Fermented Feed Supplement Relieves Caecal Microbiota Dysbiosis and Kidney Injury Caused by High-Protein Diet in the Development of Gosling Gout


Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and methods
2.1. Ethical Approvals
2.2. Experiment One
2.3. Experiment Two
2.4. Serum Metabolite Measurement
2.5. Histomorphological Observation
2.6. 16S rRNA Sequencing of the Cecum Contents
2.7. Data Analysis
3. Results
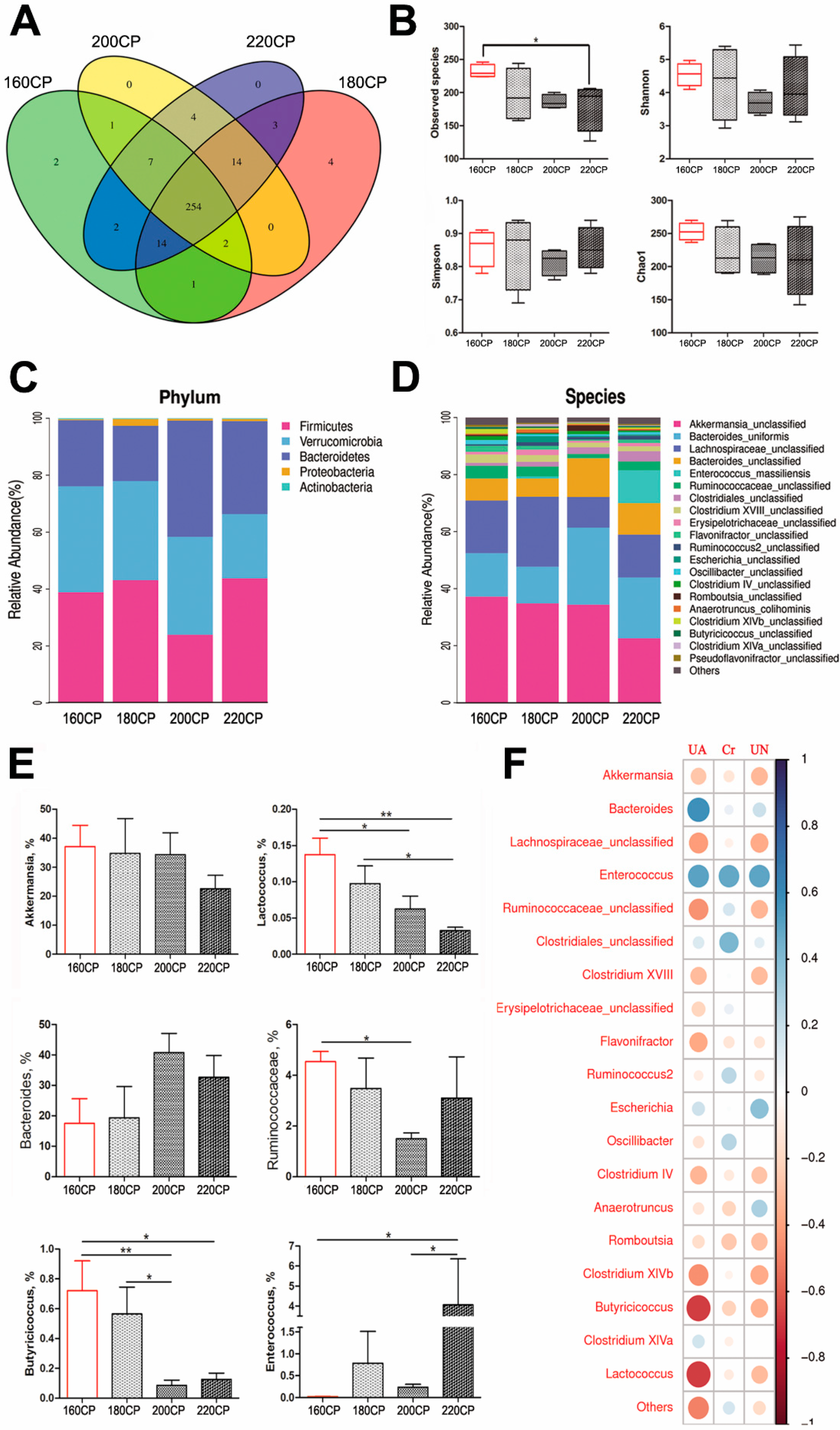
3.1. Kidney Injury and Gut Dysbiosis in High-Protein Diet-Induced Gosling Gout
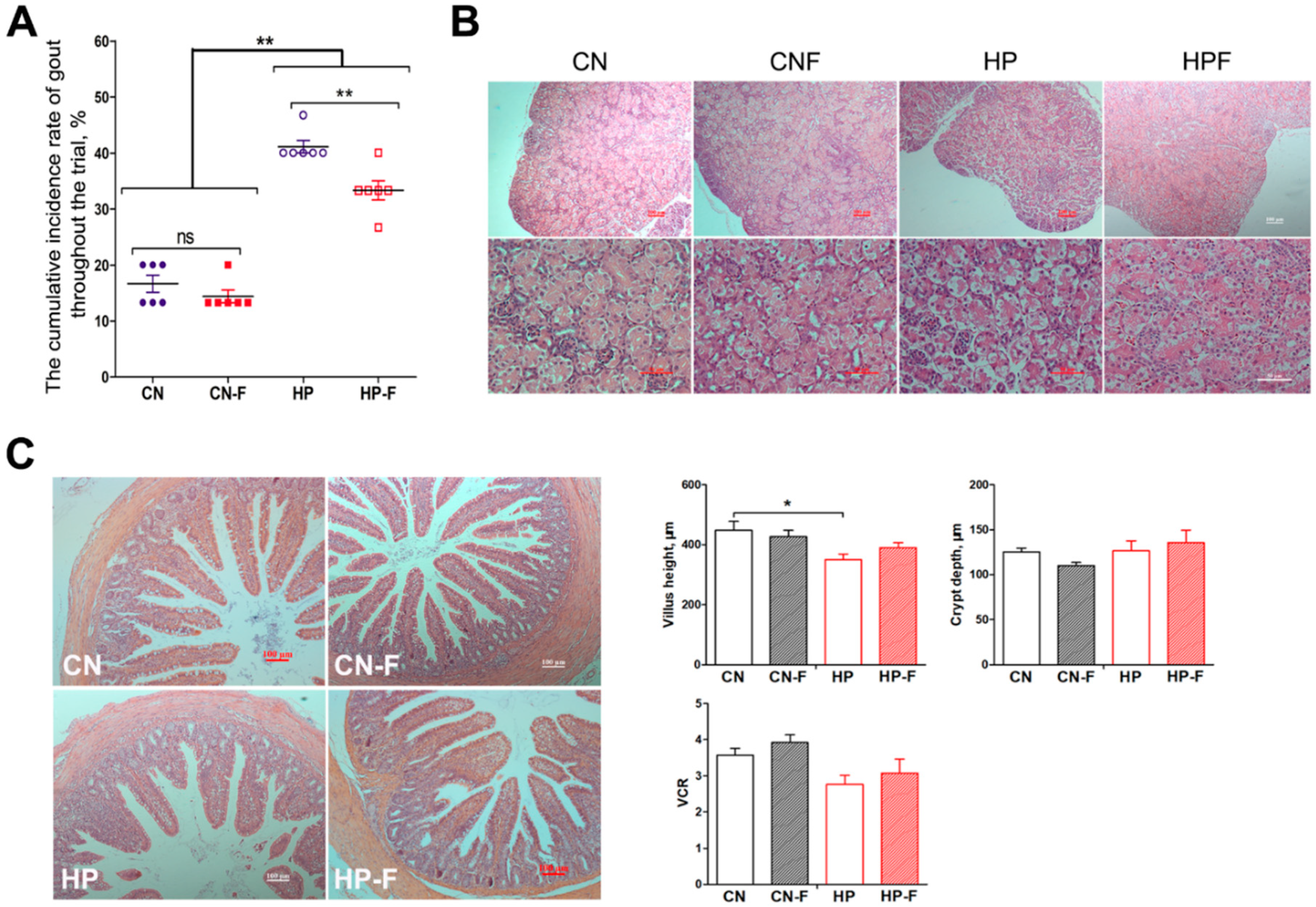
3.2. Effects of Fermented Feed on Gut Dysbiosis and Kidney Injury Caused by High-Protein Diets
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Xi, Y.; Ying, S.; Shao, C.; Zhu, H.; Yan, J.; Shi, Z. Metabolomic profiling of goslings with visceral gout reveals a distinct metabolic signature. Br. Poult. Sci. 2020, 61, 258–265. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Cao, Y.; Wang, J.; Fu, G.; Sun, M.; Zhang, L.; Meng, L.; Cui, G.; Huang, Y.; Hu, X.; et al. Isolation and characterization of an astrovirus causing fatal visceral gout in domestic goslings. Emerg. Microbes Infect. 2018, 7, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaba, A.; Dave, H.; Pal, J.K.; Prajapati, K.S. Isolation, identification and molecular characterization of IBV variant from out break of visceral gout in commercial broilers. Vet. World 2010, 65, 431–438. [Google Scholar]
- Jin, M.; Wang, X.; Ning, K.; Liu, N.; Zhang, D. Genetic characterization of a new astrovirus in goslings suffering from gout. Arch. Virol. 2018, 163, 2865–2869. [Google Scholar] [CrossRef]
- Cuenca-Sanchez, M.; Navas-Carrillo, D.; Orenes-Pinero, E. Controversies surrounding high-protein diet intake: Satiating effect and kidney and bone health. Adv. Nutr. 2015, 6, 260–266. [Google Scholar] [CrossRef]
- Bi, J. The reason analysis of gout in chicken. Hebei J. Anim. Husb. Vet. Med. 2016, 26, 41–42. (In Chinese) [Google Scholar]
- Xi, Y.; Yan, J.; Li, M.; Ying, S.; Shi, Z. Gut microbiota dysbiosis increases the risk of visceral gout in goslings through translocation of gut-derived lipopolysaccharide. Poult. Sci. 2019, 98, 5361–5373. [Google Scholar] [CrossRef]
- Shen, G.; Gong, G.F.; Lv, S.Y.; Wang, Z.Q.; Liang, Z.; Gao, H.J.; Yu, M.S. Current waterfowl production and future tendency in China. Waterfowl World 2011, 5, 7–12. (In Chinese) [Google Scholar]
- Sun, A.D.; Shi, Z.D.; Huang, Y.M.; Liang, S.D. Development of geese out-of season lay technique and its impact on goose industry in Guangdong Province, China. World’s Poult. Sci. J. 2007, 64, 481–490. [Google Scholar] [CrossRef]
- Andriamihaja, M.; Davila, A.M.; Eklou-Lawson, M.; Petit, N.; Delpal, S.; Allek, F.; Blais, A.; Delteil, C.; Tome, D.; Blachier, F. Colon luminal content and epithelial cell morphology are markedly modified in rats fed with a high-protein diet. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 299, 1030–1037. [Google Scholar] [CrossRef] [Green Version]
- Windey, K.; De Preter, V.; Verbeke, K. Relevance of protein fermentation to gut health. Mol. Nutr. Food Res. 2012, 56, 184–196. [Google Scholar] [CrossRef]
- Coppo, R. The Gut-Renal Connection in IgA Nephropathy. Semin. Nephrol. 2018, 38, 504–512. [Google Scholar] [CrossRef] [PubMed]
- Tang, R.; Jiang, Y.; Tan, A.; Ye, J.; Xian, X.; Xie, Y.; Wang, Q.; Yao, Z.; Mo, Z. 16S rRNA gene sequencing reveals altered composition of gut microbiota in individuals with kidney stones. Urolithiasis 2018, 46, 503–514. [Google Scholar] [CrossRef]
- Shao, T.; Li, S.; Li, H.; Xie, Z.; He, Z.; Wen, C. Combined Signature of the Fecal Microbiome and Metabolome in Patients with Gout. Front. Microbiol. 2017, 8, 268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, J.; Zhou, B.; Xi, Y.; Huan, H.; Li, M.; Yu, J.; Zhu, H.; Dai, Z.; Ying, S.; Zhou, W.; et al. Fermented feed regulates growth performance and the cecal microbiota community in geese. Poult. Sci. 2019, 98, 4673–4684. [Google Scholar] [CrossRef] [PubMed]
- Canibe, N.; Hojberg, O.; Badsberg, J.H.; Jensen, B.B. Effect of feeding fermented liquid feed and fermented grain on gastrointestinal ecology and growth performance in piglets. J. Anim. Sci. 2007, 85, 2959–2971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ranjitkar, S.; Karlsson, A.H.; Petersen, M.A.; Bredie, W.L.; Petersen, J.S.; Engberg, R.M. The influence of feeding crimped kernel maize silage on broiler production, nutrient digestibility and meat quality. Br. Poult. Sci. 2016, 57, 93–104. [Google Scholar] [CrossRef]
- Xu, F.Z.; Zeng, X.G.; Ding, X.L. Effects of replacing soybean meal with fermented rapeseed meal on performance, serum biochemical variables and intestinal morphology of broilers. Asian-Australas. J. Anim. Sci. 2012, 25, 1734–1741. [Google Scholar] [CrossRef]
- Zhang, J.; Zhu, J.; Sun, J.; Li, Y.; Wang, P.; Jiang, R. Effect of fermented feed on intestinal morphology, immune status, carcass and growth performance of Emei Black chickens. FASEB J. 2016, 30, lb240. [Google Scholar]
- Hossain, M.A.; Lee, S.J.; Park, N.H.; Birhanu, B.T.; Mechesso, A.F.; Park, J.Y.; Park, E.J.; Lee, S.P.; Youn, S.J.; Park, S.C. Enhancement of Lipid Metabolism and Hepatic Stability in Fat-Induced Obese Mice by Fermented Cucurbita moschata Extract. Evid. Based Complement. Alternat. Med. 2018, 2018, 3908453. [Google Scholar] [CrossRef] [Green Version]
- Friedman, A.N. High-protein diets: Potential effects on the kidney in renal health and disease. Am. J. Kidney Dis. 2004, 44, 950–962. [Google Scholar] [CrossRef]
- Guo, X.; Huang, K.; Tang, J. Clinicopathology of gout in growing layers induced by high calcium and high protein diets. Br. Poult. Sci. 2005, 46, 641–646. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Huang, K.; Chen, F.; Luo, J.; Pan, C. High dietary calcium causes metabolic alkalosis in egg-type pullets. Poult. Sci. 2008, 87, 1353–1357. [Google Scholar] [CrossRef] [PubMed]
- Reginato, A.M.; Mount, D.B.; Yang, I.; Choi, H.K. The genetics of hyperuricaemia and gout. Nat. Rev. Rheumatol. 2012, 8, 610–621. [Google Scholar] [CrossRef] [PubMed]
- Hocking, P.M. Effect of dietary crude protein concentration on semen yield and quality in male broiler breeder fowls. Br. Poult. Sci. 1989, 30, 935–945. [Google Scholar] [CrossRef]
- Baum, N.; Dichoso, C.C.; Carlton, C.E. Blood urea nitrogen and serum creatinine: Physiology and interpretations. Urology 1975, 5, 583–588. [Google Scholar] [CrossRef]
- Huang, X.; Li, S.; Yang, X.; Peng, Q.; Wang, J.; Mo, C.; Wu, J.; Sui, J.; Liu, Y.; Lu, Y.; et al. Establishing reference values for blood urea nitrogen and serum creatinine in Chinese Han ethnic adult men. Clin. Lab. 2014, 60, 1123–1128. [Google Scholar] [CrossRef]
- Lin, H.; Chen, Y.; Huang, Q.; Guo, X.; Liu, P.; Liu, W.; Zhang, C.; Cao, H.; Hu, G. Prokaryotic expression of the chicken xanthine oxidase (XOD) subunit and its localization in liver and kidney. Int. J. Biol. Macromol. 2016, 87, 341–347. [Google Scholar] [CrossRef]
- Wong, K.H.; Hynes, M.J.; Davis, M.A. Recent advances in nitrogen regulation: A comparison between Saccharomyces cerevisiae and filamentous fungi. Eukaryot. Cell 2008, 7, 917–925. [Google Scholar] [CrossRef] [Green Version]
- Knight, E.L.; Stampfer, M.J.; Hankinson, S.E.; Spiegelman, D.; Curhan, G.C. The impact of protein intake on renal function decline in women with normal renal function or mild renal insufficiency. Ann. Intern. Med. 2003, 138, 460–467. [Google Scholar] [CrossRef] [Green Version]
- Marckmann, P.; Osther, P.; Pedersen, A.N.; Jespersen, B. High-protein diets and renal health. J. Ren. Nutr. 2015, 25, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Khoury, T.; Tzukert, K.; Abel, R.; Abu Rmeileh, A.; Levi, R.; Ilan, Y. The gut-kidney axis in chronic renal failure: A new potential target for therapy. Hemodial. Int. 2017, 21, 323–334. [Google Scholar] [CrossRef] [PubMed]
- Lan, A.; Andriamihaja, M.; Blouin, J.M.; Liu, X.; Descatoire, V.; Desclee de Maredsous, C.; Davila, A.M.; Walker, F.; Tome, D.; Blachier, F. High-protein diet differently modifies intestinal goblet cell characteristics and mucosal cytokine expression in ileum and colon. J. Nutr. Biochem. 2015, 26, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.D.; Chen, J.; Hoffmann, C.; Bittinger, K.; Chen, Y.Y.; Keilbaugh, S.A.; Bewtra, M.; Knights, D.; Walters, W.A.; Knight, R.; et al. Linking long-term dietary patterns with gut microbial enterotypes. Science 2011, 334, 105–108. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Zhou, H.; Pan, X.; Xu, T.; Zhang, Z.; Zi, X.; Jiang, Y. Cassava foliage affects the microbial diversity of Chinese indigenous geese caecum using 16S rRNA sequencing. Sci. Rep. 2017, 7, 45697. [Google Scholar] [CrossRef]
- Xu, Q.; Yuan, X.; Gu, T.; Li, Y.; Dai, W.; Shen, X.; Song, Y.; Zhang, Y.; Zhao, W.; Chang, G.; et al. Comparative characterization of bacterial communities in geese fed all-grass or high-grain diets. PLoS ONE 2017, 12, e0185590. [Google Scholar] [CrossRef] [Green Version]
- Guo, Z.; Zhang, J.; Wang, Z.; Ang, K.Y.; Huang, S.; Hou, Q.; Su, X.; Qiao, J.; Zheng, Y.; Wang, L. Intestinal Microbiota Distinguish Gout Patients from Healthy Humans. Sci. Rep. 2016, 6, 20602. [Google Scholar] [CrossRef] [Green Version]
- Lehto, M.; Groop, P.H. The Gut-Kidney Axis: Putative Interconnections Between Gastrointestinal and Renal Disorders. Front. Endocrinol. 2018, 9, 553. [Google Scholar] [CrossRef]
- Huang, W.; Guo, H.L.; Deng, X.; Zhu, T.T.; Xiong, J.F.; Xu, Y.H.; Xu, Y. Short-Chain Fatty Acids Inhibit Oxidative Stress and Inflammation in Mesangial Cells Induced by High Glucose and Lipopolysaccharide. Exp. Clin. Endocrinol. Diabetes 2017, 125, 98–105. [Google Scholar] [CrossRef]
- Maasjost, J.; Muhldorfer, K.; Cortez de Jackel, S.; Hafez, H.M. Antimicrobial Susceptibility Patterns of Enterococcus faecalis and Enterococcus faecium Isolated from Poultry Flocks in Germany. Avian Dis. 2015, 59, 143–148. [Google Scholar] [CrossRef]
- Liu, X.; Blouin, J.M.; Santacruz, A.; Lan, A.; Andriamihaja, M.; Wilkanowicz, S.; Benetti, P.H.; Tome, D.; Sanz, Y.; Blachier, F.; et al. High-protein diet modifies colonic microbiota and luminal environment but not colonocyte metabolism in the rat model: The increased luminal bulk connection. Am. J. Physiol. Liver Physiol. 2014, 307, G459–G470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macfarlane, G.T.; Macfarlane, S. Fermentation in the human large intestine: Its physiologic consequences and the potential contribution of prebiotics. J. Clin. Gastroenterol. 2011, 45, S120–S127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alemany, M. The problem of nitrogen disposal in the obese. Nutr. Res. Rev. 2012, 25, 18–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Compare, D.; Coccoli, P.; Rocco, A.; Nardone, O.M.; De Maria, S.; Carteni, M.; Nardone, G. Gut--liver axis: The impact of gut microbiota on non alcoholic fatty liver disease. Nutr. Metab. Cardiovasc. Dis. 2012, 22, 471–476. [Google Scholar] [CrossRef] [PubMed]
- Sugiharto, S.; Ranjitkar, S. Recent advances in fermented feeds towards improved broiler chicken performance, gastrointestinal tract microecology and immune responses: A review. Anim. Nutr. 2019, 5, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Engberg, R.M.; Hammershoj, M.; Johansen, N.F.; Abousekken, M.S.; Steenfeldt, S.; Jensen, B.B. Fermented feed for laying hens: Effects on egg production, egg quality, plumage condition and composition and activity of the intestinal microflora. Br. Poult. Sci. 2009, 50, 228–239. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Yang, D.; Mei, L.; Yuan, L.; Xie, A.; Yuan, J. Screening and characterization of purine nucleoside degrading lactic acid bacteria isolated from Chinese sauerkraut and evaluation of the serum uric acid lowering effect in hyperuricemic rats. PLoS ONE 2014, 9, e105577. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CP Levels | ||||
|---|---|---|---|---|
| 160CP | 180CP | 200CP | 220CP | |
| Feed compositions | ||||
| Corn, g/kg | 523 | 496 | 491 | 468 |
| Bean pulp, g/kg | 200 | 214 | 255 | 300 |
| Fish meal, g/kg | − | 39 | 59 | 77 |
| Bran, g/kg | 78 | 79 | 62 | 38 |
| Rice bran, g/kg | 156 | 124 | 80 | 62 |
| Rice hulls, g/kg | 2 | 7 | 12 | 15 |
| CaCO3, g/kg | 15 | 15 | 15 | 15 |
| CaHPO4, g/kg | 13 | 13 | 13 | 12 |
| NaCl, g/kg | 3 | 3 | 3 | 3 |
| Vitamin/mineral premix 1, g/kg | 10 | 10 | 10 | 10 |
| Nutritional ingredients 2 | ||||
| Poultry ME, Kcal/kg | 2700 | 2700 | 2700 | 2700 |
| CP, g/kg | 160 | 180 | 200 | 220 |
| CF, g/kg | 38 | 38 | 38 | 38 |
| Calcium, g/kg | 10 | 10 | 10 | 10 |
| Phosphorus, g/kg | 4 | 4 | 4 | 4 |
| CN 2 | CNF | HP | HPF | |
|---|---|---|---|---|
| Feed compositions | ||||
| Corn, g/kg | 589 | 589 | 460 | 460 |
| Bean pulp, g/kg | 160 | 160 | 240 | 240 |
| Corn gluten meal, g/kg | 40 | 40 | 100 | 100 |
| Fish meal, g/kg | 0 | 0 | 50 | 50 |
| Bran, g/kg | 110 | 110 | 69 | 69 |
| Rice bran, g/kg | 30 | 30 | 30 | 30 |
| Rice hulls, g/kg | 30 | 30 | 20 | 20 |
| CaCO3, g/kg | 11 | 11 | 11 | 11 |
| CaHPO4, g/kg | 13 | 13 | 3 | 3 |
| NaCl, g/kg | 3 | 3 | 3 | 3 |
| Vitamin/mineral premix 1, g/kg | 10 | 10 | 10 | 10 |
| Basal diet supplemented with 5.0% fermented feed | − | + | − | + |
| Nutritional ingredients | ||||
| Poultry ME, KJ/kg | 11.2 | 11.3 | 11.4 | 11.4 |
| CP, g/kg | 162.9 | 167.5 | 229.7 | 230.7 |
| CF, g/kg | 41.3 | 38.2 | 39.5 | 33.8 |
| Crude Fat, g/kg | 20.7 | 19.7 | 27.3 | 19.5 |
| Calcium, g/kg | 11.6 | 11.2 | 11.9 | 11.9 |
| Phosphorus, g/kg | 6 | 6.1 | 7.2 | 7.4 |
| Indexes | Groups | SEM | p-Value | |||
|---|---|---|---|---|---|---|
| 160CP | 180CP | 200CP | 220CP | |||
| Kidney function, Day 10 1 | ||||||
| UA, μmol·L−1 | 94.57 ab | 72.83 a | 106.52 ab | 210.87 b | 43.06 | 0.04 |
| Cr, μmol·L−1 | 17.70 | 19.65 | 18.26 | 19.95 | 1.30 | 0.57 |
| UN, mmol·L−1 | 0.89 | 2.05 | 1.28 | 1.53 | 0.29 | 0.09 |
| XOD, U·L−1 | 4.84 | 4.79 | 4.18 | 5.19 | 0.39 | 0.36 |
| Kidney function, Day 15 1 | ||||||
| UA, μmol·L−1 | 248.50 Aa | 294.72 ABab | 350.99 ABbc | 400.22 Bcd | 31.48 | < 0.01 |
| Cr, μmol·L−1 | 10.37 a | 11.16 a | 13.67 ab | 18.68 b | 2.28 | 0.04 |
| UN, mmol·L−1 | 0.38 a | 0.91 b | 1.01 b | 1.04 b | 0.17 | 0.02 |
| XOD, U·L−1 | 4.42 a | 5.15 ab | 5.62 ab | 6.28 b | 0.80 | 0.03 |
| Indexes | Groups 1 | SEM | p-Value | |||||
|---|---|---|---|---|---|---|---|---|
| CN | CNF | HP | HPF | HP | F | HP*F | ||
| Day 7 | ||||||||
| UA, μmol·L−1 | 407.12 | 276.12 | 578.38 | 392.25 | 28.96 | 0.004 | 0.002 | 0.548 |
| Cr, μmol·L−1 | 18.89 | 22.01 | 19.24 | 16.70 | 1.33 | 0.367 | 0.917 | 0.303 |
| UN, mmol·L−1 | 1.54 | 1.96 | 1.83 | 1.53 | 0.09 | 0.697 | 0.740 | 0.143 |
| XOD, U·L−1 | 6.09 | 6.92 | 10.71 | 6.17 | 0.55 | 0.040 | 0.048 | 0.106 |
| Day 14 | ||||||||
| UA, μmol·L−1 | 289.25 | 151.38 | 444.75 | 251.12 | 30.94 | 0.019 | 0.003 | 0.592 |
| Cr, μmol·L−1 | 10.10 | 9.40 | 14.13 | 10.47 | 0.77 | 0.038 | 0.064 | 0.762 |
| UN, mmol·L−1 | 1.62 | 1.18 | 1.90 | 1.02 | 0.10 | 0.723 | 0.001 | 0.200 |
| XOD, U·L−1 | 6.99 | 6.95 | 8.88 | 6.85 | 0.31 | 0.853 | 0.092 | 0.103 |
| Day 21 | ||||||||
| UA, μmol·L−1 | 202.88 | 137.62 | 247.25 | 147.75 | 22.96 | 0.553 | 0.080 | 0.708 |
| Cr, μmol·L−1 | 7.57 | 7.08 | 14.45 | 5.63 | 0.97 | 0.095 | 0.006 | 0.113 |
| UN, mmol·L−1 | 1.03 | 0.88 | 1.17 | 0.87 | 0.06 | 0.588 | 0.070 | 0.509 |
| XOD, U·L−1 | 6.07 | 5.23 | 8.48 | 4.77 | 0.38 | 0.116 | 0.001 | 0.124 |
| Day 35 | ||||||||
| UA, μmol·L−1 | 382.83 | 293.00 | 813.00 | 780.83 | 55.24 | 0.000 | 0.299 | 0.620 |
| Cr, μmol·L−1 | 11.34 | 5.84 | 21.44 | 13.43 | 1.46 | 0.000 | 0.002 | 0.509 |
| UN, mmol·L−1 | 1.15 | 1.00 | 1.80 | 1.24 | 0.11 | 0.035 | 0.083 | 0.307 |
| XOD, U·L−1 | 5.91 | 7.66 | 5.66 | 5.25 | 0.44 | 0.127 | 0.431 | 0.211 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xi, Y.; Huang, Y.; Li, Y.; Yan, J.; Shi, Z. Fermented Feed Supplement Relieves Caecal Microbiota Dysbiosis and Kidney Injury Caused by High-Protein Diet in the Development of Gosling Gout. Animals 2020, 10, 2139. https://doi.org/10.3390/ani10112139
Xi Y, Huang Y, Li Y, Yan J, Shi Z. Fermented Feed Supplement Relieves Caecal Microbiota Dysbiosis and Kidney Injury Caused by High-Protein Diet in the Development of Gosling Gout. Animals. 2020; 10(11):2139. https://doi.org/10.3390/ani10112139
Chicago/Turabian StyleXi, Yumeng, Yuanpi Huang, Ying Li, Junshu Yan, and Zhendan Shi. 2020. "Fermented Feed Supplement Relieves Caecal Microbiota Dysbiosis and Kidney Injury Caused by High-Protein Diet in the Development of Gosling Gout" Animals 10, no. 11: 2139. https://doi.org/10.3390/ani10112139