Detection and Characterization of Leptospira Infection and Exposure in Rats on the Caribbean Island of Saint Kitts

 , , ,
, , ,
Abstract
:Simple Summary
Abstract
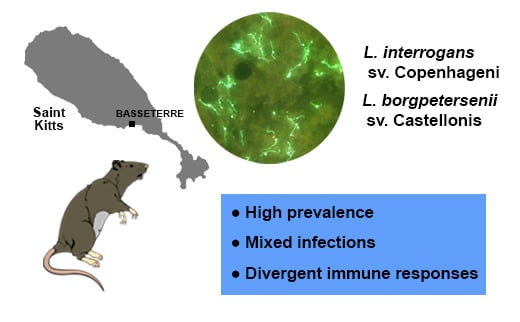
1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Study Location and Animals
2.3. Leptospira Culture
2.4. Leptospira Detection
2.5. Detection of Host Antibody Response
2.6. Characterization of Leptospira Isolates
2.7. Statistical Analysis
3. Results
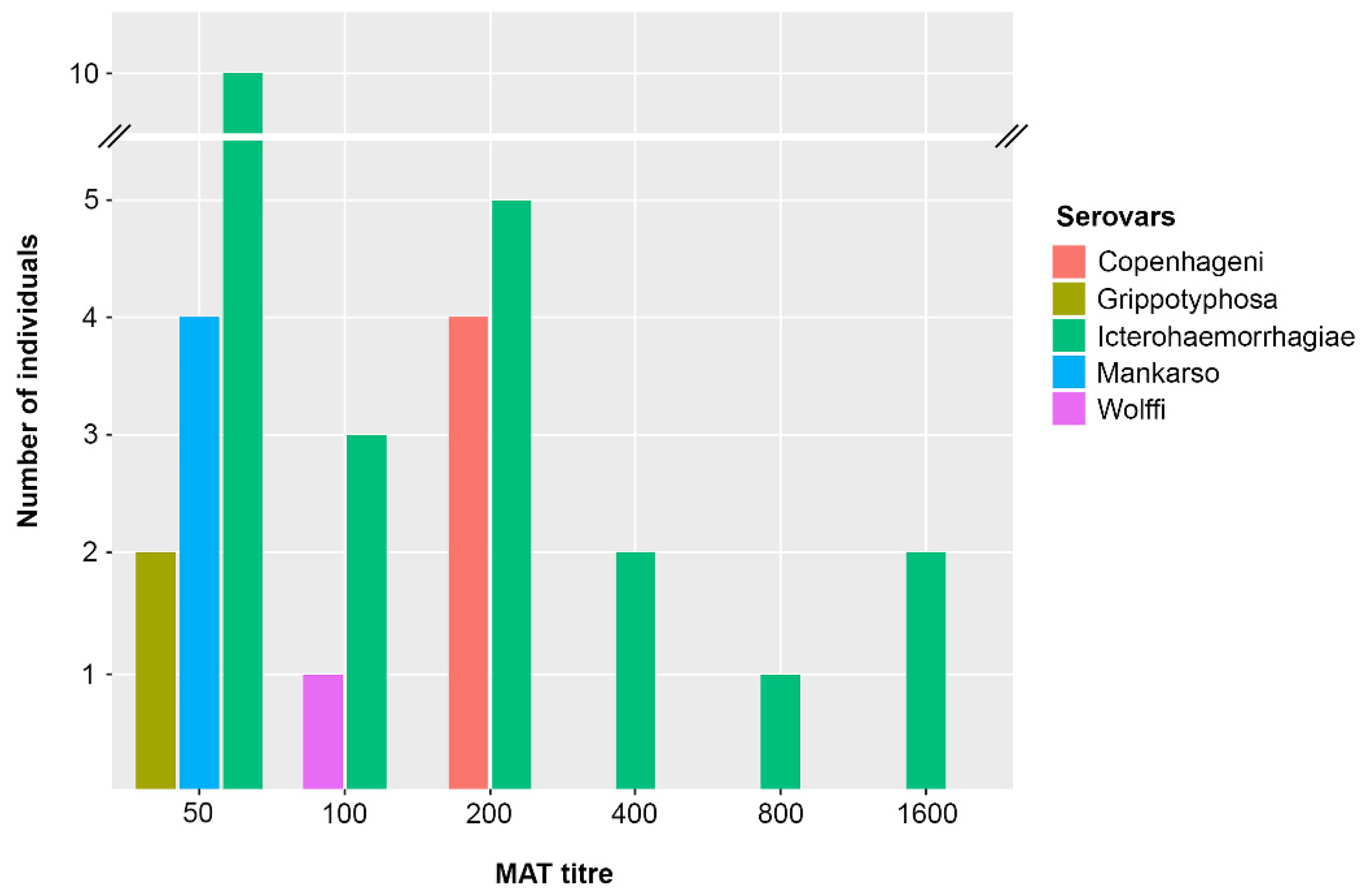
3.1. Detection of Leptospira Exposure and Infection in Rats
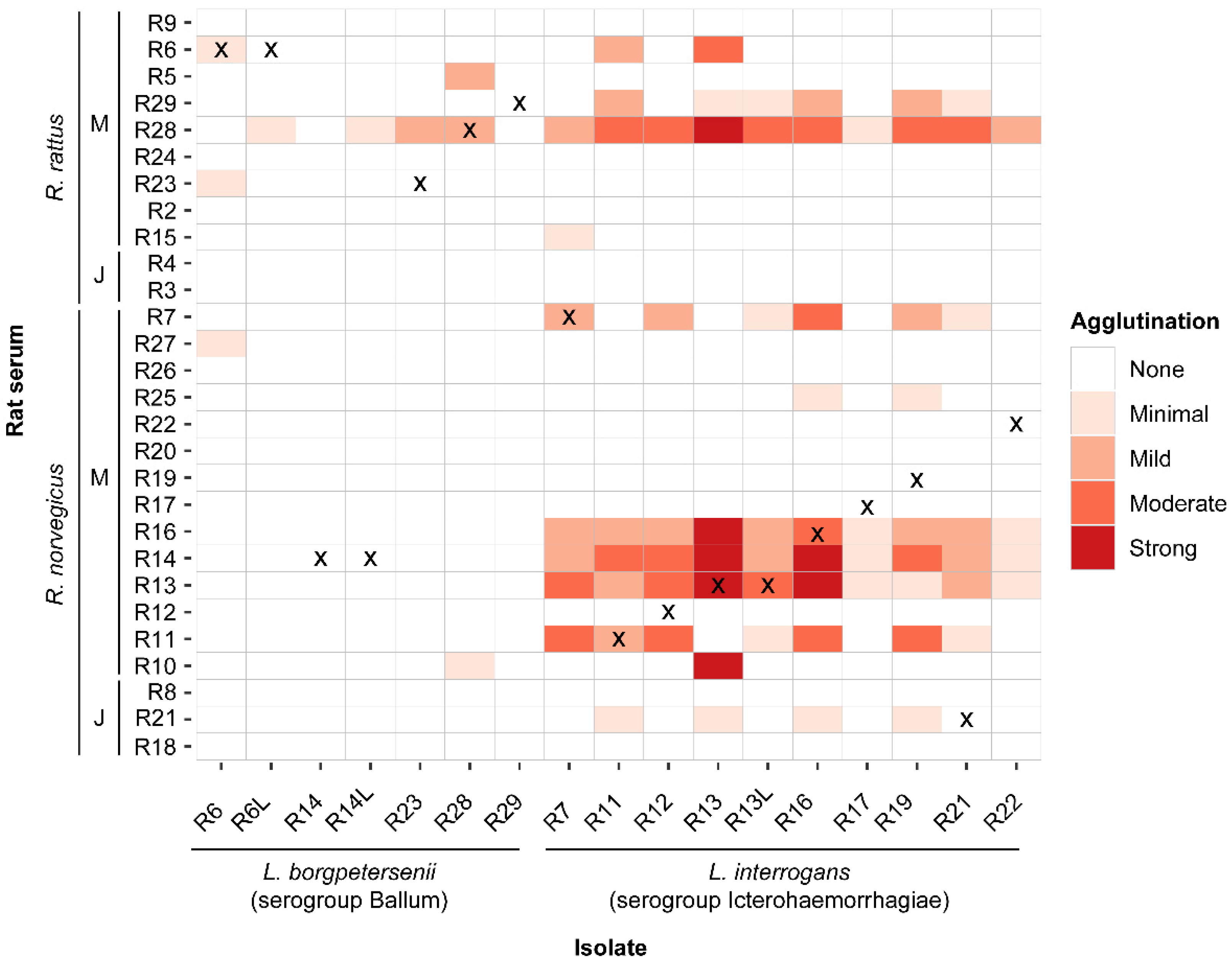
3.2. Characterization of Leptospira Isolates
3.3. Characterization of Immune Response in Rats
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Costa, F.; Hagan, J.E.; Calcagno, J.; Kane, M.; Torgerson, P.; Martinez-Silveira, M.S.; Stein, C.; Abela-Ridder, B.; Ko, A.I. Global Morbidity and Mortality of Leptospirosis: A Systematic Review. PLoS Negl. Trop. Dis. 2015, 9, e0003898. [Google Scholar] [CrossRef] [PubMed]
- Ellis, W.A. Animal leptospirosis. Curr. Top. Microbiol. Immunol. 2015, 387, 99–137. [Google Scholar] [PubMed]
- Haake, D.A.; Levett, P.N. Leptospirosis in humans. Curr. Top. Microbiol. Immunol. 2015, 387, 65–97. [Google Scholar] [PubMed] [Green Version]
- Ido, Y.; Hoki, R.; Ito, H.; Wani, H. The Rat As a Carrier of Sirochaeta Icterohaemorrhagiae, The Causative Agent of Weil’s Disease (Sirochaetosis Icterohaemorrhagica). J. Exp. Med. 1917, 26, 341–353. [Google Scholar] [CrossRef] [Green Version]
- Costa, F.; Wunder, E.A.; de Oliveira, D.; Bisht, V.; Rodrigues, G.; Reis, M.G.; Ko, A.I.; Begon, M.; Childs, J.E. Patterns in Leptospira Shedding in Norway Rats (Rattus norvegicus) from Brazilian Slum Communities at High Risk of Disease Transmission. PLoS Negl. Trop. Dis. 2015, 9, e0003819. [Google Scholar] [CrossRef] [Green Version]
- Boey, K.; Shiokawa, K.; Rajeev, S. Leptospira infection in rats: A literature review of global prevalence and distribution. PLoS Negl. Trop. Dis. 2019, 13, e0007499. [Google Scholar] [CrossRef]
- Shiokawa, K.; Llanes, A.; Hindoyan, A.; Cruz-Martinez, L.; Welcome, S.; Rajeev, S. Peridomestic small Indian mongoose: An invasive species posing as potential zoonotic risk for leptospirosis in the Caribbean. Acta Trop. 2019, 190, 166–170. [Google Scholar] [CrossRef]
- Pratt, N.; Conan, A.; Rajeev, S. Leptospira Seroprevalence in Domestic Dogs and Cats on the Caribbean Island of Saint Kitts. Vet. Med. Int. 2017, 2017, 5904757. [Google Scholar] [CrossRef] [Green Version]
- Shiokawa, K.; Welcome, S.; Kenig, M.; Lim, B.; Rajeev, S. Epidemiology of Leptospira infection in livestock species in Saint Kitts. Trop. Anim. Health Prod. 2019, 51, 1645–1650. [Google Scholar] [CrossRef]
- Rajeev, S.; Conan, A.; Pratt, N.; Beierschmitt, A.; Palmour, R. High Leptospira seroprevalence in captive and wild-caught vervet monkeys (Chlorocebus sabeus) on the Caribbean island of Saint Kitts. J. Vet. Diagn. Invest. 2017, 29, 930–934. [Google Scholar] [CrossRef] [Green Version]
- Larson, C.R.; Dennis, M.; Nair, R.V.; Llanes, A.; Peda, A.; Welcome, S.; Rajeev, S. Isolation and characterization of Leptospira interrogans serovar Copenhageni from a dog from Saint Kitts. JMM Case Rep. 2017, 4, e005120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faine, S.; Adler, B.; Bolin, C.; Perolat, P. Prevention of leptospirosis—Part 1: Identification of high prevalence areas through surveillance and statistics. In Leptospira and Leptospirosis, 2nd ed.; Faine, S., Adler, B., Bolin, C., Perolat, P., Eds.; MedSci: Melbourn, Australia, 1999; pp. 143–150. ISBN 9783662450581. [Google Scholar]
- Yasuda, S.P.; Vogel, P.; Tsuchiya, K.; Han, S.H.; Lin, L.K.; Suzuki, H. Phylogeographic patterning of mtDNA in the widely distributed harvest mouse (Micromys minutus) suggests dramatic cycles of range contraction and expansion during the mid- to late Pleistocene. Can. J. Zool. 2005, 83, 1411–1420. [Google Scholar] [CrossRef] [Green Version]
- Zuerner, R.L. Laboratory Maintenance of Pathogenic Leptospira. Curr. Protoc. Microbiol. 2005. [Google Scholar] [CrossRef]
- Stoddard, R.A. Detection of pathogenic leptospira spp. through real-time PCR (qPCR) targeting the lipL32 gene. Methods Mol. Biol. 2013, 943, 257–266. [Google Scholar]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Tatusova, T.; DiCuccio, M.; Badretdin, A.; Chetvernin, V.; Nawrocki, E.P.; Zaslavsky, L.; Lomsadze, A.; Pruitt, K.D.; Borodovsky, M.; Ostell, J. NCBI prokaryotic genome annotation pipeline. Nucleic Acids Res. 2016, 44, 6614–6624. [Google Scholar] [CrossRef]
- Li, H.; Durbin, R. Fast and accurate long-read alignment with Burrows-Wheeler transform. Bioinformatics 2010, 26, 589–595. [Google Scholar] [CrossRef] [Green Version]
- McKenna, A.; Hanna, M.; Banks, E. The Genome Analysis Toolkit: A MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef] [Green Version]
- Rajeev, S.; Ilha, M.; Woldemeskel, M.; Berghaus, R.D.; Pence, M.E. Detection of asymptomatic renal Leptospira infection in abattoir slaughtered cattle in southeastern Georgia, United States. SAGE Open Med. 2014, 2, 2050312114544696. [Google Scholar] [CrossRef] [Green Version]
- Bulach, D.M.; Zuerner, R.L.; Wilson, P.; Seemann, T.; McGrath, A.; Cullen, P.A.; Davis, J.; Johnson, M.; Kuczek, E.; Alt, D.P.; et al. Genome reduction in Leptospira borgpetersenii reflects limited transmission potential. Proc. Natl. Acad. Sci. USA 2006, 103, 14560–14565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goris, M.G.A.; Hartskeerl, R.A. Leptospirosis serodiagnosis by the microscopic agglutination test. Curr. Protoc. Microbiol. 2014, 1–18. [Google Scholar]
- Levett, P.N. Usefulness of Serologic Analysis as a Predictor of the Infecting Serovar in Patients with Severe Leptospirosis. Clin. Infect. Dis. 2003, 36, 447–452. [Google Scholar] [CrossRef] [PubMed]
- Hathaway, S.C.; Blackmore, D.K. Ecological aspects of the epidemiology of infection with leptospires of the Ballum serogroup in the black rat (Rattus rattus) and the brown rat (Rattus norvegicus) in New Zealand. J. Hyg. 1981, 87, 427–436. [Google Scholar] [CrossRef] [Green Version]
- Minter, A.; Diggle, P.J.; Costa, F.; Childs, J.; Ko, A.I.; Begon, M. Evidence of multiple intraspecific transmission routes for Leptospira acquisition in Norway rats (Rattus norvegicus). Epidemiol. Infect. 2017, 145, 3438–3448. [Google Scholar] [CrossRef] [Green Version]
- De Oliveira, D.; Figueira, C.P.; Zhan, L.; Pertile, A.C.; Pedra, G.G.; Gusmão, I.M.; Wunder, E.A.; Rodrigues, G.; Ramos, E.A.G.; Ko, A.I.; et al. Leptospira in breast tissue and milk of urban Norway rats (Rattus norvegicus). Epidemiol. Infect. 2016, 144, 2420–2429. [Google Scholar] [CrossRef] [Green Version]
- Zuerner, R.L.; Alt, D.P.; Palmer, M.V.; Thacker, T.C.; Olsen, S.C. A Leptospira borgpetersenii serovar Hardjo vaccine induces a Th1 response, activates NK cells, and reduces renal colonization. Clin. Vaccine Immunol. 2011, 18, 684–691. [Google Scholar] [CrossRef] [Green Version]
- Dikken, H.; Kmety, E. Serological typing methods of leptospires. In Methods in Microbiology; Academic Press: London, UK, 1978; Volume 11, pp. 259–307. [Google Scholar]
- Terpstra, W.J.; Korver, H.; Schoone, G.J.; Leeuwen, J.V.; Schönemann, C.E.; De Jonge-Aglibut, S.; Kolk, A.H.J. Comparative classification of Leptospira serovars of the Pomona group by monoclonal antibodies and restriction-endonuclease analysis. Zentralblatt Bakteriol. Mikrobiol. Hyg Abt. 1 Orig. A 1987, 266, 412–421. [Google Scholar] [CrossRef]
- Moseley, M.; Rahelinirina, S.; Rajerison, M.; Garin, B.; Piertney, S.; Telfer, S. Mixed Leptospira infections in a diverse reservoir host community, Madagascar, 2013–2015. Emerg. Infect. Dis. 2018, 24, 1138–1140. [Google Scholar] [CrossRef] [Green Version]
- Rahelinirina, S.; Moseley, M.H.; Allan, K.J.; Ramanohizakandrainy, E.; Ravaoarinoro, S.; Rajerison, M.; Rakotoharinome, V.; Telfer, S. Leptospira in livestock in Madagascar: Uncultured strains, mixed infections and small mammal-livestock transmission highlight challenges in controlling and diagnosing leptospirosis in the developing world. Parasitology 2019, 146, 1707–1713. [Google Scholar] [CrossRef]
- Villanueva, S.Y.A.M.; Ezoe, H.; Baterna, R.A.; Yanagihara, Y.; Muto, M.; Koizumi, N.; Fukui, T.; Okamoto, Y.; Masuzawa, T.; Cavinta, L.L.; et al. Serologic and molecular studies of Leptospira and leptospirosis among rats in the Philippines. Am. J. Trop. Med. Hyg. 2010, 82, 889–898. [Google Scholar] [CrossRef] [PubMed]
- Suepaul, S.M.; Carrington, C.V.; Campbell, M.; Borde, G.; Adesiyun, A.A. Seroepidemiology of leptospirosis in dogs and rats in Trinidad. Trop. Biomed. 2014, 31, 853–861. [Google Scholar] [PubMed]
- Nally, J.E.; Chantranuwat, C.; Wu, X.Y.; Fishbein, M.C.; Pereira, M.M.; Pereira Da Silva, J.J.; Blanco, D.R.; Lovett, M.A. Alveolar Septal Deposition of Immunoglobulin and Complement Parallels Pulmonary Hemorrhage in a Guinea Pig Model of Severe Pulmonary Leptospirosis. Am. J. Pathol. 2004, 164, 1115–1127. [Google Scholar] [CrossRef] [Green Version]
- Cole, J.R.; Ellinghausen, H.C.; Rubin, H.L. Laboratory diagnosis of leptospirosis of domestic animals. Proc. Annu. Meet. U. S. Anim. Health Assoc. 1979, 83, 189–195. [Google Scholar]
- Hathaway, S.C.; Little, T.W.; Pritchard, D.G. Problems associated with the serological diagnosis of Leptospira interrogans serovar Hardjo infection in bovine populations. Vet. Rec. 1986, 119, 84–86. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Species | Number Tested | Number of Positives (Percentage; 95% Confidence Interval) | ||
|---|---|---|---|---|
| RT-PCR | DFA | Culture | ||
| Rattus spp. | 29 | 17 (59; 41–77) | 11 (38; 20–56) | 14 (48; 30–67) |
| R. norvegicus | 18 | 12 (67; 45–89) | 9 (50; 27–73) | 10 (56; 33–79) |
| R. rattus | 11 | 5 (46; 16–75) | 2 (18; 0–41) | 4 (36; 8–65) |
| Isolate Name | WGS ID | Serogrouping Results |
|---|---|---|
| R6 (RR) | L. borgpetersenii | Ballum (Alexi) 1 |
| R6L(RR) | L. borgpetersenii | Ballum (Alexi) |
| R7 (RN) | L. interrogans | Ictero/Copenhageni/Mankarso (Canicola) |
| R11 (RN) | L. interrogans | Ictero/Copenhageni/Mankarso (Canicola) |
| R12 (RN) | L. interrogans | Ictero/Copenhageni/Mankarso (Canicola) |
| R13 (RN) | L. interrogans | Ictero/Copenhageni/Mankarso |
| R13L (RN) | L. interrogans | Ictero/Copenhageni/Mankarso |
| R14 (RN) | L. borgpetersenii | Ballum |
| R14L (RN) | L. borgpetersenii | Ballum/Ictero |
| R16 (RN) | L. interrogans | Ictero/Copenhageni/Mankarso |
| R17 (RN) | L. interrogans | Ictero/Copenhageni/Mankarso |
| R19 (RN) | L. interrogans | Ictero/Copenhageni/Mankarso |
| R21 (RN) | L. interrogans | Ictero/Copenhageni/Mankarso |
| R22 (RN) | L. interrogans | Ictero/Copenhageni/Mankarso |
| R23 (RR) | L. borgpetersenii | Ballum |
| R28 (RR) | L. borgpetersenii | Ballum |
| R29 (RR) | L. borgpetersenii | Ballum |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rajeev, S.; Shiokawa, K.; Llanes, A.; Rajeev, M.; Restrepo, C.M.; Chin, R.; Cedeño, E.; Ellis, E. Detection and Characterization of Leptospira Infection and Exposure in Rats on the Caribbean Island of Saint Kitts. Animals 2020, 10, 350. https://doi.org/10.3390/ani10020350
Rajeev S, Shiokawa K, Llanes A, Rajeev M, Restrepo CM, Chin R, Cedeño E, Ellis E. Detection and Characterization of Leptospira Infection and Exposure in Rats on the Caribbean Island of Saint Kitts. Animals. 2020; 10(2):350. https://doi.org/10.3390/ani10020350
Chicago/Turabian StyleRajeev, Sreekumari, Kanae Shiokawa, Alejandro Llanes, Malavika Rajeev, Carlos Mario Restrepo, Raymond Chin, Eymi Cedeño, and Esteban Ellis. 2020. "Detection and Characterization of Leptospira Infection and Exposure in Rats on the Caribbean Island of Saint Kitts" Animals 10, no. 2: 350. https://doi.org/10.3390/ani10020350