Effects of Dietary Supplementation of Garlic and Oregano Essential Oil on Biomarkers of Oxidative Status, Stress and Inflammation in Postweaning Piglets

 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Additives and Feed Composition
2.2. Animals, Housing, and Experimental Design
2.3. Sample Collection and Productive Measures
2.4. Measurements of Biomarkers
2.4.1. Oxidative Status
2.4.2. Salivary Cortisol
2.4.3. Serum CRP
2.5. Statistical Analysis
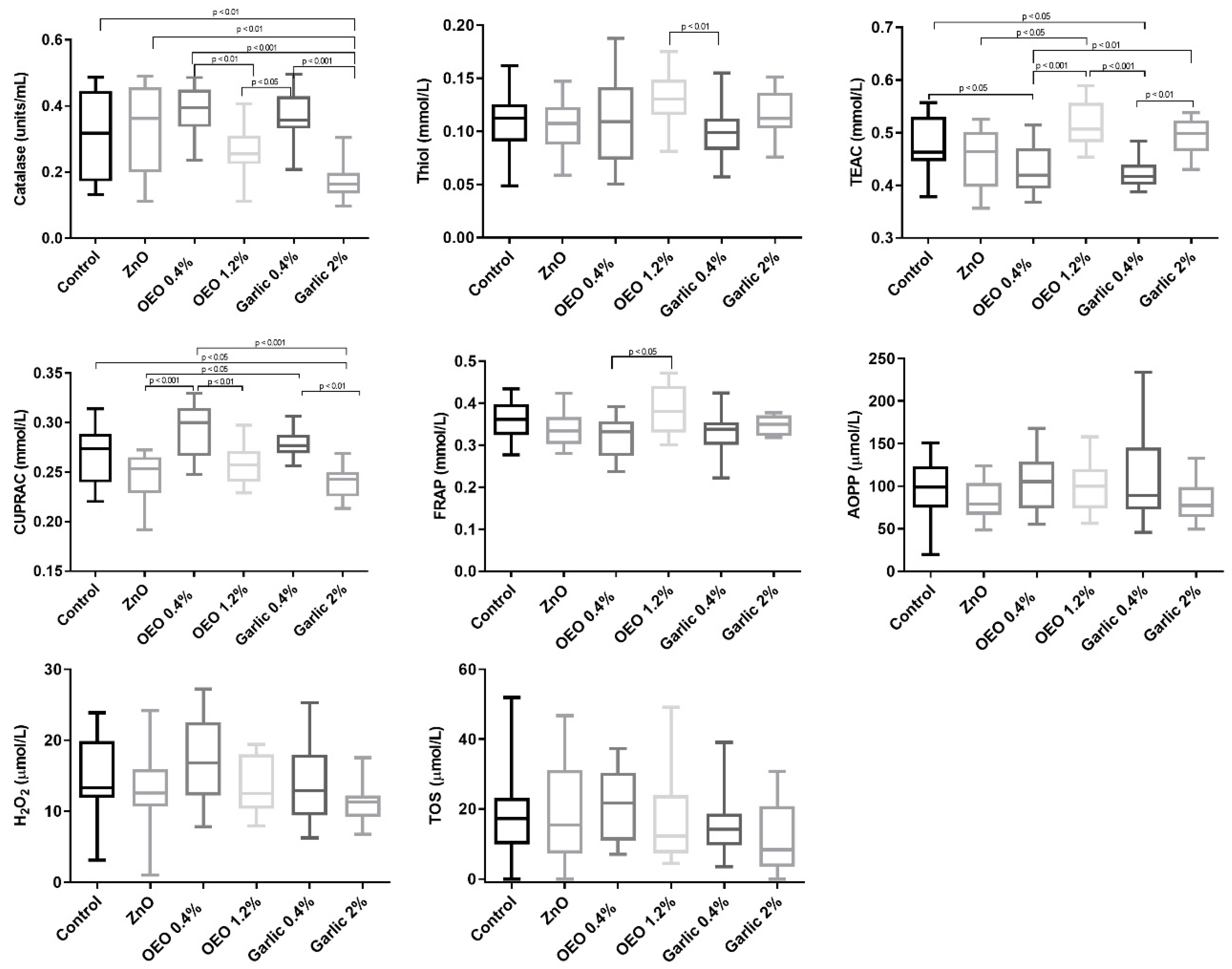
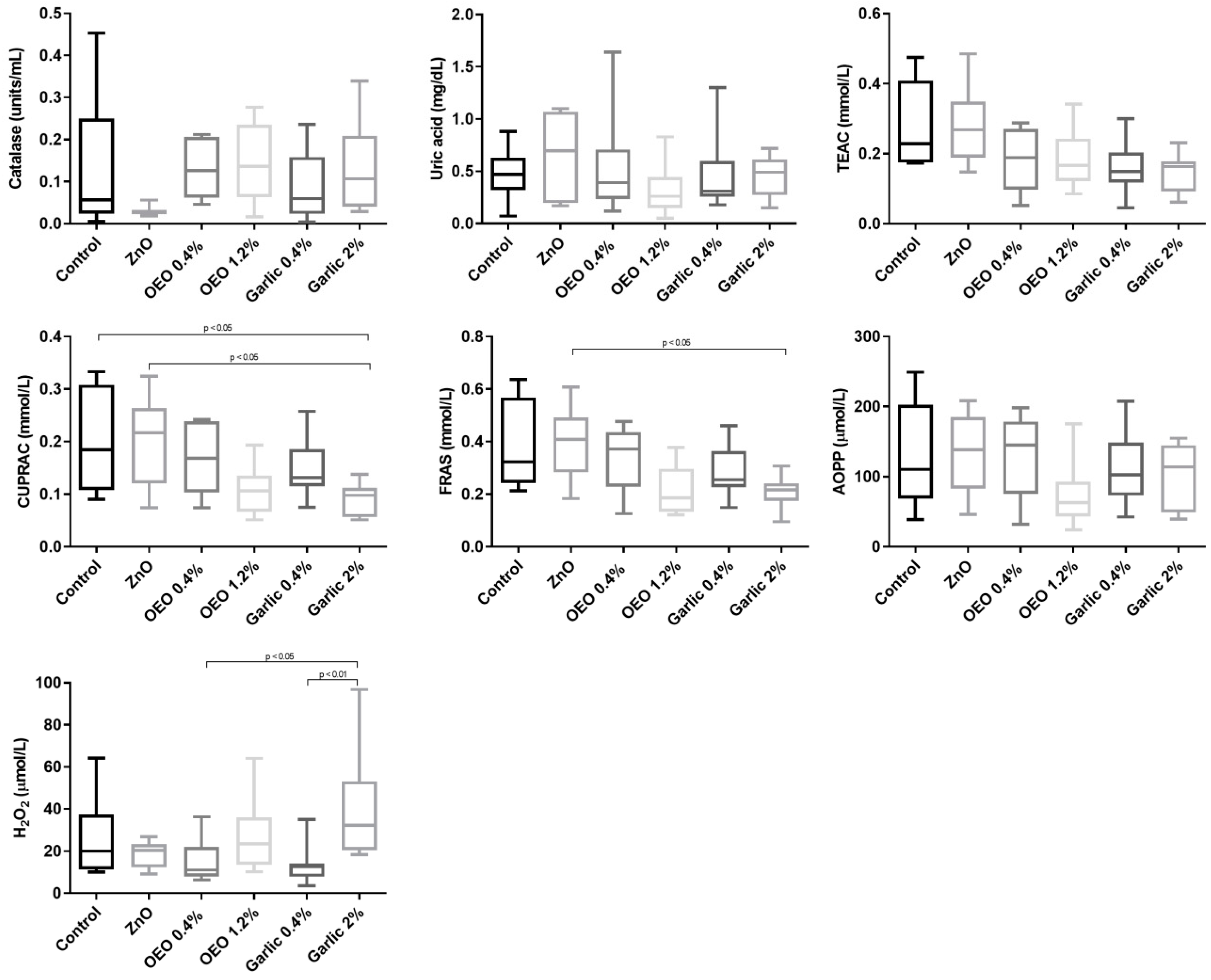
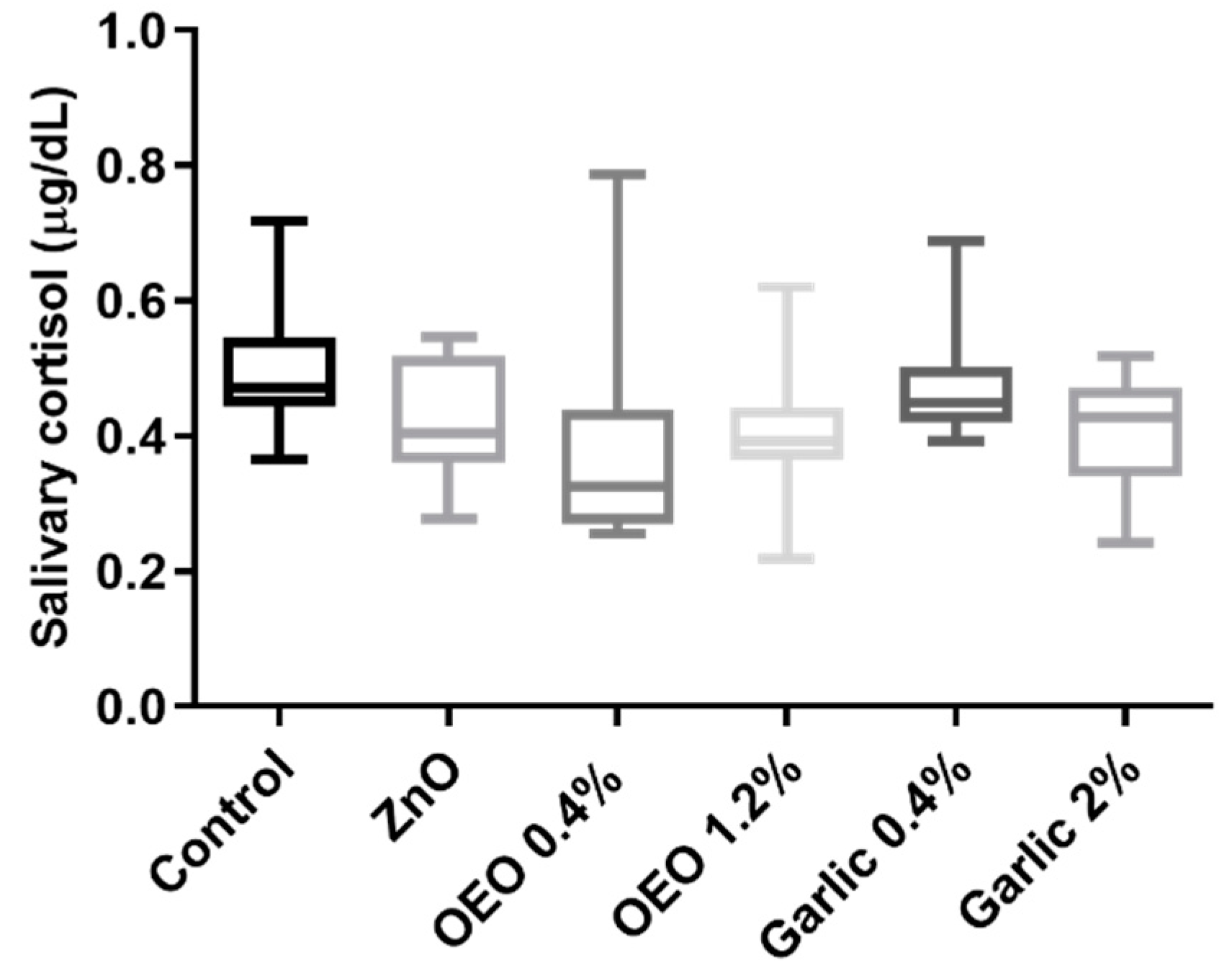
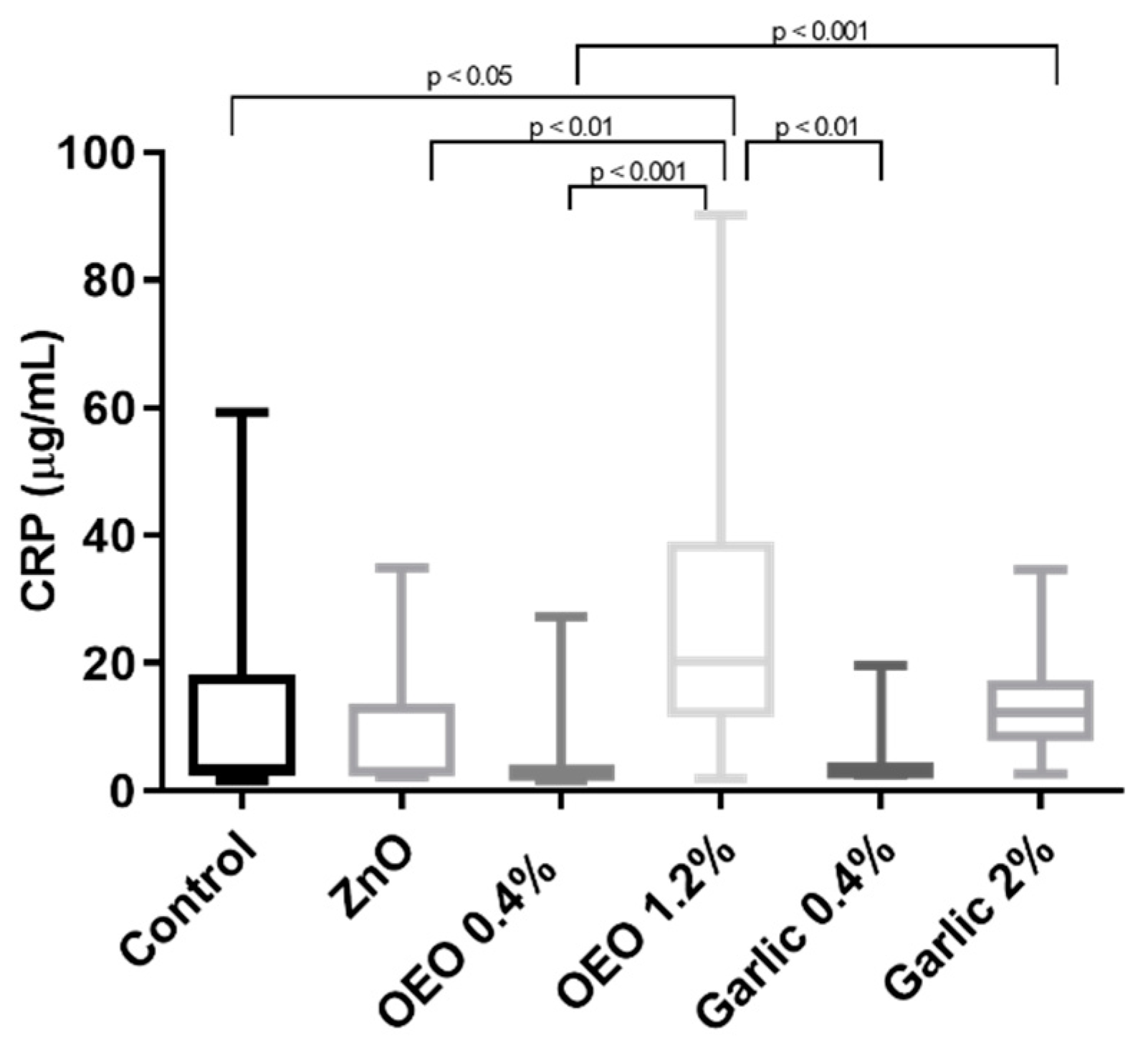
3. Results
3.1. Biomarkers of Oxidative Stress
3.2. Salivary Cortisol
3.3. Serum CRP
3.4. Correlations Between Serum and Saliva
3.5. Productive Parameters
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Moberg, G.P.; Mench, J.A. The Biology of Animal Stress: Basic Principles and Implications for Animal Welfare; CAB International: Wallingford, UK, 2000. [Google Scholar]
- Srinongkote, S.; Smriga, M.; Nakagawa, K.; Toride, Y. A Diet Fortified withl-lysine andl-arginine Reduces Plasma Cortisol and Blocks Anxiogenic Response to Transportation in Pigs. Nutr. Neurosci. 2003, 6, 283–289. [Google Scholar] [CrossRef]
- Gupta, S.; Earley, B.; Crowe, M. Effect of 12-hour road transportation on physiological, immunological and haematological parameters in bulls housed at different space allowances. Veter J. 2007, 173, 605–616. [Google Scholar] [CrossRef]
- Sporer, K.B.; Weber, P.S.D.; Burton, J.L.; Earley, B.; Crowe, M. Transportation of young beef bulls alters circulating physiological parameters that may be effective biomarkers of stress1. J. Anim. Sci. 2008, 86, 1325–1334. [Google Scholar] [CrossRef]
- Escribano, D.; Ko, H.-L.; Chong, Q.; Llonch, L.; Manteca, X.; Llonch, P. Salivary biomarkers to monitor stress due to aggression after weaning in piglets. Res. Veter Sci. 2019, 123, 178–183. [Google Scholar] [CrossRef]
- Zhu, H.; Jia, Z.; Misra, H.; Li, Y.R. Oxidative stress and redox signaling mechanisms of alcoholic liver disease: Updated experimental and clinical evidence. J. Dig. Dis. 2012, 13, 133–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pié, S.; Lallès, J.P.; Blazy, F.; Laffitte, J.; Sève, B.; Oswald, I.P. Weaning Is Associated with an Upregulation of Expression of Inflammatory Cytokines in the Intestine of Piglets. J. Nutr. 2004, 134, 641–647. [Google Scholar] [CrossRef] [Green Version]
- Wijtten, P.J.A.; Van Der Meulen, J.; Verstegen, M.W.A. Intestinal barrier function and absorption in pigs after weaning: A review. Br. J. Nutr. 2011, 105, 967–981. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Hansen, C.; Mullan, B.; Pluske, J. Nutrition and pathology of weaner pigs: Nutritional strategies to support barrier function in the gastrointestinal tract. Anim. Feed. Sci. Technol. 2012, 173, 3–16. [Google Scholar] [CrossRef] [Green Version]
- Hahn, J.D.; Baker, D.H. Growth and plasma zinc responses of young pigs fed pharmacologic levels of zinc. J. Anim. Sci. 1993, 71, 3020–3024. [Google Scholar] [CrossRef]
- Patel, A.; Mamtani, M.; Dibley, M.J.; Badhoniya, N.; Kulkarni, H. Therapeutic Value of Zinc Supplementation in Acute and Persistent Diarrhea: A Systematic Review. PLoS ONE 2010, 5, e10386. [Google Scholar] [CrossRef] [Green Version]
- Sargeant, H.R.; Miller, H.M.; Shaw, M.-A. Inflammatory response of porcine epithelial IPEC J2 cells to enterotoxigenic E. coli infection is modulated by zinc supplementation. Mol. Immunol. 2011, 48, 2113–2121. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Song, J.; Li, Y.; Luan, Z.; Zhu, K. Diosmectite–zinc oxide composite improves intestinal barrier function, modulates expression of pro-inflammatory cytokines and tight junction protein in early weaned pigs. Br. J. Nutr. 2013, 110, 681–688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoque, K.M.; Rajendran, V.M.; Binder, H.J. Zinc inhibits cAMP-stimulated Cl secretion via basolateral K-channel blockade in rat ileum. Am. J. Physiol. Liver Physiol. 2005, 288, G956–G963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Söderberg, T.A.; Sunzel, B.; Holm, S.; Elmros, T.; Hallmans, G.; Sjöberg, S. Antibacterial Effect of Zinc Oxide In Vitro. Scand. J. Plast. Reconstr. Surg. Hand Surg. 1990, 24, 193–197. [Google Scholar] [CrossRef]
- Poulsen, H.D.; Larsen, T. Zinc excretion and retention in growing pigs fed increasing levels of zinc oxide. Livest. Prod. Sci. 1995, 43, 235–242. [Google Scholar] [CrossRef]
- Sargeant, H.R.; McDowall, K.J.; Miller, H.M.; Shaw, M.-A. Dietary zinc oxide affects the expression of genes associated with inflammation: Transcriptome analysis in piglets challenged with ETEC K88. Veter Immunol. Immunopathol. 2010, 137, 120–129. [Google Scholar] [CrossRef]
- Cavaco, L.M.; Hasman, H.; Stegger, M.; Andersen, P.S.; Skov, R.; Fluit, A.C.; Ito, T.; Aarestrup, F.M. Cloning and occurrence of czrC, a gene conferring cadmium and zinc resistance in methicillin-resistant Staphylococcus aureus CC398 isolates. Antimicrob. Agents Chemother. 2010, 54, 3605–3608. [Google Scholar] [CrossRef] [Green Version]
- Cavaco, L.M.; Hasman, H.; Aarestrup, F.M. Zinc resistance of Staphylococcus aureus of animal origin is strongly associated with methicillin resistance. Veter Microbiol. 2011, 150, 344–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Standing Committee on Veterinary Medicinal Products. 2017. Brussels, 26.6.2017 C (2017) 4529 Final. 2017. Available online: https://ec.europa.eu/health/documents/community-register/2017/20170626136754/dec_136754_en.pdf (accessed on 14 April 2020).
- Allan, P.; Bilkei, G. Oregano improves reproductive performance of sows. Theriogenology 2005, 63, 716–721. [Google Scholar] [CrossRef] [PubMed]
- Pandey, A.; Rai, M.; Acharya, D. Chemical Composition and Antimycotic Activity of the Essential Oils of Corn Mint (Mentha arvensis) and Lemon Grass (Cymbopogon flexuosus) against Human Pathogenic Fungi. Pharm. Biol. 2003, 41, 421–425. [Google Scholar] [CrossRef]
- Daouk, R.K.; Dagher, S.M.; Sattout, E.J. Antifungal Activity of the Essential Oil of Origanum syriacum L. J. Food Prot. 1995, 58, 1147–1149. [Google Scholar] [CrossRef] [PubMed]
- Cervato, G.; Carabelli, M.; Gervasio, S.; Cittera, A.; Cazzola, R.; Cestaro, B. Antioxbdant properties of oregano (origanum vulgare) leaf extracts. J. Food Biochem. 2000, 24, 453–465. [Google Scholar] [CrossRef]
- Dorman, H.J.D.; Deans, S.G. Antimicrobial agents from plants: Antibacterial activity of plant volatile oils. J. Appl. Microbiol. 2000, 88, 308–316. [Google Scholar] [CrossRef] [PubMed]
- Marcin, A.; Lauková, A.; Mati, R. Comparison of the effects of Enterococcus faecium and aromatic oils from sage and oregano on growth performance and diarrhoeal diseases of weaned pigs. Biologia 2006, 61. [Google Scholar] [CrossRef]
- Neill, C.R.; Nelssen, J.L.; Tokach, M.D.; Goodband, R.D.; DeRouchey, J.M.; Dritz, S.S. Effects of oregano oil on growth performance of nursery pigs. J. Swine Heal. Prod. 2006, 14, 312–316. [Google Scholar]
- Bampidis, V.; Christodoulou, V.; Florou-Paneri, P.; Christaki, E.; Spais, A.; Chatzopoulou, P.S. Effect of dietary dried oregano leaves supplementation on performance and carcass characteristics of growing lambs. Anim. Feed. Sci. Technol. 2005, 121, 285–295. [Google Scholar] [CrossRef]
- Hernández, F.; Madrid, J.; García, V.; Orengo, J.; Megías, M.D. Influence of two plant extracts on broilers performance, digestibility, and digestive organ size. Poult. Sci. 2004, 83, 169–174. [Google Scholar] [CrossRef]
- Hong, J.-C.; Steiner, T.; Aufy, A.; Lien, T.-F. Effects of supplemental essential oil on growth performance, lipid metabolites and immunity, intestinal characteristics, microbiota and carcass traits in broilers. Livest. Sci. 2012, 144, 253–262. [Google Scholar] [CrossRef]
- Zhang, T.; Zhou, Y.; Zou, Y.; Hu, X.; Zheng, L.; Wei, H.; Giannenas, I.; Jin, L.; Peng, J.; Jiang, S. Effects of dietary oregano essential oil supplementation on the stress response, antioxidative capacity, and HSPs mRNA expression of transported pigs. Livest. Sci. 2015, 180, 143–149. [Google Scholar] [CrossRef]
- Leyva-López, N.; Gutiérrez-Grijalva, E.P.; Vazquez-Olivo, G.; Heredia, J.B. Essential Oils of Oregano: Biological Activity beyond Their Antimicrobial Properties. Molecules 2017, 22, 989. [Google Scholar] [CrossRef] [Green Version]
- Bakry, A.M.; Abbas, S.; Ali, B.; Majeed, H.; Abouelwafa, M.Y.; Mousa, A.; Liang, L. Microencapsulation of Oils: A Comprehensive Review of Benefits, Techniques, and Applications. Compr. Rev. Food Sci. Food Saf. 2015, 15, 143–182. [Google Scholar] [CrossRef]
- Amagase, H.; Petesch, B.L.; Matsuura, H.; Kasuga, S.; Itakura, Y. Intake of Garlic and Its Bioactive Components. J. Nutr. 2001, 131, 955S–962S. [Google Scholar] [CrossRef] [PubMed]
- Amagase, H. Clarifying the Real Bioactive Constituents of Garlic. J. Nutr. 2006, 136, 716S–725S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sallam, K.H.; Ishioroshi, I.M.; Samejima, K. Antioxid Antimicrob Eff garlic Chick sausage. Leb. Wiss U Technol. 2004, 37, 849–855. [Google Scholar] [CrossRef] [Green Version]
- Aydın, S.; Başaran, A.A.; Başaran, N.; Aydin, S. Modulating Effects of Thyme and Its Major Ingredients on Oxidative DNA Damage in Human Lymphocytes. J. Agric. Food Chem. 2005, 53, 1299–1305. [Google Scholar] [CrossRef]
- Li, W.-R.; Shi, Q.-S.; Dai, H.-Q.; Liang, Q.; Xie, X.-B.; Huang, X.-M.; Zhao, G.-Z.; Zhang, L. Antifungal activity, kinetics and molecular mechanism of action of garlic oil against Candida albicans. Sci. Rep. 2016, 6, 22805. [Google Scholar] [CrossRef]
- Tatara, M.R.; Sliwa, E.; Dudek, K.; Gawron, A.; Piersiak, T.; Dobrowolski, P.; Mosiewicz, J.; Siwicki, A.; Studzinski, T. Aged garlic extract and allicin improve performance and gastrointestinal tract development of piglets reared in artificial sow. Ann. Agric. Environ. Med. 2008, 15, 63–69. [Google Scholar]
- Huang, R.H.; Qiu, X.S.; Shi, F.X.; Hughes, C.L.; Lu, Z.F.; Zhu, W.Y. Effects of dietary allicin on health and growth performance of weanling piglets and reduction in attractiveness of faeces to flies. Animals 2011, 5, 304–311. [Google Scholar] [CrossRef]
- Wang, J.P.; Yoo, J.S.; Jang, H.D.; Lee, J.H.; Cho, J.H.; Kim, I.H. Effect of dietary fermented garlic by Weissella koreensis powder on growth performance, blood characteristics, and immune response of growing pigs challenged with Escherichia coli lipopolysaccharide. J. Anim. Sci. 2011, 89, 2123–2131. [Google Scholar] [CrossRef] [Green Version]
- Horn, N.; Miller, G.; Ajuwon, K.M.; Adeola, O. Ability of garlic-derived diallyl disulfide and diallyl trisulfide supplemented by oral gavage to mitigate effects of an acute post weaning feed and water deprivation event in nursery pigs. J. Anim. Sci. 2017, 95, 3579. [Google Scholar]
- Prieto, M.; Campo, J. Effect of heat and several additives related to stress levels on fluctuating asymmetry, heterophil:lymphocyte ratio, and tonic immobility duration in White Leghorn chicks. Poult. Sci. 2010, 89, 2071–2077. [Google Scholar] [CrossRef] [PubMed]
- Buchet, A.; Belloc, C.; Leblanc-Maridor, M.; Merlot, E. Effects of age and weaning conditions on blood indicators of oxidative status in pigs. PLoS ONE 2017, 12, e0178487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, Z.; Zhu, W.; Guo, Q.; Luo, W.; Zhang, J.; Xu, W.; Xu, J. Weaning Induced Hepatic Oxidative Stress, Apoptosis, and Aminotransferases through MAPK Signaling Pathways in Piglets. Oxidative Med. Cell. Longev. 2016, 2016, 1–10. [Google Scholar] [CrossRef]
- Rubio, C.P.; Mainau, E.; Cerón, J.J.; Contreras-Aguilar, M.D.; Martínez-Subiela, S.; Navarro, E.; Tecles, F.; Manteca, X.; Escribano, D. Biomarkers of oxidative stress in saliva in pigs: Analytical validation and changes in lactation. BMC Veter Res. 2019, 15, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, H.; Chen, G.; Wang, R.-J.; Peng, J. Oregano essential oil decreased susceptibility to oxidative stress-induced dysfunction of intestinal epithelial barrier in rats. J. Funct. Foods 2015, 18, 1191–1199. [Google Scholar] [CrossRef]
- Zou, Y.; Wang, J.; Peng, J.; Wei, H. Oregano Essential Oil Induces SOD1 and GSH Expression through Nrf2 Activation and Alleviates Hydrogen Peroxide-Induced Oxidative Damage in IPEC-J2 Cells. Oxidative Med. Cell. Longev. 2016, 2016, 1–13. [Google Scholar] [CrossRef]
- Liu, Q.; Duan, R.J.; Zhou, Y.F.; Wei, H.K.; Peng, J.; Li, J.L. Supplementing oregano essential oil to boar diet with strengthened fish oil: Effects on semen antioxidant status and semen quality parameters. Andrologia 2017, 49, e12764. [Google Scholar] [CrossRef]
- Cheng, C.; Zou, Y.; Peng, J. Oregano Essential Oil Attenuates RAW264.7 Cells from Lipopolysaccharide-Induced Inflammatory Response through Regulating NADPH Oxidase Activation-Driven Oxidative Stress. Molecules 2018, 23, 1857. [Google Scholar] [CrossRef] [Green Version]
- Peng, J.; Liu, Z.; Zhou, Y.; Wei, H.; Zhang, X.; Xiaming, Z.; Deng, Z.; Zou, Y.; Jiang, S.; Peng, J. Effect of oregano essential oil supplementation to a reduced-protein, amino acid-supplemented diet on meat quality, fatty acid composition, and oxidative stability of Longissimus thoracis muscle in growing-finishing pigs. Meat Sci. 2017, 133, 103–109. [Google Scholar] [CrossRef]
- Peng, J.; Xia, M.; Zhang, X.; Wang, C.; Jiang, S.; Peng, J. Supplementing Oregano Essential Oil in a Reduced-Protein Diet Improves Growth Performance and Nutrient Digestibility by Modulating Intestinal Bacteria, Intestinal Morphology, and Antioxidative Capacity of Growing-Finishing Pigs. Animals 2018, 8, 159. [Google Scholar] [CrossRef] [Green Version]
- Rubio, C.P.; Hernández-Ruiz, J.; Martinez-Subiela, S.; Tvarijonaviciute, A.; Ceron, J.J. Spectrophotometric assays for total antioxidant capacity (TAC) in dog serum: An update. BMC Veter Res. 2016, 12, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- National Research Council. Nutrient Requirements of Swine; The National Academies Press: Washington, DC, USA, 2012; pp. 1–399. [Google Scholar]
- European Commission. Council Directive 98/58/EC of 20 July 1998 concerning the protection of animals kept for farming purposes. Off. J. 1998, L 221, 23–27. [Google Scholar]
- European Commission. Council Directive 2008/120/EC of 18 December 2008 laying down minimum standards for the protection of pigs. Off. J. 2008, L 47, 5–13. Available online: http://eur-lex.europa.eu/legal-content/EN/TXT/?qid=1487548100586&uri=CELEX:32008L0120 (accessed on 20 February 2020).
- Jocelyn, P.C. Spectrophotometric assay of thiols. Methods Enzymol. 1987, 143, 44–67. [Google Scholar] [CrossRef] [PubMed]
- Da Costa, C.M.; Dos Santos, R.C.C.; Lima, E.S. A simple automated procedure for thiol measurement in human serum samples. J. Bras. Patol. Med. Lab. 2006, 42, 345–350. [Google Scholar] [CrossRef]
- Slaughter, M.R.; O’Brien, P.J. Fully-automated spectrophotometric method for measurement of antioxidant activity of catalase. Clin. Biochem. 2000, 33, 525–534. [Google Scholar] [CrossRef]
- Arnao, M.; Caño, A.; Hernández-Ruiz, J.; García-Cánovas, F.; Acosta, M. Inhibition byl-Ascorbic Acid and Other Antioxidants of the 2,2′-Azino-bis(3-ethylbenzthiazoline-6-sulfonic Acid) Oxidation Catalyzed by Peroxidase: A New Approach for Determining Total Antioxidant Status of Foods. Anal. Biochem. 1996, 236, 255–261. [Google Scholar] [CrossRef]
- Campos, C.; Guzmán, R.; López-Fernández, E.; Casado, Á. Evaluation of the copper(II) reduction assay using bathocuproinedisulfonic acid disodium salt for the total antioxidant capacity assessment: The CUPRAC–BCS assay. Anal. Biochem. 2009, 392, 37–44. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [Green Version]
- Erel, O. A new automated colorimetric method for measuring total oxidant status. Clin. Biochem. 2005, 38, 1103–1111. [Google Scholar] [CrossRef]
- Witko-Sarsat, V.; Friedlander, M.; Capeillère-Blandin, C.; Nguyen-Khoa, T.; Nguyen, A.T.; Zingraff, J.; Jungers, P.; Descamps-Latscha, B. Advanced oxidation protein products as a novel marker of oxidative stress in uremia. Kidney Int. 1996, 49, 1304–1313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rhee, S.G.; Chang, T.-S.; Jeong, W.; Kang, D. Methods for detection and measurement of hydrogen peroxide inside and outside of cells. Mol. Cells 2010, 29, 539–549. [Google Scholar] [CrossRef] [PubMed]
- Escribano, D.; Fuentes-Rubio, M.; Cerón, J.J. Validation of an automated chemiluminescent immunoassay for salivary cortisol measurements in pigs. J. Veter Diagn. Investig. 2012, 24, 918–923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernández-Caravaca, I.; Gourgues, S.F.; Rodríguez, V.; Estrada, E.D.; Cerón, J.J.; Escribano, D.; Gourgues, S.F.; Rodriguez-Vega, V. Serum acute phase response induced by different vaccination protocols against circovirus type 2 and Mycoplasma hyopneumoniae in piglets. Res. Veter Sci. 2017, 114, 69–73. [Google Scholar] [CrossRef]
- Sauerwein, H.; Schmitz, S.; Hiss, S. The acute phase protein haptoglobin and its relation to oxidative status in piglets undergoing weaning-induced stress. Redox Rep. 2005, 10, 295–302. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Zhao, X.; Jiang, X.; Chen, L.; Hong, L.; Zhuo, Y.; Lin, Y.; Fang, Z.; Che, L.; Feng, B.; et al. Effects of dietary supplementation with exogenous catalase on growth performance, oxidative stress, and hepatic apoptosis in weaned piglets challenged with lipopolysaccharide. J. Anim. Sci. 2020, 98. [Google Scholar] [CrossRef]
- Hemken, R.; Bremel, D. Possible Role of Beta-Carotene in Improving Fertility in Dairy Cattle. J. Dairy Sci. 1982, 65, 1069–1073. [Google Scholar] [CrossRef]
- Lauridsen, C.; Theil, P.K.; Jensen, S.K. Composition of α-tocopherol and fatty acids in porcine tissues after dietary supplementation with vitamin E and different fat sources. Anim. Feed. Sci. Technol. 2013, 179, 93–102. [Google Scholar] [CrossRef]
- Shaish, A.; Daugherty, A.; O’Sullivan, F.; Schonfeld, G.; Heinecke, J.W. Beta-carotene inhibits atherosclerosis in hypercholesterolemic rabbits. J. Clin. Investig. 1995, 96, 2075–2082. [Google Scholar] [CrossRef]
- Munday, R.; Munday, J.S.; Munday, C.M. Comparative effects of mono-, di-, tri-, and tetrasulfides derived from plants of the Allium family: Redox cycling in vitro and hemolytic activity and Phase 2 enzyme induction in vivo. Free Radic. Biol. Med. 2003, 34, 1200–1211. [Google Scholar] [CrossRef]
- Vega Montalvo, R.; Carrillo Domínguez, C. Efecto sobre la motilidad intestinal y toxicidad aguda oral del extracto fluido de Ocimum gratissimum L. (orégano cimarrón). Rev. Cuba Plantas Med. 1997, 2, 14–18. [Google Scholar]
- Zou, Y.; Xiang, Q.; Wang, J.; Peng, J.; Wei, H. Oregano Essential Oil Improves Intestinal Morphology and Expression of Tight Junction Proteins Associated with Modulation of Selected Intestinal Bacteria and Immune Status in a Pig Model. BioMed Res. Int. 2016, 2016, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, L.; Kim, I.H. Effects of dietary supplementation of fermented garlic powder on growth performance, apparent total tract digestibility, blood characteristics and faecal microbial concentration in weanling pigs. J. Anim. Physiol. Anim. Nutr. 2012, 97, 457–464. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.; Liu, S.; Kim, I.H. Effects of dietary Korean aged garlic extract by Leuconostoc mesenteroides KCCM35046 on growth performance, digestibility, blood profiles, gas emissions and microbiotain weanling pigs. Can. J. Anim. Sci. 2020. [Google Scholar] [CrossRef]
- Liu, Y.; Song, M.; Che, T.M.; Lee, J.J.; Bravo, D.; Maddox, C.W.; Pettigrew, J.E. Dietary plant extracts modulate gene expression profiles in ileal mucosa of weaned pigs after an Escherichia coli infection1. J. Anim. Sci. 2014, 92, 2050–2062. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Components | % |
|---|---|
| α-Pinene | 0.84 |
| α-Thuyene | 1.10 |
| Camphene | 0.07 |
| β-Pinene | 0.23 |
| β-Mircene | 1.56 |
| α-Phellandrene | 0.22 |
| α-Terpinene | 1.25 |
| Limonene | 0.31 |
| 1,8-Cineole+ β-Phellandrene | 0.30 |
| γ-Terpinene | 5.20 |
| 3-Octonone | 0.11 |
| p-Cymene | 5.99 |
| Terpinolene | 0.12 |
| 1-Octen-3-ol | 0.26 |
| (E)-Thuyanol | 0.10 |
| Linalool | 1.73 |
| (Z)-Thuyanol | 0.05 |
| 1-Octanol | 0.02 |
| Terpinene-4-ol | 0.79 |
| β-Caryophyllene | 2.59 |
| α-Humulene | 0.12 |
| α-Terpineol | 0.16 |
| Borneol | 0.22 |
| β-Bisabolene | 0.23 |
| Caryophellene oxyde | 0.14 |
| Thymol | 4.10 |
| Carvacrol Isomere | 0.05 |
| Carvacrol | 70.32 |
| Total | 98.19 |
| Analytical Components | Quantity |
|---|---|
| Protein (%) | 3.9 |
| Carbohydrates (%) | 15.4 |
| Sugars (%) | 1.4 |
| Crude Fibre (%) | 0.7 |
| Crude Fat (%) | 0.2 |
| Crude Ash (%) | 35.4 |
| Humidity (%) | 44.4 |
| Inuline (%) | 8.1 |
| Energy (kJ/kg) | 341 |
| Aminoacids (g/kg) | |
| Methionine | 0.5 |
| Lysine | 2.1 |
| Isoleucine | 0.9 |
| Leucine | 1.5 |
| Valine | 1.4 |
| Phenylalanine | 1.1 |
| Tryptophan | 0.4 |
| Threonine | 1.1 |
| Histidine | 0.6 |
| Arginine | 8.8 |
| Minerals (mg/kg) | |
| Calcium | 1100 |
| Phosphorus | 1060 |
| Potassium | 3500 |
| Sodium | 7247 |
| Copper | 1.6 |
| Cobalt | 0.12 |
| Iron | 20 |
| Manganese | 13.2 |
| Selenium | 0.2 |
| Zinc | 6.3 |
| Sulfur | 6000 |
| Vitamins | |
| Thiamine (mg/kg) | 2.6 |
| Niacin (mg/kg) | 17 |
| Ascorbic acid (mg/kg) | 116.5 |
| Cholecalciferol (µg) | 4 |
| Riboflavin (mg/kg) | 0.3 |
| Cyanocobalamin (µg) | 5 |
| Retinol (mg/kg) | 0.23 |
| Tocopherol (mg/kg) | 3 |
| Ingredients Prestarter (g/kg) | Prestarter | Starter |
|---|---|---|
| Maize | 299.8 | |
| Wheat | 200 | |
| Soybean | 183.5 | |
| Soybean meal | 135 | |
| Barley | 80 | |
| Whey | 75 | |
| Vitamins and oligoelements premix | 25 | |
| Lysine | 0.75 | |
| Threonine | 0.6 | |
| Tryptophan | 0.2 | |
| Methionine | 0.15 | |
| Ingredients Starter (g/kg) | ||
| Maize | 329.5 | |
| Barley | 250 | |
| Wheat | 170 | |
| Soybean | 129 | |
| Soybean meal | 61 | |
| Vitamins and oligoelements premix | 25 | |
| Soy husk | 10 | |
| Liquid lysine | 7.5 | |
| Monocalcic phosphate | 5.5 | |
| Lard | 5 | |
| Salt | 4 | |
| Calcium carbonate | 3.5 | |
| Vitamins | ||
| Vitamin A UI/kg | 6500 | 6500 |
| Vitamin D3 UI/kg | 1500 | 1500 |
| Vitamin E UI/kg | 92 | 92 |
| Coline chloride mg/kg | 1356 | 1356 |
| Oligoelements (mg/kg) | ||
| Fe | 120 | 120 |
| Mn | 30 | 30 |
| Zn | 110 | 110 |
| Se | 0.20 | 0.20 |
| I | 0.8 | 0.8 |
| Cu | 80 | 80 |
| Analytical Components (%) | ||
| Crude protein | 16.0 | 16.0 |
| Crude fibre | 2.7 | 4.0 |
| Crude fat | 3.8 | 3.0 |
| Crude ash | 6.3 | 4.3 |
| Calcium | 0.85 | 0.72 |
| Phosphorus | 0.67 | 0.50 |
| Sodium | 0.26 | 0.25 |
| Metionine | 0.49 | 0.44 |
| Lisine | 1.18 | 1.09 |
| Group | Mean (kg) | SEM (kg) |
|---|---|---|
| Control | 5.66 | 0.072 |
| ZnO | 5.59 | 0.070 |
| OEO 0.4% | 5.49 | 0.090 |
| OEO 1.2% | 5.87 | 0.061 |
| Garlic 0.4% | 5.68 | 0.082 |
| Garlic 2% | 5.67 | 0.174 |
| Group | ADG | FCR | Feed Intake | FBW | ||||
|---|---|---|---|---|---|---|---|---|
| Mean | SEM | Mean | SEM | Mean | SEM | Mean | SEM | |
| Control | 0.265 a | 0.025 | 2.120 | 0.187 | 26.763 | 1.839 | 18.645 a | 1.251 |
| ZnO | 0.263 a | 0.014 | 1.811 | 0.091 | 23.150 | 0.992 | 18.493 a | 0.734 |
| OEO 0.4% | 0.351 c | 0.007 | 1.456 | 0.089 | 24.974 | 1.373 | 22.701 b | 0.408 |
| OEO 1.2% | 0.307 abc | 0.007 | 1.787 | 0.064 | 26.903 | 0.877 | 20.937 ab | 0.368 |
| Garlic 0.4% | 0.342 bc | 0.011 | 1.876 | 0.217 | 31.338 | 3.442 | 22.523 b | 0.555 |
| Garlic 2% | 0.285 ab | 0.009 | 1.985 | 0.074 | 27.710 | 1.275 | 19.635 ab | 0.469 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rivera-Gomis, J.; Peres Rubio, C.; Martínez Conesa, C.; Otal Salaverri, J.; Cerón, J.J.; Tortosa, D.E.; Cubero Pablo, M.J. Effects of Dietary Supplementation of Garlic and Oregano Essential Oil on Biomarkers of Oxidative Status, Stress and Inflammation in Postweaning Piglets. Animals 2020, 10, 2093. https://doi.org/10.3390/ani10112093
Rivera-Gomis J, Peres Rubio C, Martínez Conesa C, Otal Salaverri J, Cerón JJ, Tortosa DE, Cubero Pablo MJ. Effects of Dietary Supplementation of Garlic and Oregano Essential Oil on Biomarkers of Oxidative Status, Stress and Inflammation in Postweaning Piglets. Animals. 2020; 10(11):2093. https://doi.org/10.3390/ani10112093
Chicago/Turabian StyleRivera-Gomis, Jorge, Camila Peres Rubio, Cristina Martínez Conesa, Julio Otal Salaverri, José Joaquín Cerón, Damián Escribano Tortosa, and María José Cubero Pablo. 2020. "Effects of Dietary Supplementation of Garlic and Oregano Essential Oil on Biomarkers of Oxidative Status, Stress and Inflammation in Postweaning Piglets" Animals 10, no. 11: 2093. https://doi.org/10.3390/ani10112093