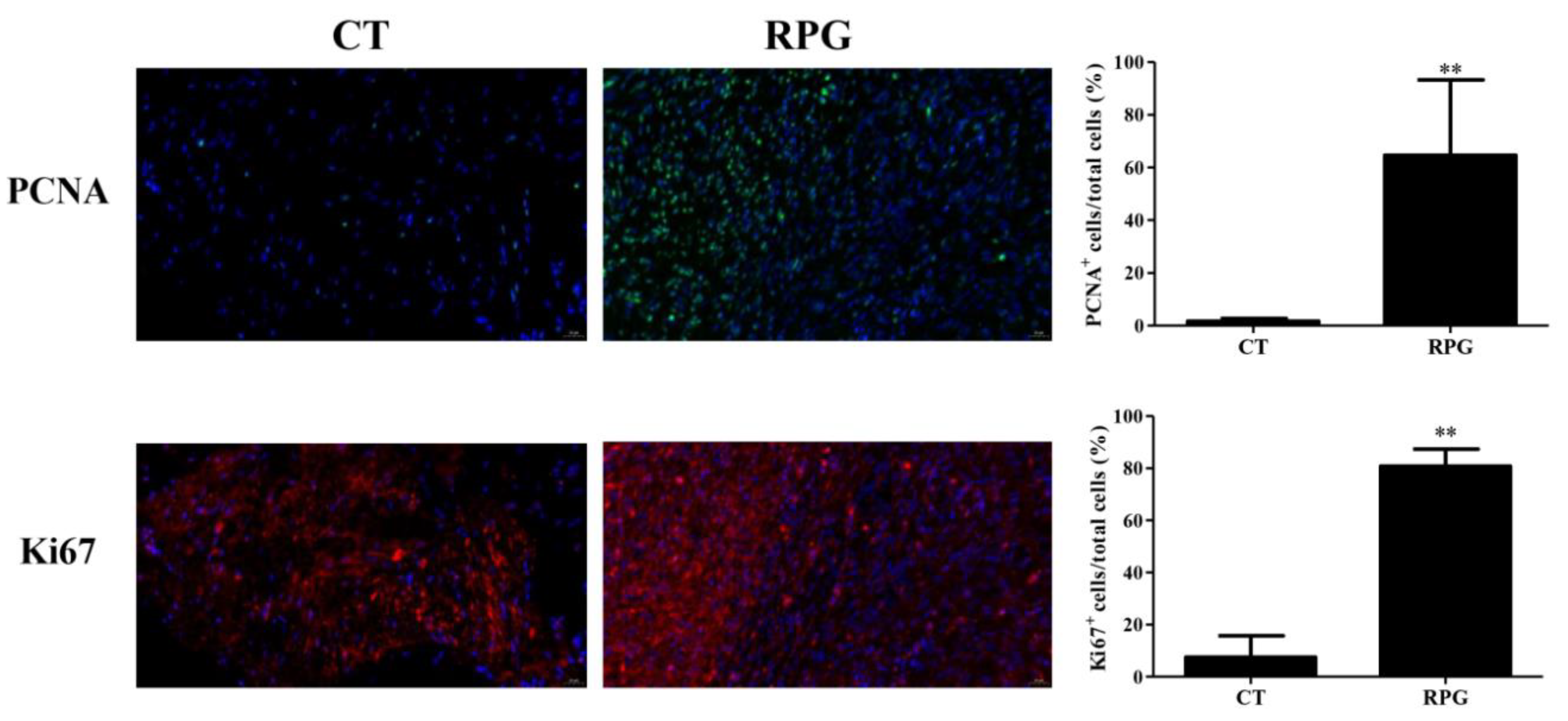
3.2. Proliferation Status in the Uterus
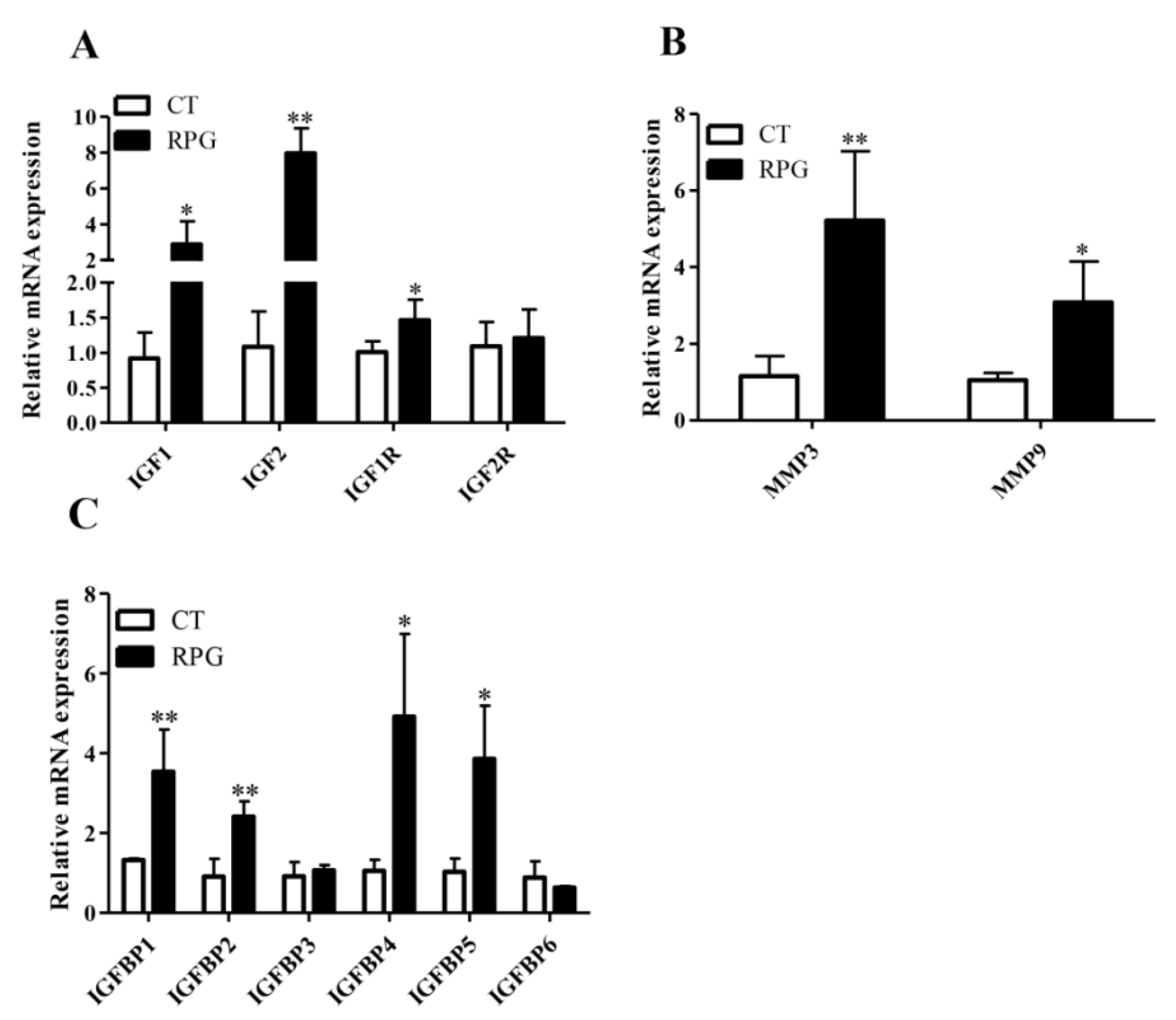
3.3. Gene Expression Profile
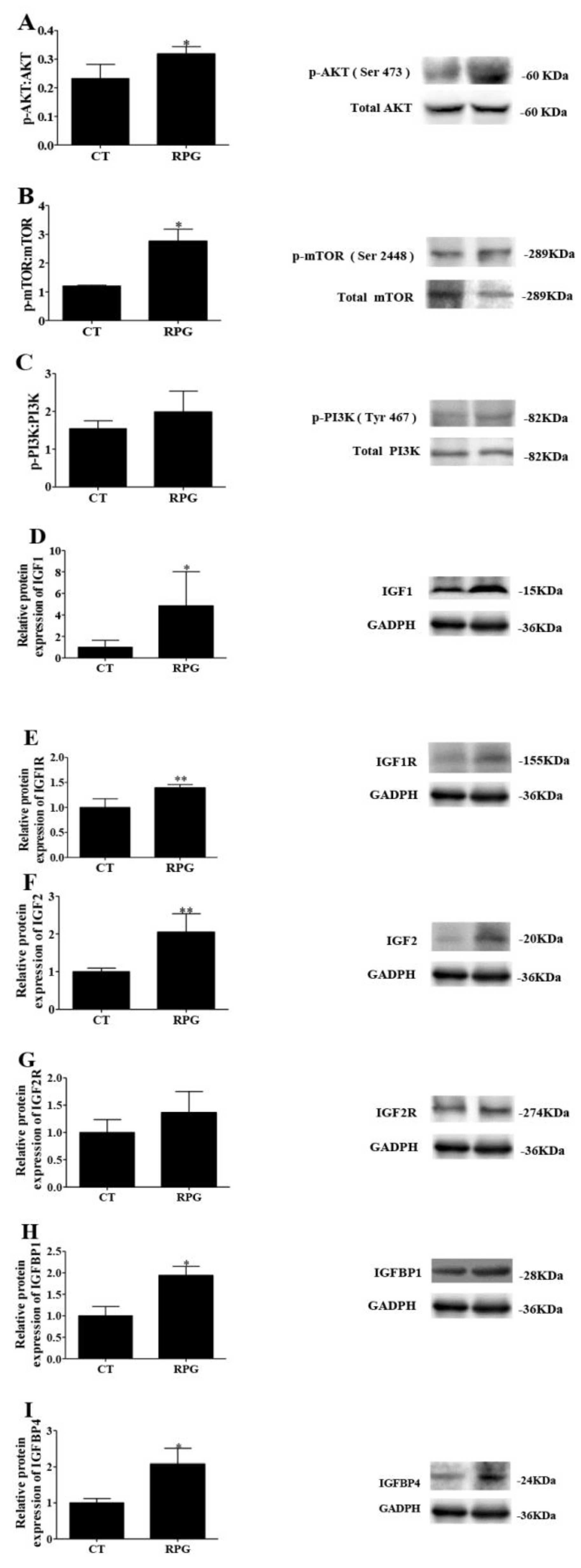
3.4. Functional Protein Expression Pattern
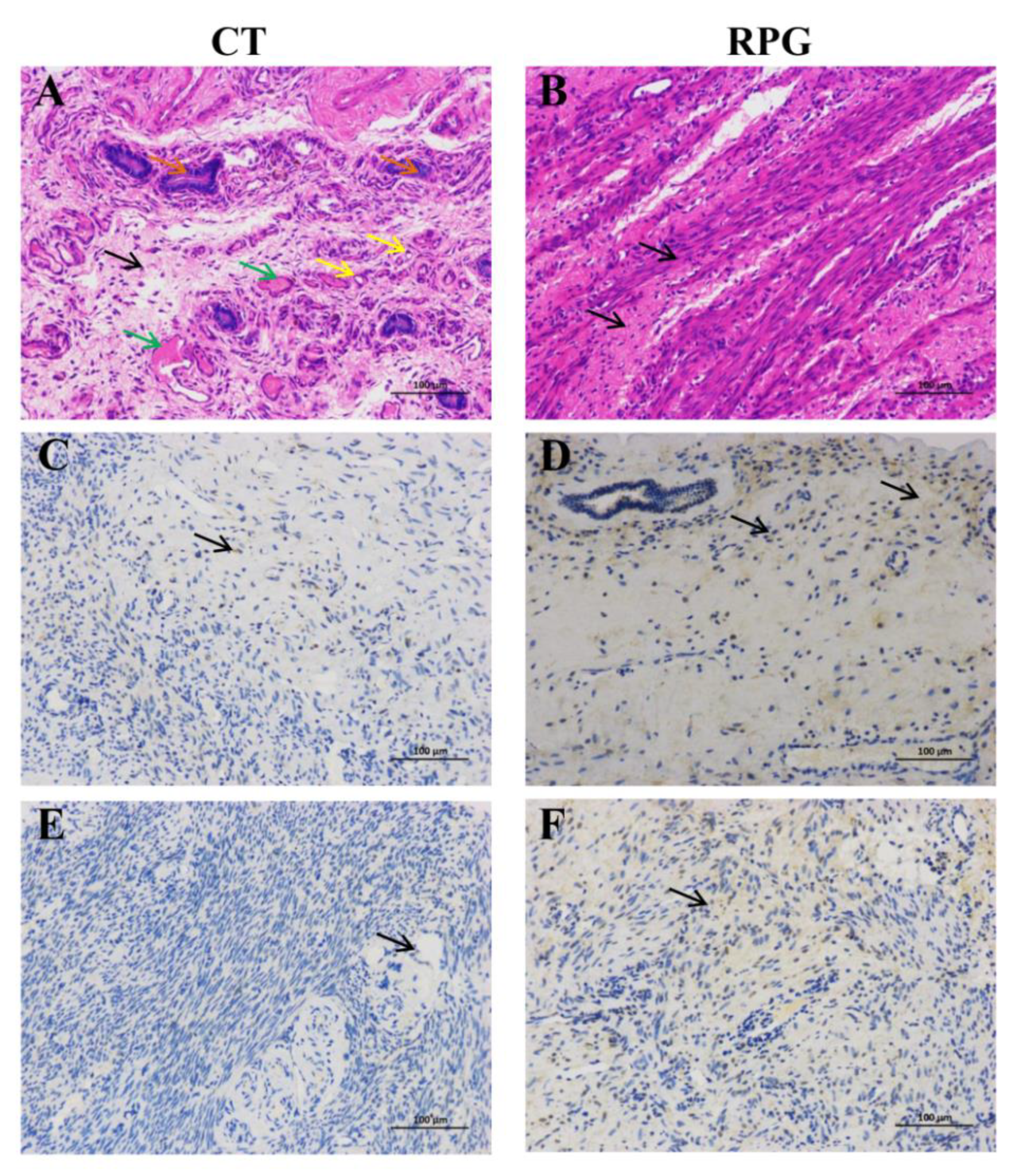
3.5. Histopathologic Changes and Immunohistochemical Localization of IGF1R and IGF2R in the Uterus
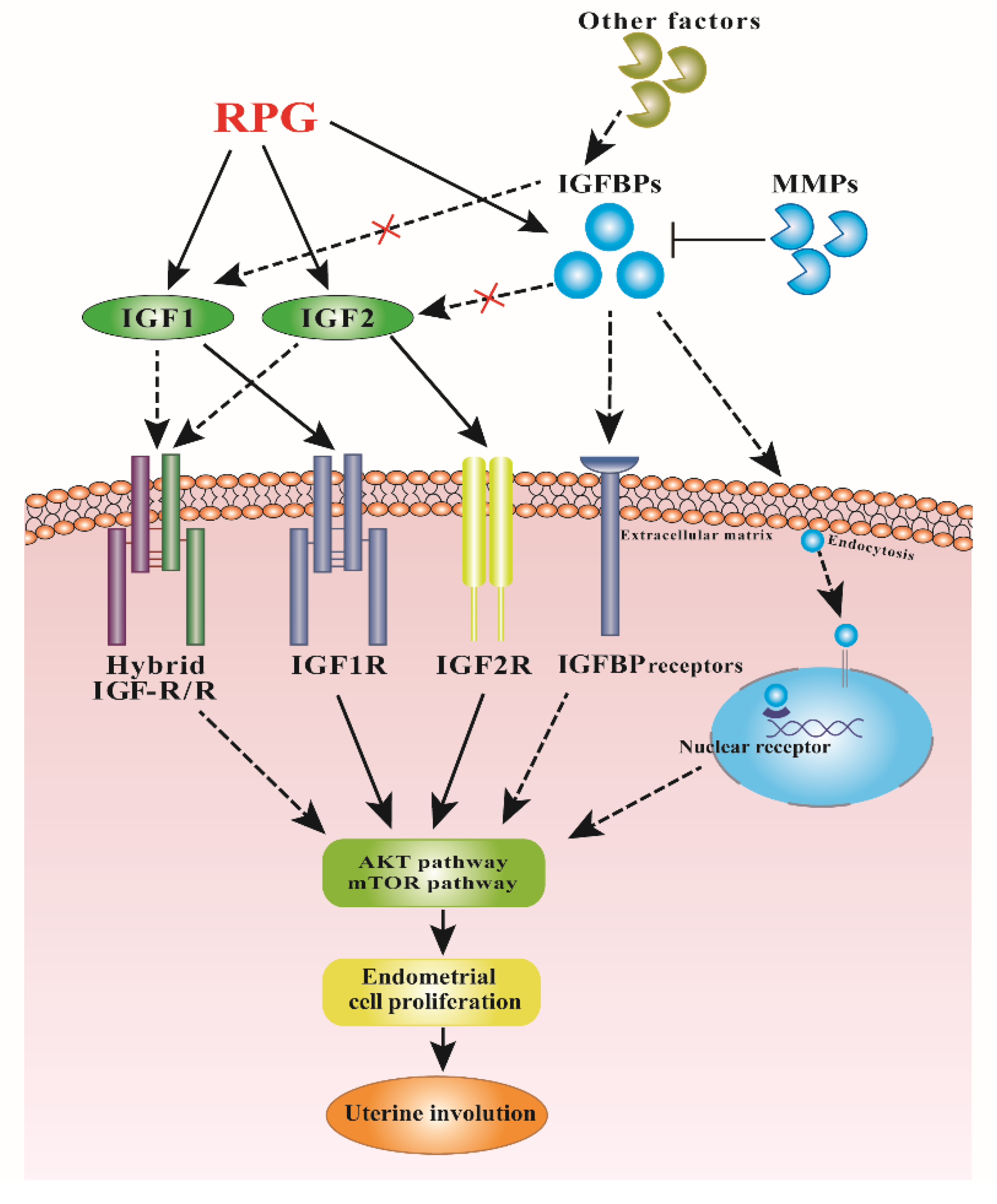
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lucy, M.C.; Butler, S.T.; Garverick, H.A. Endocrine and metabolic mechanisms linking postpartum glucose with early embryonic and foetal development in dairy cows. Animal 2014, 8, 82–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bell, A.W. Regulation of organic nutrient metabolism during transition from late pregnancy to early lactation. J. Anim. Sci. 1995, 73, 2804–2819. [Google Scholar] [CrossRef] [PubMed]
- Wathes, D.C.; Cheng, Z.; Bourne, N.; Taylor, V.J.; Coffey, M.P.; Brotherstone, S. Differences between primiparous and multiparous dairy cows in the inter-relationships between metabolic traits, milk yield and body condition score in the periparturient period. Domest. Anim. Endocrinol. 2007, 33, 203–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valour, D.; Hue, I.; Degrelle, S.A.; Dejean, S.; Marot, G.; Dubois, O.; Germain, G.; Humblot, P.; Ponter, A.A.; Charpigny, G.; et al. Pre-and post-partum mild underfeeding influences gene expression in the reproductive tract of cyclic dairy cows. Reprod. Domest. Anim. 2013, 48, 484–499. [Google Scholar] [CrossRef] [PubMed]
- Salamonsen, L.A. Tissue injury and repair in the female human reproductive tract. Reproduction 2003, 125, 301–311. [Google Scholar] [CrossRef] [PubMed]
- Llewellyn, S.; Fitzpatrick, R.; Kenny, D.A.; Patton, J.; Wathes, D.C. Endometrial expression of the insulin-like growth factor system during uterine involution in the postpartum dairy cow. Domest. Anim. Endocrinol. 2008, 34, 391–402. [Google Scholar] [CrossRef] [Green Version]
- Clemmons, D.R. Role of insulin-like growth factor binding proteins in controlling IGF actions. Mol. Cell. Endocrinol. 1998, 140, 19–24. [Google Scholar] [CrossRef]
- Taipale, J.; KeskiOja, J. Growth factors in the extracellular matrix. FASEB J. 1997, 11, 51–59. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, L.; Liu, Y.; Cui, J.; Che, S.; An, X.; Song, Y.; Cao, B. Circ-8073 regulates CEP55 by sponging miR-449a to promote caprine endometrial epithelial cells proliferation via the PI3K/AKT/mTOR pathway. Biochim. Biophys. Acta Mol. Cell Res. 2018, 1865, 1130–1147. [Google Scholar] [CrossRef]
- Hindle, V.A.; Vuuren van, A.M.; Klop, A.; Mathijssen-Kamman, A.A.; van Gelder, A.H.; Cone, J.W. Site and extent of starch degradation in the dairy cow-a comparison between in vivo, in situ and in vitro measurements. J. Anim. Physiol. Anim. Nutr. (Berl) 2005, 89, 158–165. [Google Scholar] [CrossRef]
- Li, X.P.; Tan, Z.L.; Jiao, J.Z.; Long, D.L.; Zhou, C.S.; Yi, K.L.; Liu, C.H.; Kang, J.H.; Wang, M.; Duan, F.H.; et al. Supplementation with fat-coated rumen-protected glucose during the transition period enhances milk production and influences blood biochemical parameters of liver function and inflammation in dairy cows. Anim. Feed Sci. Technol. 2019, 252, 92–102. [Google Scholar] [CrossRef]
- Oh, Y.S.; Koh, I.K.; Choi, B.; Gye, M.C. ESR1 inhibits hCG-induced steroidogenesis and proliferation of progenitor Leydig cells in mice. Sci. Rep. 2017, 7, 43459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, F.; Chong Lee Shin, O.L.S.; Pei, G.; Hu, Z.; Yang, J.; Zhu, H.; Wang, M.; Mou, J.; Sun, J.; Wang, Y.; et al. Adipose-derived mesenchymal stem cells employed exosomes to attenuate AKI-CKD transition through tubular epithelial cell dependent Sox9 activation. Oncotarget 2017, 8, 70707–70726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiao, J.; Wu, J.; Wang, M.; Zhou, C.; Zhong, R.; Tan, Z. Rhubarb supplementation promotes intestinal mucosal innate immune homeostasis through modulating intestinal epithelial microbiota in goat kids. J. Agric. Food Chem. 2018, 66, 1047–1057. [Google Scholar] [CrossRef] [PubMed]
- Wathes, D.C.; Cheng, Z.R.; Fenwick, M.A.; Fitzpatrick, R.; Patton, J. Influence of energy balance on the somatotrophic axis and matrix metalloproteinase expression in the endometrium of the postpartum dairy cow. Reproduction 2011, 141, 269–281. [Google Scholar] [CrossRef] [Green Version]
- De Cesaro, M.P.; Dos Santos, J.T.; Ferst, J.G.; Nobrega, J.E., Jr.; Rosa, P.; Rovani, M.T.; Ilha, G.F.; Bohrer, R.C.; Ferreira, R.; Gasperin, B.G.; et al. Natriuretic peptide system regulation in granulosa cells during follicle deviation and ovulation in cattle. Reprod. Domest. Anim. 2018, 53, 710–717. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Lagaly, D.V.; Aad, P.Y.; Grado-Ahuir, J.A.; Hulsey, L.B.; Spicer, L.J. Role of adiponectin in regulating ovarian theca and granulosa cell function. Mol. Cell. Endocrinol. 2008, 284, 38–45. [Google Scholar] [CrossRef]
- Luna, L.G.; Armed Forces Institute of Pathology. Manual of Histologic Staining Methods of the Armed Forces Institute of Pathology, 3rd ed.; Blakiston Division: New York, NY, USA, 1968; Volume XII, 258p. [Google Scholar]
- Kowalik, M.K.; Martyniak, M.; Rekawiecki, R.; Kotwica, J. Expression and immunolocalization of membrane progesterone receptors in the bovine oviduct. Domest. Anim. Endocrinol. 2016, 55, 83–96. [Google Scholar] [CrossRef]
- Thatcher, W.W.; Bilby, T.R.; Bartolome, J.A.; Silvestre, F.; Staples, C.R.; Santos, J.E. Strategies for improving fertility in the modern dairy cow. Theriogenology 2006, 65, 30–44. [Google Scholar] [CrossRef]
- Crowe, M.A.; Hostens, M.; Opsomer, G. Reproductive management in dairy cows-the future. Ir. Vet. J. 2018. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Wang, H.; Xu, Y.; Yu, D.; Li, D.; Liu, X.; Du, W. Insulin-like growth factor-1 (IGF-1) promotes myoblast proliferation and skeletal muscle growth of embryonic chickens via the PI3K/Akt signalling pathway. Cell Biol. Int. 2015, 39, 910–922. [Google Scholar] [CrossRef] [PubMed]
- Chao, W.; D’Amore, P.A. IGF2: Epigenetic regulation and role in development and disease. Cytokine Growth Factor Rev. 2008, 19, 111–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lucy, M.C.; Escalante, R.C.; Keisler, D.H.; Lamberson, W.R.; Mathew, D.J. Short communication: Glucose infusion into early postpartum cows defines an upper physiological set point for blood glucose and causes rapid and reversible changes in blood hormones and metabolites. J. Dairy Sci. 2013, 96, 5762–5768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirby, C.J.; Thatcher, W.W.; Collier, R.J.; Simmen, F.A.; Lucy, M.C. Effects of growth hormone and pregnancy on expression of growth hormone receptor, insulin-like growth factor-I, and insulin-like growth factor binding protein-2 and -3 genes in bovine uterus, ovary, and oviduct. Biol. Reprod. 1996, 55, 996–1002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimizu, M.; Deguchi, A.; Hara, Y.; Moriwaki, H.; Weinstein, I.B. EGCG inhibits activation of the insulin-like growth factor-1 receptor in human colon cancer cells. Biochem. Biophys. Res. Commun. 2005, 334, 947–953. [Google Scholar] [CrossRef]
- Hwa, V.; Oh, Y.; Rosenfeld, R.G. The insulin-like growth factor-binding protein (IGFBP) superfamily. Endocr. Rev. 1999, 20, 761–787. [Google Scholar] [CrossRef]
- Robinson, R.S.; Mann, G.E.; Gadd, T.S.; Lamming, G.E.; Wathes, D.C. The expression of the IGF system in the bovine uterus throughout the oestrous cycle and early pregnancy. J. Endocrinol. 2000, 165, 231–243. [Google Scholar] [CrossRef] [Green Version]
- Ha, W.T.; Jeong, H.Y.; Lee, S.Y.; Song, H. Effects of the insulin-like growth factor pathway on the regulation of mammary gland development. Dev. Reprod. 2016, 20, 179–185. [Google Scholar] [CrossRef] [Green Version]
- Firth, S.M.; Baxter, R.C. Cellular actions of the insulin-like growth factor binding proteins. Endocr. Rev. 2002, 23, 824–854. [Google Scholar] [CrossRef]
- Andress, D.L.; Birnbaum, R.S. Human osteoblast-derived insulin-like growth factor (IGF) binding protein-5 stimulates osteoblast mitogenesis and potentiates IGF action. J. Biol. Chem. 1992, 267, 22467–22472. [Google Scholar] [PubMed]
- Badinga, L.; Song, S.; Simmen, R.C.; Clarke, J.B.; Clemmons, D.R.; Simmen, F.A. Complex mediation of uterine endometrial epithelial cell growth by insulin-like growth factor-II (IGF-II) and IGF-binding protein-2. J. Mol. Endocrinol. 1999, 23, 277–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wathes, D.C.; Reynolds, T.S.; Robinson, R.S.; Stevenson, K.R. Role of the insulin-like growth factor system in uterine function and placental development in ruminants. J. Dairy Sci. 1998, 81, 1778–1789. [Google Scholar] [CrossRef]
- Jones, J.I.; Gockerman, A.; Busby, W.H., Jr.; Wright, G.; Clemmons, D.R. Insulin-like growth factor binding protein 1 stimulates cell migration and binds to the alpha 5 beta 1 integrin by means of its Arg-Gly-Asp sequence. Proc. Natl. Acad. Sci. USA 1993, 90, 10553–10557. [Google Scholar] [CrossRef] [Green Version]
- Russo, V.C.; Azar, W.J.; Yau, S.W.; Sabin, M.A.; Werther, G.A. IGFBP-2: The dark horse in metabolism and cancer. Cytokine Growth Factor Rev. 2015, 26, 329–346. [Google Scholar] [CrossRef]
- Bach, L.A. IGF-binding proteins. J. Mol. Endocrinol. 2018, 61, T11–T28. [Google Scholar] [CrossRef] [Green Version]
- Chakravarthy, M.V.; Abraha, T.W.; Schwartz, R.J.; Fiorotto, M.L.; Booth, F.W. Insulin-like growth factor-I extends in vitro replicative life span of skeletal muscle satellite cells by enhancing G1/S cell cycle progression via the activation of phosphatidylinositol 3’-kinase/Akt signaling pathway. J. Biol. Chem. 2000, 275, 35942–35952. [Google Scholar] [CrossRef] [Green Version]
- Tian, B.; Zhao, Y.; Liang, T.; Ye, X.; Li, Z.; Yan, D.; Fu, Q.; Li, Y. Curcumin inhibits urothelial tumor development by suppressing IGF2 and IGF2-mediated PI3K/AKT/mTOR signaling pathway. J. Drug Target. 2017, 25, 626–636. [Google Scholar] [CrossRef]
- Fernandez, J.G.; Rodriguez, D.A.; Valenzuela, M.; Calderon, C.; Urzua, U.; Munroe, D.; Rosas, C.; Lemus, D.; Diaz, N.; Wright, M.C.; et al. Survivin expression promotes VEGF-induced tumor angiogenesis via PI3K/Akt enhanced beta-catenin/Tcf-Lef dependent transcription. Mol. Cancer 2014, 13, 209. [Google Scholar] [CrossRef] [Green Version]
- Coppock, H.A.; White, A.; Aplin, J.D.; Westwood, M. Matrix metalloprotease-3 and -9 proteolyze insulin-like growth factor-binding protein-1. Biol. Reprod. 2004, 71, 438–443. [Google Scholar] [CrossRef] [Green Version]
- Estella, C.; Herrer, I.; Atkinson, S.P.; Quinonero, A.; Martinez, S.; Pellicer, A.; Simon, C. Inhibition of histone deacetylase activity in human endometrial stromal cells promotes extracellular matrix remodelling and limits embryo invasion. PLoS ONE 2012, 7, e30508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manase, K.; Endo, T.; Chida, M.; Nagasawa, K.; Honnma, H.; Yamazaki, K.; Kitajima, Y.; Goto, T.; Kanaya, M.; Hayashi, T.; et al. Coordinated elevation of membrane type 1-matrix metalloproteinase and matrix metalloproteinase-2 expression in rat uterus during postpartum involution. Reprod. Biol. Endocrinol. 2006, 4, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagase, H.; Visse, R.; Murphy, G. Structure and function of matrix metalloproteinases and TIMPs. Cardiovasc. Res. 2006, 69, 562–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Prepartum | Post-Natal |
|---|---|---|
| Ingredient (% of DM) | ||
| Corn silage | 24.7 | 30.2 |
| Oat hay | 55.3 | 12.8 |
| Alfalfa hay | - | 17.1 |
| Ground shelled corn | 5.9 | 7.0 |
| Wheat bran | 9.1 | 21.3 |
| Soybean meal (49% CP) | 4.1 | 8.1 |
| Calcium carbonate | 0.23 | 1.13 |
| Calcium hydrophosphate | 0.23 | 0.45 |
| Salt | 0.16 | 0.45 |
| Sodium bicarbonate | - | 0.68 |
| Magnesium oxide | 0.05 | 0.09 |
| Potassium chloride | - | 0.32 |
| Vitamin and mineral mix 1 | 0.23 | 0.41 |
| Chemical analysis, % of DM | ||
| CP | 11.6 | 14.6 |
| Fat | 2.0 | 2.1 |
| Starch | 10.3 | 14.8 |
| NDF | 53.6 | 45.2 |
| ADF | 31.3 | 25.5 |
| Ash | 6.8 | 6.0 |
| Ca | 0.50 | 0.98 |
| P | 0.41 | 0.54 |
| NEL 2, Mcal/kg | 1.30 | 1.37 |
| Target Genes 1 | Primer Sequence (5’ → 3’) | Product Length, bp | Accession No. 2 |
|---|---|---|---|
| GAPDH | Forward (F): ACCCAGAAGACTGTGGATGG | 178 | NM_001034034.2 |
| Reverse (R): CAACAGACACGTTGGGAGTG | |||
| IGF1 | F: TCCCATCTCCCTGGATTTCT | 105 | NM_001077828 |
| R: GGGTTGGAAGACTGCTGATT | |||
| IGF2 | F: GCTTCTACTTCAGCCGACCAT | 113 | NM_174087 |
| R:GGCACAGTAAGTCTCCAGCAG | |||
| IGF1R | F: AAGGTCCTCAGCGAGTTGTTT | 101 | XM_606794 |
| R: GATCCCGTGTTCTTCTACGTTC | |||
| IGF2R | F: AAGCCTCCCACTATCAACAGAA | 111 | NM_174352 |
| R: TACAACTTCCGGTGGTACACCA | |||
| MMP3 | F: GATGATGAACAATGGACAAAGG | 134 | XM_586521 |
| R: CGAGGGTCGTAGACTGGGTA | |||
| MMP9 | F: GAGGGTAAGGTGCTGCTGTTC | 236 | NM_174744 |
| R: AAGGTCACGTAGCCCACATAGT | |||
| IGFBP1 | F: TCAAGAAGTGGAAGGAGCCCT | 127 | NM_174554 |
| R: AATCCATTCTTGTTGCAGTTT | |||
| IGFBP2 | F: AATCCATTCTTGTTGCAGTTT | 120 | NM_174555 |
| R: AGGGTGGCAAACATCACCT | |||
| IGFBP3 | F: GAAGGCGCATGGTGGAGAT | 102 | NM_174556 |
| R: ACAGACACCCAGAACTTCTCCTC | |||
| IGFBP4 | F: GCCCTGTGGGGTGTACAC | 342 | NM_174557 |
| R: TGCAGCTCACTCTGGCAG | |||
| IGFBP5 | F: TGCGAGCTGGTCAAGGAG | 257 | NM_001105327 |
| R: TCCTCTGCCATCTCGGAG | |||
| IGFBP6 | F: AGAAAGAGGATTTGCCTTTGC | 324 | NM_001040495 |
| R: TCCGGTAGAAGCCCCTATG |
| Antibodies 1 | Type | Suppliers | Dilution |
|---|---|---|---|
| Primary antibodies | |||
| p-AKT | Rabbit Polyclonal 4060 | Cell Signaling Technology, Danvers, MA, USA | 1/2000 |
| AKT | Rabbit Polyclonal 9272 | Cell Signaling Technology, Danvers, MA, USA | 1/1000 |
| p-mTOR | Rabbit Polyclonal 2971 | Cell Signaling Technology, Danvers, MA, USA | 1/1000 |
| mTOR | Rabbit Polyclonal ab2732 | Abcam, Cambridge, UK | 1/1500 |
| p-PI3K | Rabbit Polyclonal bs-5582R | Biosynthesis Biotechnology, Beijing, China | 1/200 |
| PI3K | Rabbit Polyclonal | Biorbyt, Cambridge, UK | 1/300 |
| IGF-1 | Goat Polyclonal orb125044 | Biorbyt, Cambridge, UK | 1/500 |
| IGF-1R | Rabbit Polyclonal ab39675 | Biosynthesis Biotechnology, Beijing, China | 1/500 |
| IGF-2 | Rabbit Polyclonal ab9574 | Abcam, Cambridge, UK | 1/2000 |
| IGF-2R | Rabbit Polyclonal | Proteintech, USA | 1/1000 |
| IGFBP1 | Rabbit Polyclonal ab231254 | Abcam, Cambridge, UK | 2 μg/mL |
| IGFBP4 | Rabbit Polyclonal 483393 | United States Biological, Swampscott, MA, USA | 1/500 |
| GAPDH | Mouse Monoclonal 60004-1-lg | Proteintech, USA | 1/5000 |
| Secondary antibodies | |||
| Rabbit anti-goat IgG | SA00001-4 | Proteintech, USA | 1/3000 |
| Goat anti-rabbit IgG | SA00001-2 | Proteintech, USA | 1/3000 |
| Donkey anti-Mouse | SA00001-8 | Proteintech, USA | 1/3000 |
| Item | CT | RPG | p |
|---|---|---|---|
| Body weight (kg) | 537.5 ± 65.14 | 486.66 ± 20.27 | 0.28 |
| Uterine weight (Kg) | 1.42 ± 0.36 | 1.72 ± 0.21 | 0.18 |
| Uterine length (cm) | 35 ± 5.25 | 35.4 ± 3.21 | 0.91 |
| Uterine width (cm) | 25.3 ± 3.46 | 29.58 ± 2.78 | 0.77 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Han, X.; Tan, Z.; Kang, J.; Wang, Z. Rumen-Protected Glucose Stimulates the Insulin-Like Growth Factor System and mTOR/AKT Pathway in the Endometrium of Early Postpartum Dairy Cows. Animals 2020, 10, 357. https://doi.org/10.3390/ani10020357
Wang Y, Han X, Tan Z, Kang J, Wang Z. Rumen-Protected Glucose Stimulates the Insulin-Like Growth Factor System and mTOR/AKT Pathway in the Endometrium of Early Postpartum Dairy Cows. Animals. 2020; 10(2):357. https://doi.org/10.3390/ani10020357
Chicago/Turabian StyleWang, Yan, Xuefeng Han, Zhiliang Tan, Jinhe Kang, and Zheng Wang. 2020. "Rumen-Protected Glucose Stimulates the Insulin-Like Growth Factor System and mTOR/AKT Pathway in the Endometrium of Early Postpartum Dairy Cows" Animals 10, no. 2: 357. https://doi.org/10.3390/ani10020357