Diversity and Composition of Rumen Bacteria, Fungi, and Protozoa in Goats and Sheep Living in the Same High-Altitude Pasture

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Handling, Diet, and Sampling
2.2. DNA Extraction, PCR Amplification, and Sequencing
2.3. Amplicon Gene Sequence Processing
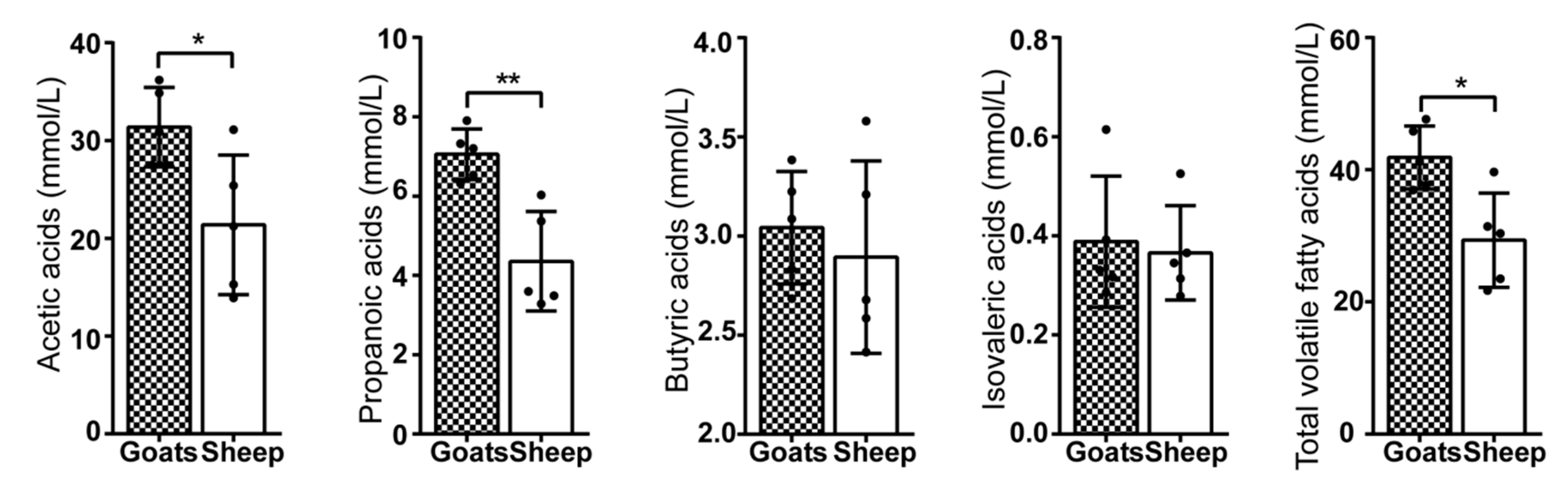
2.4. Volatile Fatty Acids
2.5. Data Analysis
3. Results
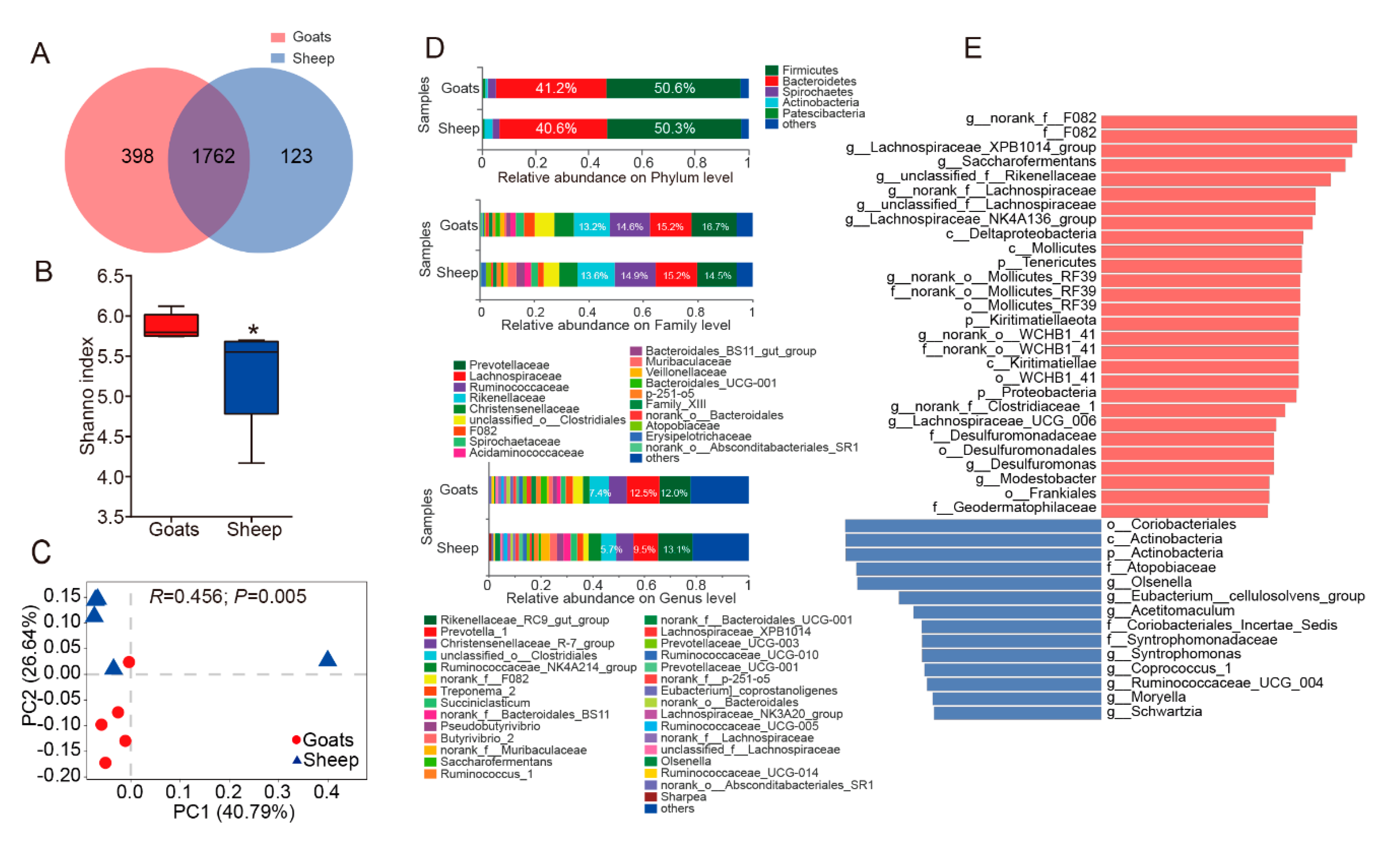
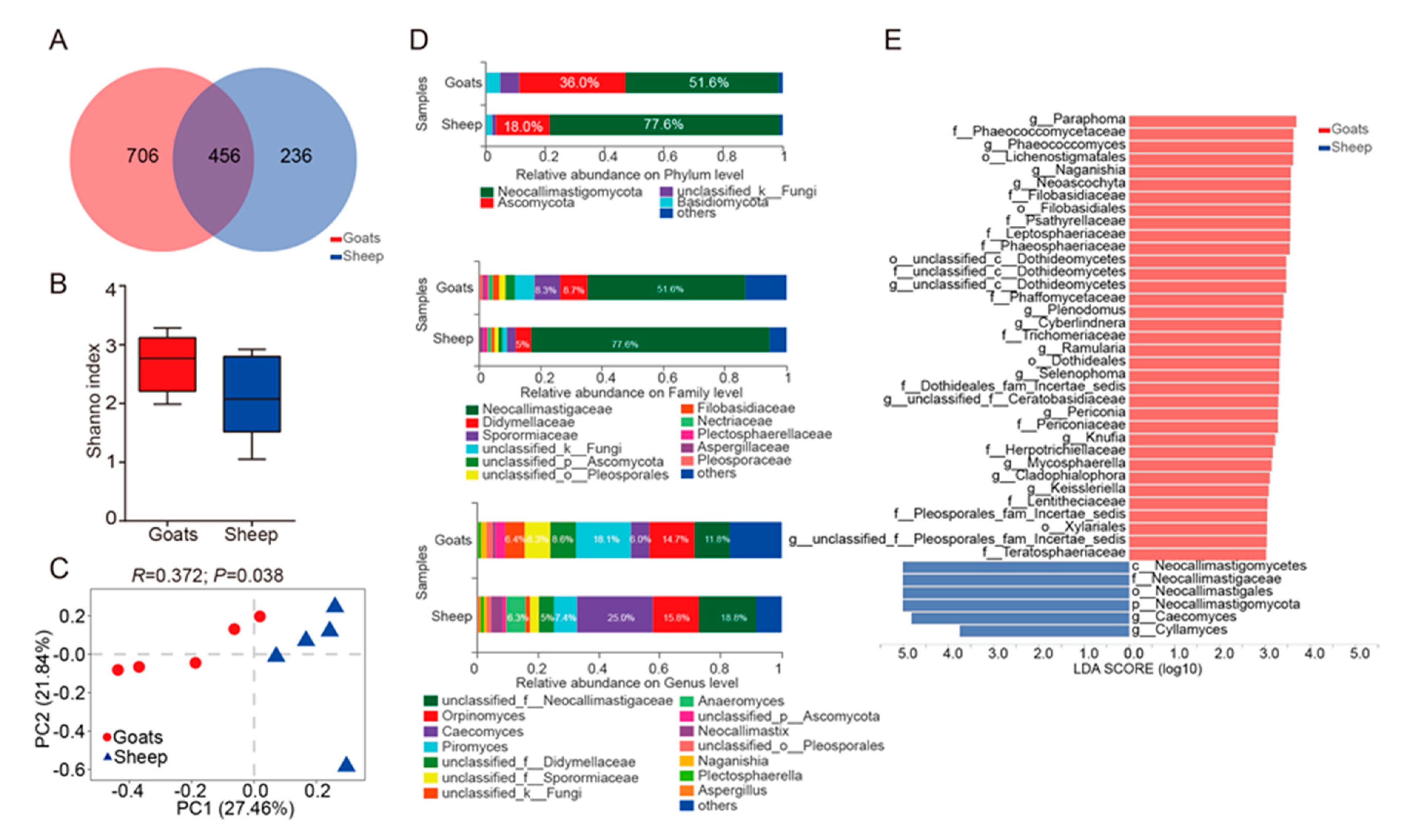
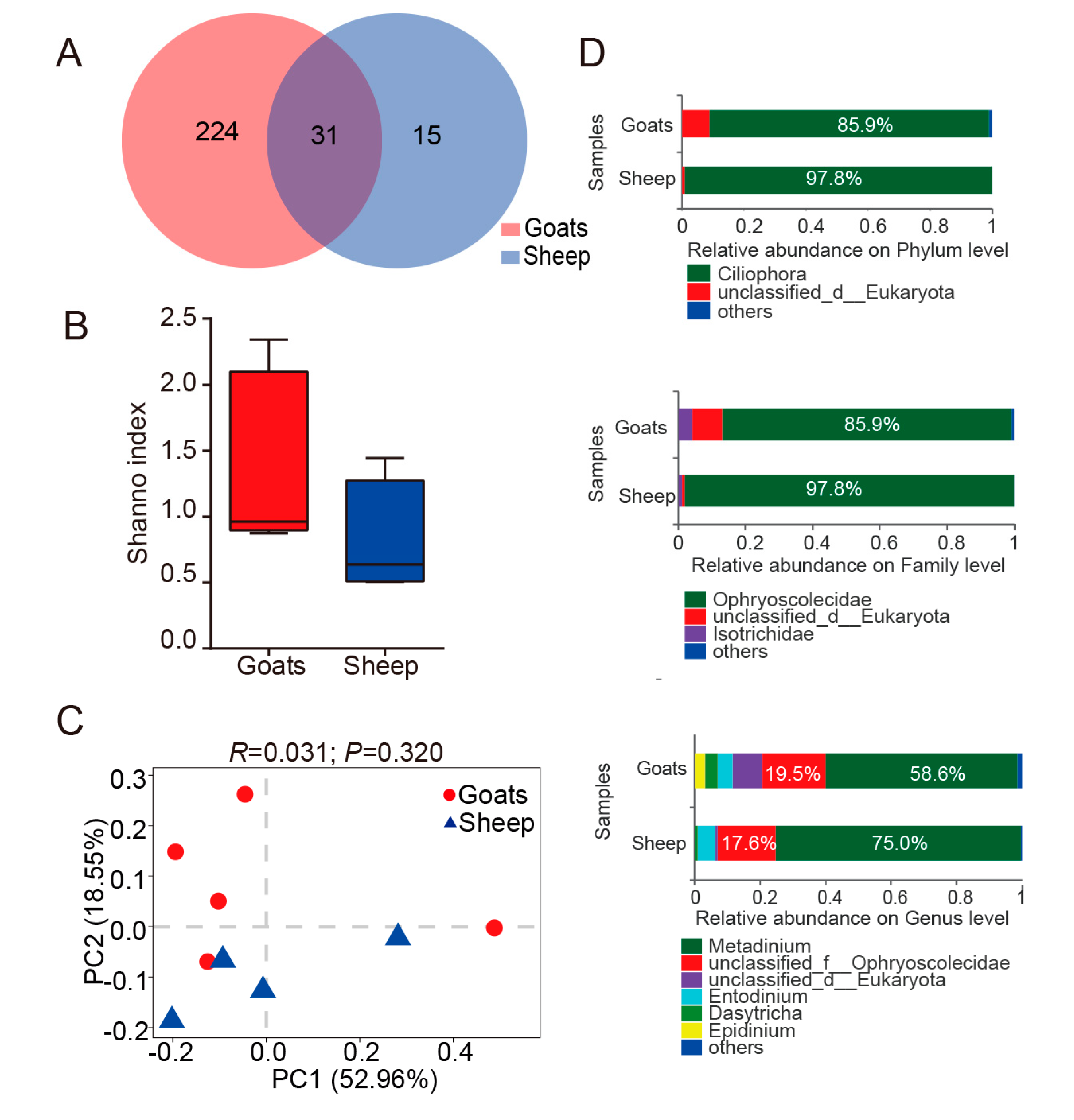
3.1. An Overview of Ruminal Bacteria, Fungi, and Protozoa Composition
3.2. Different Composition of Ruminal Bacteria, Fungi, and Protozoa in Goats and Sheep
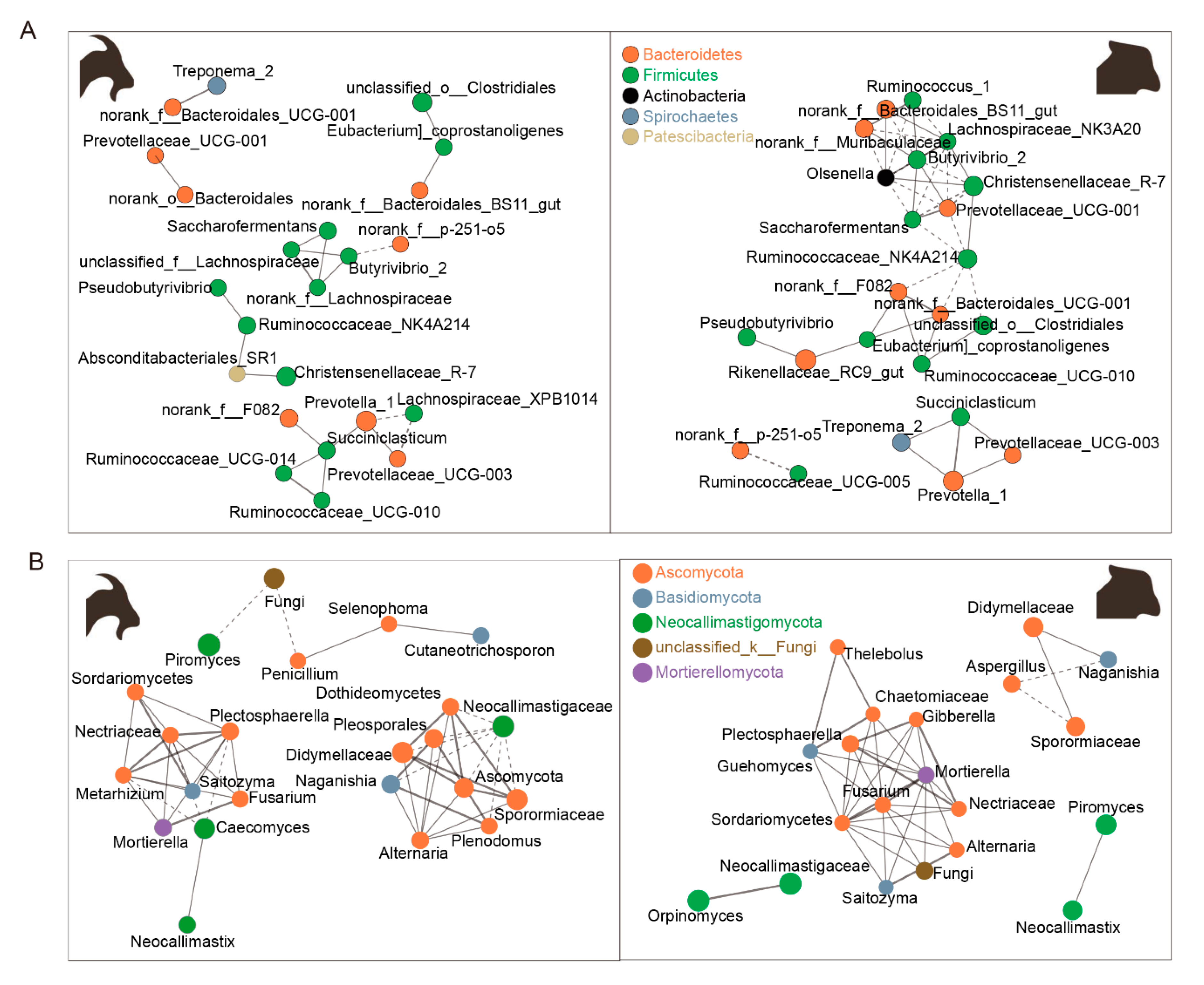
3.3. Rumen Bacteria and Fungi Network between Goats and Sheep
4. Discussion
4.1. Feature of Rumen Microbiota in the High-Altitude Pasture
4.2. Rumen Microbiota May Reflect Host’s Adaptive Capacity of Environment
4.3. The Role of the Diet in Rumen Microbiota of Goats and Sheep
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Simmons, P.; Ekarius, C. Storey’s Guide to Raising Sheep; Storey Publishing: North Adams, MA, USA, 2010; pp. 1–10. ISBN 978-160-342-459-2. [Google Scholar]
- Zeder, M.A.; Hesse, B. The initial domestication of goats (Capra hircus) in the Zagros mountains 10,000 years ago. Science 2000, 287, 2254–2257. [Google Scholar] [CrossRef] [PubMed]
- Jing, X.P.; Zhou, J.W.; Wang, W.J.; Degen, A.A.; Guo, Y.M.; Kang, J.P.; Xu, W.X.; Liu, P.P.; Yang, C.; Shi, F.Y.; et al. Tibetan sheep are better able to cope with low energy intake than Small-tailed Han sheep due to lower maintenance energy requirements and higher nutrient digestibilities. Anim. Feed Sci. Technol. 2019, 254, 114200. [Google Scholar] [CrossRef] [Green Version]
- Malechek, J.C.; Provenza, F. Feeding behaviour and nutrition of goats on rangelands. World Anim. Rev. 1983, 47, 38–48. [Google Scholar]
- Zhang, P.F. Comparative Study on Small Intestinal Mucosal Structure between Tibetan Sheep and Tibetan Goat. J. Anhui Agric. Sci. 2012, 42, 567–571. [Google Scholar] [CrossRef]
- Tao, T.; Kai-Dong, Z.; Yu-Jiang, W.U.; Fang, Y.; Yu-Lin, C.; Yu-Xin, Y. Seasonal Changes of Grazing Grassland Nutrition and Yields and Feeding Preferences of Tibetan Cashmere Goats in Nagqu Prefecture of Tibetan Plateau. Acta Agrestia Sin. 2019, 27, 105–111. [Google Scholar] [CrossRef]
- Narjisse, H.; Elhonsali, M.A.; Olsen, J.D. Effects of oak (Quercus ilex) tannins on digestion and nitrogen balance in sheep and goats. Small Rumin. Res. 1995, 18, 201–206. [Google Scholar] [CrossRef]
- Hofmann, R.R. Evolutionary steps of ecophysiological adaptation and diversification of ruminants: A comparative view of their digestive system. Oecologia 1989, 78, 443–457. [Google Scholar] [CrossRef]
- Kronberg, S.L. Comparative Foraging Ecology of Sheep and Goats in Caatinga Woodland in Northeastern Brazil. Diss. Abstr. Int. B Sci. Eng. 1990, 51, 3200B. [Google Scholar]
- Zhang, Z.; Xu, D.; Li, W.; Hao, J.; Wang, J.; Xin, Z.; Wang, W.; Qiang, Q.; Huang, X.; Zhou, J. Convergent Evolution of Rumen Microbiomes in High-Altitude Mammals. Curr. Biol. 2016, 26, 1873–1879. [Google Scholar] [CrossRef] [Green Version]
- Narjisse, H.; Hansali, M.E. Effect of tannins on nitrogen balance and microbial activity of rumen fluid in sheep and goats. Ann. Zootech. 1985, 34, 482. [Google Scholar] [CrossRef] [Green Version]
- Seth, O.N.; Rai, G.S.; Yadav, P.C.; Pandey, M.D. A note on the rate of secretion and chemical composition of parotid saliva in sheep and goats. Indian J. Anim. Sci. 1977, 46, 660–663. [Google Scholar]
- Silanikove, N.; Tagari, H.; Shkolnik, A. Gross energy digestion and urea recycling in the desert black Bedouin goat. Comp. Biochem. Physiol. Part A Physiol. 1980, 67, 215–218. [Google Scholar] [CrossRef]
- Odenyo, A.A.; Osuji, P.O. Tannin-tolerant ruminal bacteria from East African ruminants. Can. J. Microbiol. 1998, 44, 905–909. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Morales, E.; Arco-Pérez, A.; Martín-García, A.I.; Yáñez-Ruiz, D.R.; Frutos, P.; Hervás, G. Use of stomach tubing as an alternative to rumen cannulation to study ruminal fermentation and microbiota in sheep and goats. Anim. Feed Sci. Technol. 2014, 198, 57–661. [Google Scholar] [CrossRef] [Green Version]
- Frank, K.L.; Rogers, D.R.; Olins, H.C.; Vidoudez, C.; Girguis, P.R. Characterizing the distribution and rates of microbial sulfate reduction at Middle Valley hydrothermal vents. ISME J. 2013, 7, 1391–1401. [Google Scholar] [CrossRef]
- Adams, R.I.; Miletto, M.; Taylor, J.W.; Bruns, T.D. Dispersal in microbes: Fungi in indoor air are dominated by outdoor air and show dispersal limitation at short distances. ISME J. 2013, 7, 1262–1273. [Google Scholar] [CrossRef] [Green Version]
- Lin, L.; Xie, F.; Sun, D.; Liu, J.; Zhu, W.-Y.; Seew, M. Ruminal microbiome-host crosstalk stimulates the development of the ruminal epithelium in a lamb model. Microbiome 2019, 7, 83. [Google Scholar] [CrossRef]
- Martin, M. CUTADAPT removes adapter sequences from high-throughput sequencing reads. EMBnet J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996. [Google Scholar] [CrossRef]
- Yang, S.; Liebner, S.; Alawi, M.; Ebenhöh, O.; Wagner, D. Taxonomic database and cut-off value for processing mcrA gene 454 pyrosequencing data by MOTHUR. J. Microbiol. Methods 2014, 103, 3–5. [Google Scholar] [CrossRef]
- Christian, Q.; Elmar, P.; Pelin, Y.; Jan, G.; Timmy, S.; Pablo, Y.; Jörg, P.; Oliver, G.F. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef]
- Kõljalg, U.; Nilsson, R.H.; Abarenkov, K.; Tedersoo, L.; Taylor, A.; Bahram, M.; Bates, S.; Bruns, T.; Bengtsson-Palme, J.; Callaghan, T.; et al. Towards a unified paradigm for sequence-based identification of Fungi. Mol. Ecol. 2013, 22, 5271–5277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, F.; Yang, X.J.; Cao, Y.C.; Li, S.X.; Yao, J.H.; Li, Z.J.; Sun, F.F. Effects of dietary effective fiber to rumen degradable starch ratios on the risk of sub-acute ruminal acidosis and rumen content fatty acids composition in dairy goat. Anim. Feed Sci. Technol. 2014, 189, 54–62. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, X.T.; Yan, B.Y.; Ren, Q.M.; Dou, J.J.; Qiu, Q. Effect of slow-release urea on the composition of ruminal bacteria and fungi communities in yak. Anim. Feed Sci. Technol. 2018, 244, 18–27. [Google Scholar] [CrossRef]
- Liu, C.; Wu, H.; Liu, S.; Chai, S.; Meng, Q.; Zhou, Z. Dynamic Alterations in Yak Rumen Bacteria Community and Metabolome Characteristics in Response to Feed. Front. Microbiol. 2019, 10, 1116. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.C.; Chen, Y.C.; Hseu, R.S. Purification and characterization of a cellulolytic multienzyme complex produced by Neocallimastix patriciarum J11. Biochem. Biophys. Res. Commun. 2014, 451, 190–195. [Google Scholar] [CrossRef]
- Imai, S.; Abdullah, N.; Ho, Y.W.; Jalaludin, S.; Hussain, H.Y.; Onodera, R.; Kudo, H. Comparative study on the rumen ciliate populations in small experimental herds of water buffalo and Kedah Kelantan cattle in Malaysia. Anim. Feed Sci. Technol. 1995, 52, 345–351. [Google Scholar] [CrossRef]
- Ugbogu, E.A.; Elghandour, M.M.M.Y.; Ikpeazu, V.O.; Buendía, G.R.; Molina, O.M.; Arunsi, U.O.; Emmanuel, O.; Salem, A.Z.M. The potential impacts of dietary plant natural products on the sustainable mitigation of methane emission from livestock farming. J. Clean. Prod. 2019, 213, 915–925. [Google Scholar] [CrossRef]
- Olijhoek, D.W.; Hellwing, A.L.F.; Brask, M.; Weisbjerg, M.R.; Højberg, O.; Larsen, M.K.; Dijkstra, J.; Erlandsen, E.J.; Lund, P. Effect of dietary nitrate level on enteric methane production, hydrogen emission, rumen fermentation, and nutrient digestibility in dairy cows. J. Dairy Sci. 2016, 99, 6191–6205. [Google Scholar] [CrossRef] [Green Version]
- Wei, Y.Q.; Long, R.-J.; Yang, H.; Yang, H.-J.; Shen, X.-H.; Shi, R.-F.; Wang, Z.-Y.; Du, J.-G.; Qi, X.-J.; Ye, Q.-H. Fiber degradation potential of natural co-cultures of Neocallimastix frontalis and Methanobrevibacter ruminantium isolated from yaks (Bos grunniens) grazing on the Qinghai Tibetan Plateau. Anaerobe 2016, 39, 158–164. [Google Scholar] [CrossRef] [PubMed]
- Flint, H.J.; Martin, J.C.; Thomson, A.M. Prevotella bryantii, P. Ruminicola and Bacteroides Strains; Springer: Berlin/Heideberg, Germany, 2000; pp. 140–149. [Google Scholar] [CrossRef]
- Graber, J.R.; Breznak, J.A. Physiology and nutrition of Treponema primitia, an H2CO2-acetogenic spirochete from termite hindguts. Appl. Environ. Microbiol. 2004, 70, 1307–1314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xin, J.; Chai, Z.; Zhang, C.; Zhang, Q.; Zhu, Y.; Cao, H.; Zhong, J.; Ji, Q. Comparing the Microbial Community in Four Stomach of Dairy Cattle, Yellow Cattle and Three Yak Herds in Qinghai-Tibetan Plateau. Front. Microbiol. 2019, 10, 1547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, D.; Chen, H.; Zhao, X.; Xu, S.; Hu, L.; Xu, T.; Jiang, L.; Zhan, W. Rumen prokaryotic communities of ruminants under different feeding paradigms on the Qinghai-Tibetan Plateau. Syst. Appl. Microbiol. 2017, 40, 227–236. [Google Scholar] [CrossRef]
- Wei, G.; Sisi, B.; Jingpeng, K.; Ying, Z.; Ruijun, L.; Xiaodan, H.; Shan, M.N.; Anderson, R.C. Bacterial communities related to 3-nitro-1-propionic acid degradation in the rumen of grazing ruminants in the Qinghai-Tibetan Plateau. Anaerobe 2018, 54, 42–54. [Google Scholar] [CrossRef]
- Hernández, C.; Alamilla-Ortiz, Z.L.; Escalante, A.E.; Navarro-Díaz, M.; Carrillo-Reyes, J.; Moreno-Andrade, I.; Valdez-Vazquez, I. Heat-shock treatment applied to inocula for H2 production decreases microbial diversities, interspecific interactions and performance using cellulose as substrate. Int. J. Hydrog. Energy 2019, 44, 13126–13134. [Google Scholar] [CrossRef]
- Collins, M.D.; Lawson, P.A.; Willems, A.; Cordoba, J.J.; Fernandezgarayzabal, J.; Garcia, P.; Cai, J.; Hippe, H.; Farrow, J.A.E. The Phylogeny of the Genus Clostridium: Proposal of Five New Genera and Eleven New Species Combinations. Int. J. Syst. Bacteriol. 1994, 44, 812–826. [Google Scholar] [CrossRef] [Green Version]
- Yangchun, T.; Sibing, W.; Diaoqi, Y.; Zengshu, J.; Tuyan, Q. Rumen fermentation and bacterial communities in weaned Chahaer lambs on diet swith different protein levels. J. Integr. Agric. 2016, 15, 1564–1574. [Google Scholar] [CrossRef]
- Kabir, F.; Sultana, M.S.; Shahjalal, M.; Khan, M.J.; Alam, M.Z. Effect of Protein Supplementation on Growth Performance in Female Goats and Sheep under Grazing Condition. Pak. J. Nutr. 2004, 223, 134–140. [Google Scholar] [CrossRef]
- Roesch, L.F.; Lorca, G.L.; Casella, G.; Giongo, A.; Naranjo, A.; Pionzio, A.M.; Li, N.; Mai, V.; Wasserfall, C.H.; Schatz, D. Culture-independent identification of gut bacteria correlated with the onset of diabetes in a rat model. ISME J. 2009, 3, 536–548. [Google Scholar] [CrossRef] [Green Version]
- Finlay, B.J.; Esteban, G.; Clarke, K.J.; Williams, A.G.; Embley, T.M.; Hirt, R.P. Some rumen ciliates have endosymbiotic methanogens. FEMS Microbiol. Lett. 1994, 117, 157–161. [Google Scholar] [CrossRef] [PubMed]
- Naeem, A.; Drackley, J.K.; Stamey, J.; Loor, J.J. Role of metabolic and cellular proliferation genes in ruminal development in response to enhanced plane of nutrition in neonatal Holstein calves. J. Dairy Sci. 2012, 95, 1807–1820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Den Besten, G.; Van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.J.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, X.F.; Zhou, X.L.; Lian, G.Q.; Blachier Francois Liu, G.; Tan, B.E.; Nyachoti, C.M.; Yin, Y.L. Dietary supplementation with chitooligosaccharides alters gut microbiota and modifies intestinal luminal metabolites in weaned Huanjiang mini-piglets. Livest. Sci. 2014, 160, 97–101. [Google Scholar] [CrossRef]
- Frese, S.A.; Parker, K.; Calvert, C.C.; Mills, D.A. Diet shapes the gut microbiome of pigs during nursing and weaning. Microbiome 2015, 3, 28. [Google Scholar] [CrossRef] [Green Version]
- Amir, Z.; Amandine, C.; Shibu, Y.; Satchidananda, P. Diet and feeding pattern affect the diurnal dynamics of the gut microbiome. Cell Metab. 2014, 20, 1006–1017. [Google Scholar] [CrossRef] [Green Version]
- Mckenney, E.A.; O’Connell, T.M.; Rodrigo, A.; Yoder, A.D. Feeding strategy shapes gut metagenomic enrichment and functional specialization in captive lemurs. Gut Microbes 2018, 9, 202–217. [Google Scholar] [CrossRef]
- Ma, Y.; Ma, S.; Chang, L.; Wang, H.; Ga, Q.; Ma, L.; Bai, Z.; Shen, Y.; Ge, R.-L. Gut microbiota adaptation to high altitude in indigenous animals. Biochem. Biophys. Res. Commun. 2019, 516, 120–126. [Google Scholar] [CrossRef]
- Perea, K.; Perz, K.; Olivo, S.K.; Williams, A.; Lachman, M.; Ishaq, S.L.; Thomson, J.; Yeoman, C.J. Feed efficiency phenotypes in lambs involve changes in ruminal, colonic, and small-intestine-located microbiota. J. Anim. Sci 2017, 95, 2585–2592. [Google Scholar] [CrossRef] [Green Version]
- Van Gylswyk, N.O. Succiniclasticum ruminis gen. nov., sp. nov., a ruminal bacterium converting succinate to propionate as the sole energy-yielding mechanism. Int. J. Syst. Bacteriol 1995, 45, 297–300. [Google Scholar] [CrossRef] [Green Version]
- Hooda, S.; Boler, B.M.V.; Serao, M.C.R.; Brulc, J.M.; Staeger, M.A.; Boileau, T.W.; Dowd, S.E.; Fahey, G.C., Jr.; Swanson, K.S. 454 pyrosequencing reveals a shift in fecal microbiota of healthy adult men consuming polydextrose or soluble corn fiber. J. Nutr. 2012, 142, 1259–1265. [Google Scholar] [CrossRef] [PubMed]
- Hespell, R.B. The Genera Succinivibrio and Succinimonas; Springer-Verlag: Berlin, Germany, 1992. [Google Scholar] [CrossRef]
- Grilli, D.J.; Fliegerová, K.; Kopečný, J.; Lama, S.P.; Mrázek, J. Analysis of the rumen bacterial diversity of goats during shift from forage to concentrate diet. Anaerobe 2016, 42, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Kelly, T.N.; Bazzano, L.A.; Ajami, N.J.; He, H.; Zhao, J.; Petrosino, J.F.; Correa, A.; He, J. Gut microbiome associates with lifetime cardiovascular disease risk profile among Bogalusa heart study participants. Circ. Res. 2016, 119, 956–964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hausenloy, D.J.; Yellon, D.M. Myocardial ischemia-reperfusion injury: A neglected therapeutic target. J. Clin. Investig. 2013, 123, 92–100. [Google Scholar] [CrossRef]
- Feldman, B.M. Feeding strategy of the goat. In Caprinos; Cornell International Agriculture, Mimeograph-Cornell University (EUA), Department of Animal Science: Ithaca, NY, USA, 1981. [Google Scholar]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Langda, S.; Zhang, C.; Zhang, K.; Gui, B.; Ji, D.; Deji, C.; Cuoji, A.; Wang, X.; Wu, Y. Diversity and Composition of Rumen Bacteria, Fungi, and Protozoa in Goats and Sheep Living in the Same High-Altitude Pasture. Animals 2020, 10, 186. https://doi.org/10.3390/ani10020186
Langda S, Zhang C, Zhang K, Gui B, Ji D, Deji C, Cuoji A, Wang X, Wu Y. Diversity and Composition of Rumen Bacteria, Fungi, and Protozoa in Goats and Sheep Living in the Same High-Altitude Pasture. Animals. 2020; 10(2):186. https://doi.org/10.3390/ani10020186
Chicago/Turabian StyleLangda, Suo, Chenguang Zhang, Ke Zhang, Ba Gui, De Ji, Ciren Deji, Awang Cuoji, Xiaolong Wang, and Yujiang Wu. 2020. "Diversity and Composition of Rumen Bacteria, Fungi, and Protozoa in Goats and Sheep Living in the Same High-Altitude Pasture" Animals 10, no. 2: 186. https://doi.org/10.3390/ani10020186