Unrestricted Feed Intake Induces β-Cell Death and Impairs Insulin Secretion in Broiler Breeder Hens

 , and
, and 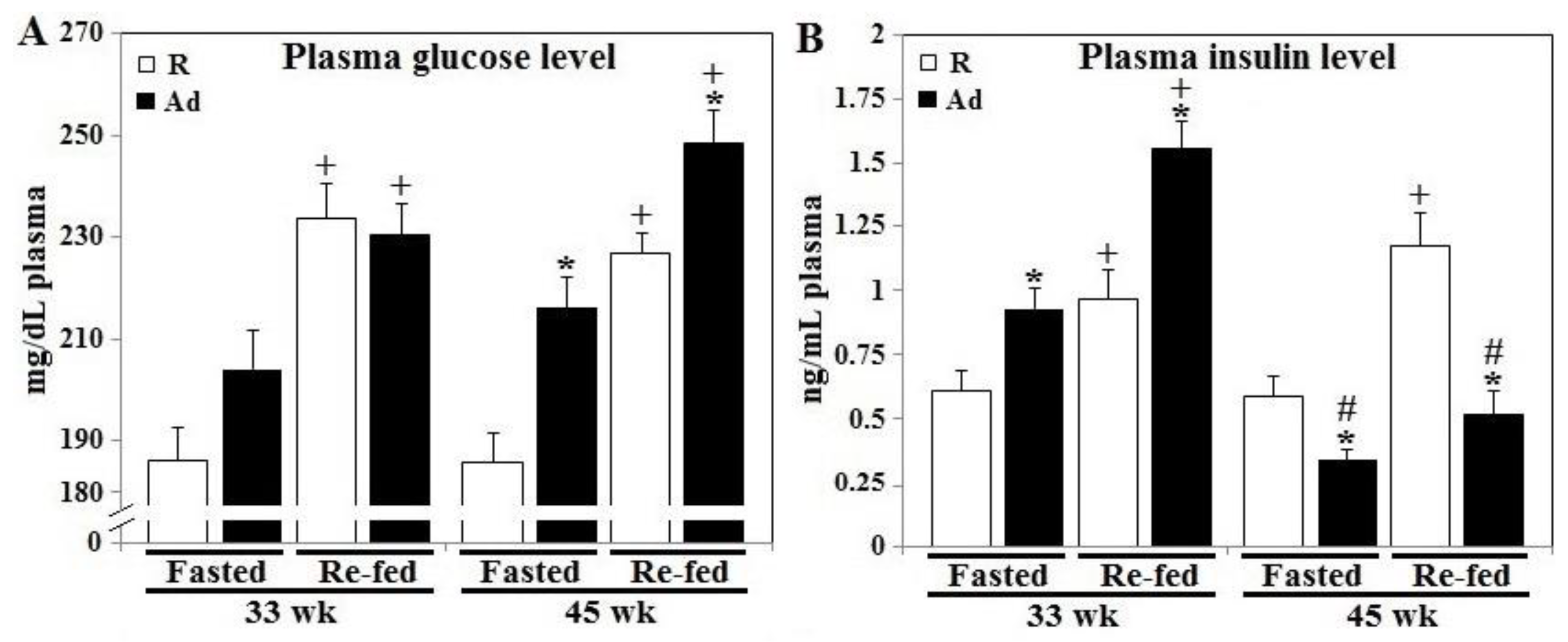{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Management
2.2. Necropsy and Tissue Collection
2.3. Prandial Changes of Plasma Insulin and Glucose Levels
2.4. Glucose Tolerance Test
2.5. Isolation of Pancreatic β-Islets
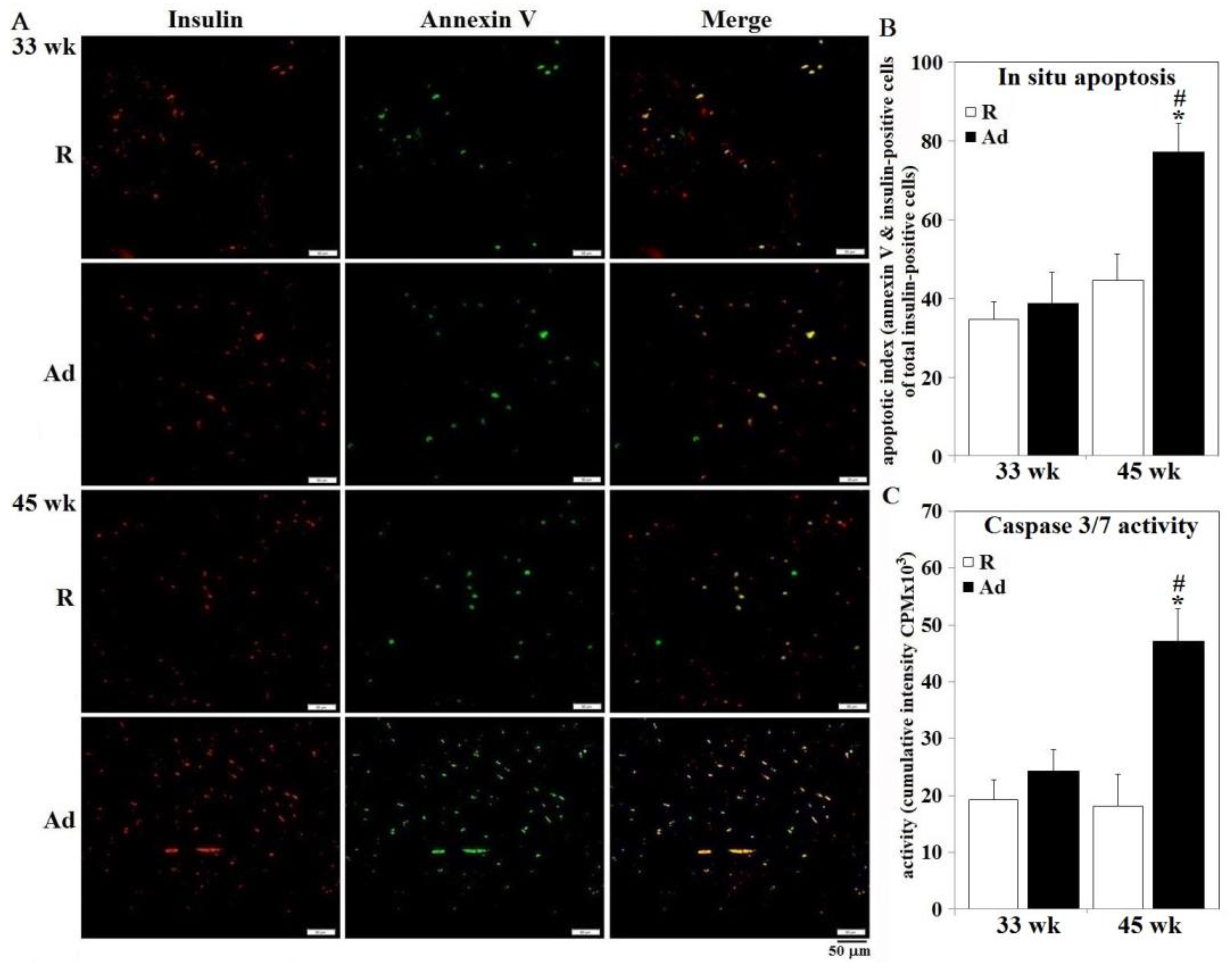
2.6. Immunohistochemistry and Apoptosis Analysis
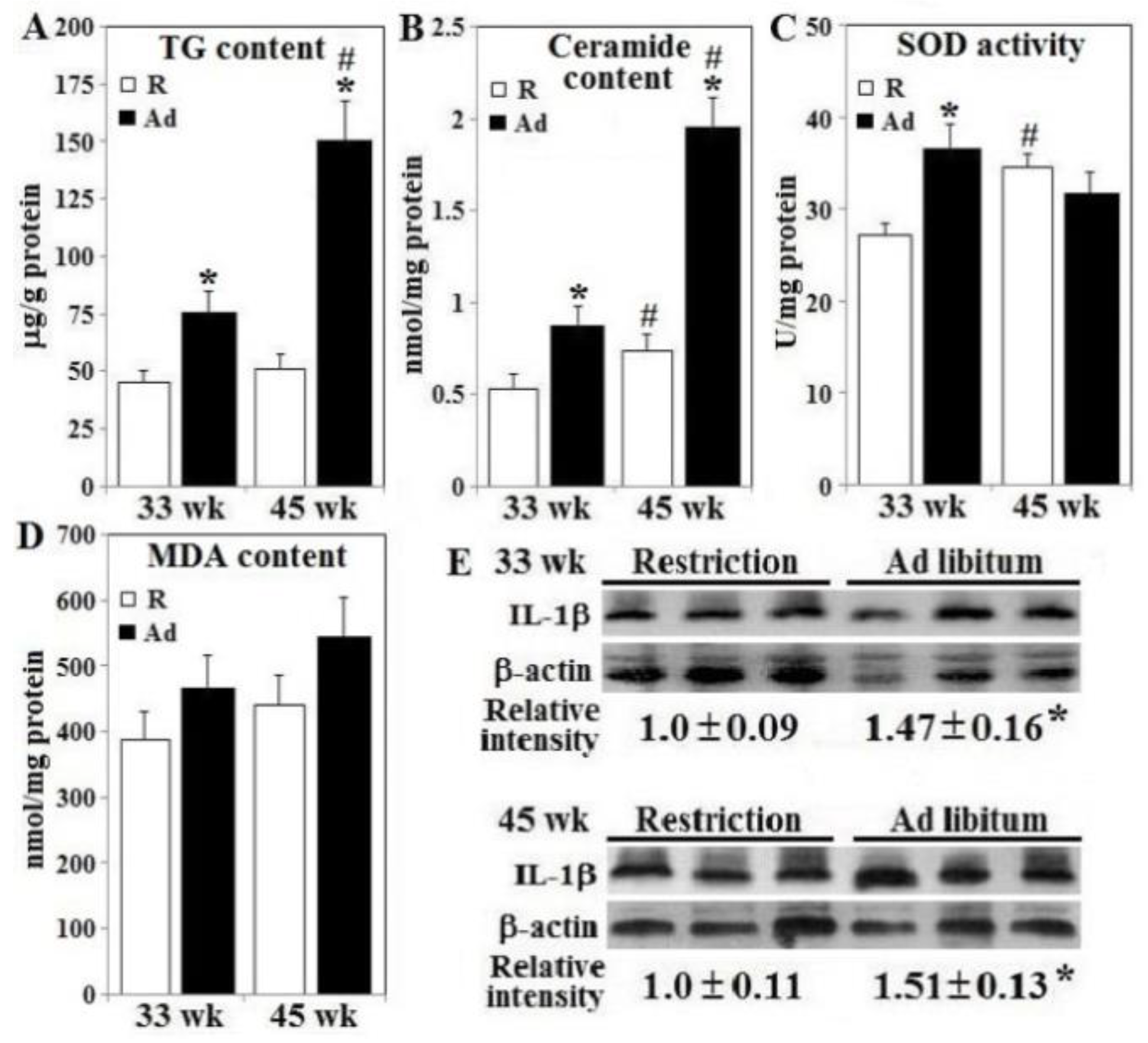
2.7. Pancreatic β-Islet TG, Ceramide, MDA Content, SOD and PDH Activity Analysis
2.8. Gene Expression by qRT-PCR Analysis
2.9. Western Blot Analysis
2.10. Statistics
3. Results
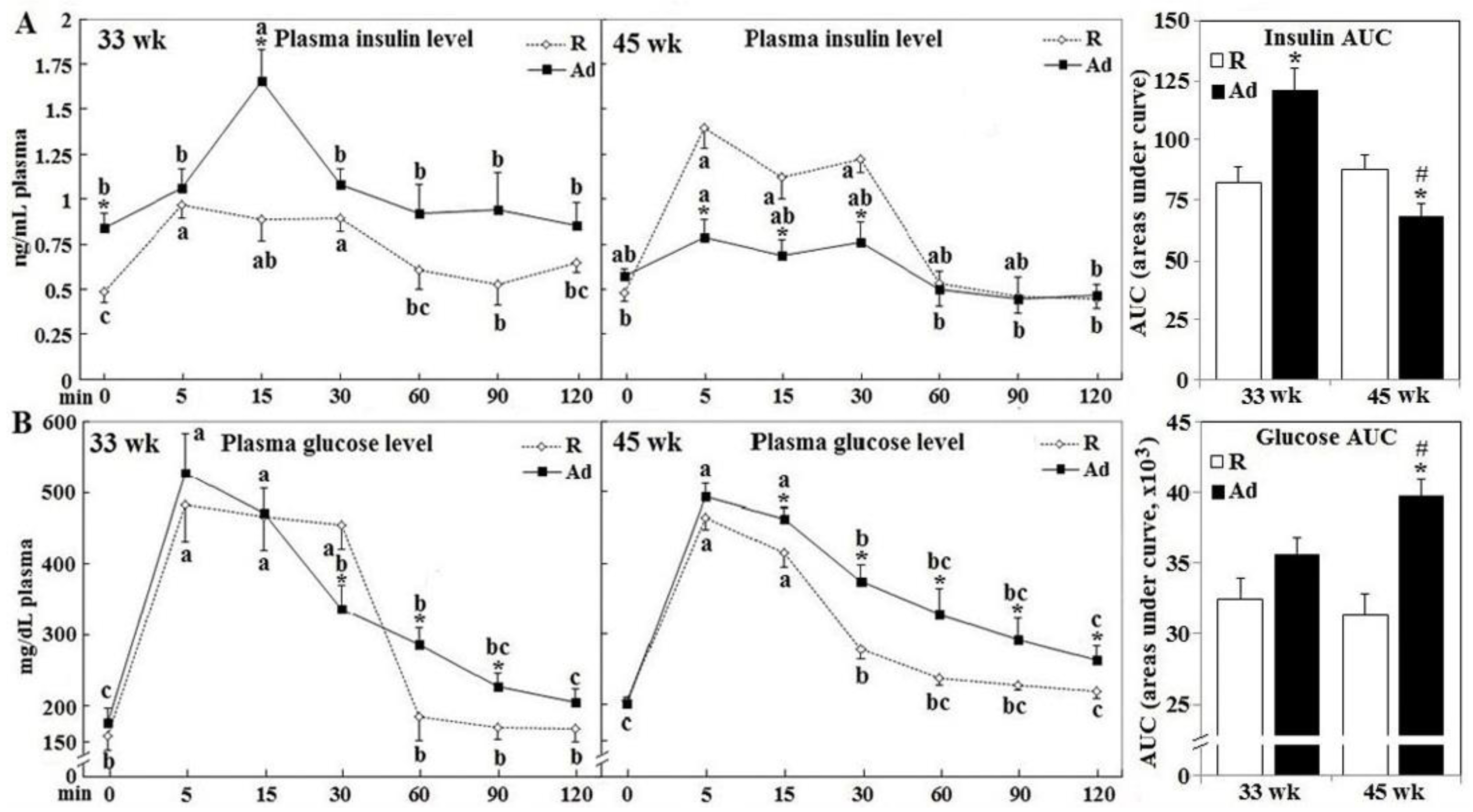
3.1. Prandial Changes of Plasma Glucose and Insulin Levels
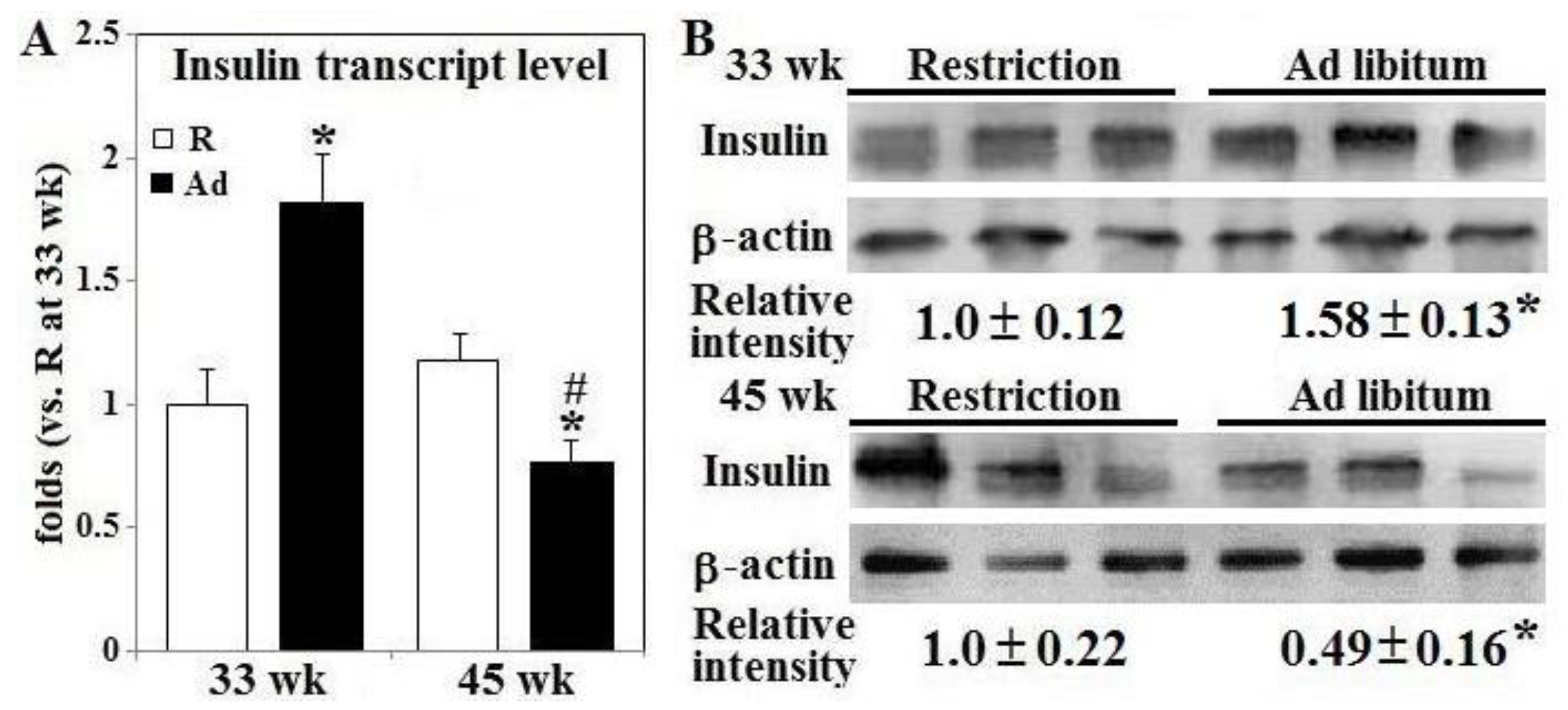
3.2. Glucose Disposal, Insulin Secretion, and Gene Expression
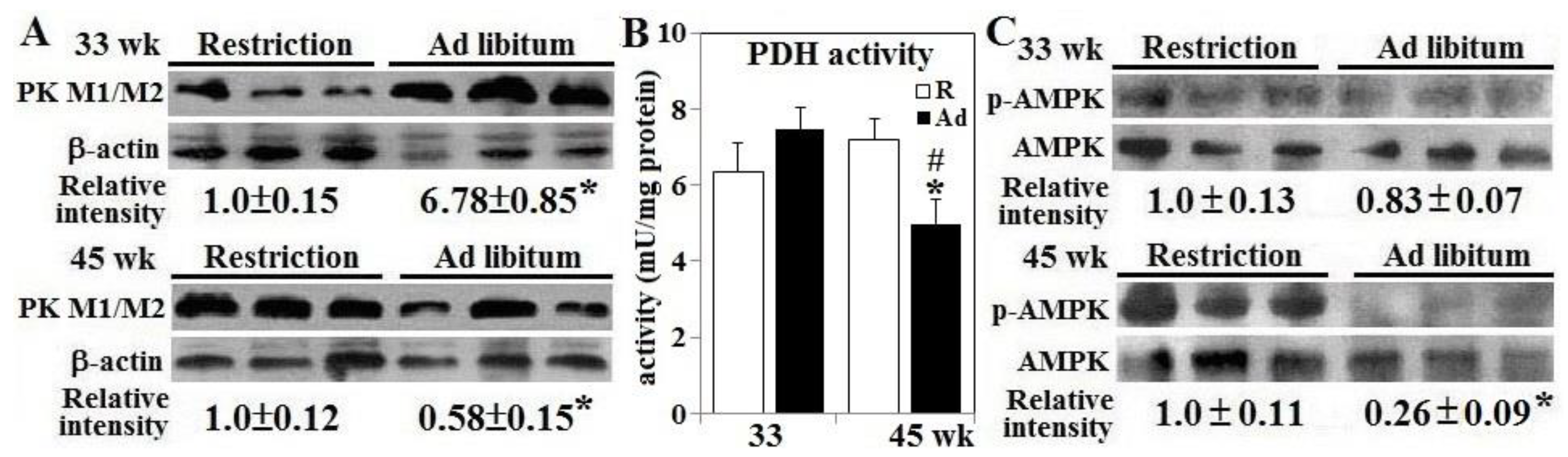
3.3. AMPK Activation and Pyruvate Metabolism
3.4. Lipotoxicity, Oxidative Stress, Inflammation Status, and β-Cell Apoptosis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Griffin, H.D.; Goddard, C. Rapidly growing broiler (meat-type) chickens: Their origin and use for comparative studies of the regulation of growth. Int. J. Biochem. 1994, 26, 19–28. [Google Scholar] [CrossRef]
- Julian, R.J. Physiological, management and environmental triggers of the ascites syndrome: A review. Avian Pathol. 2000, 9, 519–527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.E.; McMurtry, J.P.; Walzem, R.L. Overfeeding-induced ovarian dysfunction in broiler breeder hens is associated with lipotoxicity. Poult. Sci. 2006, 85, 70–81. [Google Scholar] [CrossRef] [PubMed]
- Richards, M.P.; Proszkowiec-Weglarz, M. Mechanisms regulating feed intake, energy expenditure, and body weight in poultry. Poult. Sci. 2007, 86, 1478–1490. [Google Scholar] [CrossRef]
- Dupont, J.; Chen, J.; Derouet, M.; Simon, J.; Leclercq, B.; Taouis, M. Metabolic differences between genetically lean and fat chickens are partly attributed to the alteration of insulin signaling in liver. J. Nutr. 1999, 129, 1937–1944. [Google Scholar] [CrossRef] [Green Version]
- Dupont, J.; Tesseraud, S.; Derouet, M.; Collin, A.; Rideau, N.; Crochet, S.; Godet, E.; Cailleau-Audouin, E.; Métayer-Coustard, S.; Duclos, M.J.; et al. Insulin immuno-neutralization in chicken: Effects on insulin signaling and gene expression in liver and muscle. J. Endocrinol. 2008, 197, 531–542. [Google Scholar] [CrossRef] [Green Version]
- Dupont, J. Endocrine Pancreas. In Sturkie’s Avian Physiology Chapter 27-Endocrine Pancreas, 6th ed.; Scanes, C.G., Ed.; Academic Press: Cambridge, MA, USA, 2015; pp. 613–631. [Google Scholar]
- Shiraishi, J.; Yanagita, K.; Fukumori, R.; Sugino, T.; Fujita, M.; Kawakami, S.; McMurtry, J.P.; Bungo, T. Comparisons of insulin related parameters in commercial-type chicks: Evidence for insulin resistance in broiler chicks. Physiol. Behav. 2011, 103, 233–239. [Google Scholar] [CrossRef]
- Rideau, N. Insulin secretion. In Perspectives in Avian Endocrinology; Harvey, S., Etches, R.J., Eds.; Journal of Endocrinology Ltd.: Bristol, UK, 1997; pp. 329–334. [Google Scholar]
- Zawalich, W.S.; Yamazaki, H.; Zawalich, K.C. Biphasic insulin secretion from freshly isolated or cultured, perifused rodent islets: Comparative studies with rats and mice. Metabolism 2008, 57, 30–39. [Google Scholar] [CrossRef] [Green Version]
- King, D.L.; Hazelwood, R.L. Regulation of avian insulin secretion by isolated perfused chicken pancreas. Am. J. Physiol. 1976, 231, 1830–1839. [Google Scholar] [CrossRef] [Green Version]
- Lin, H.Y.; Chung, T.K.; Chen, Y.H.; Walzem, R.L.; Chen, S.E. Dietary supplementation of 25-hydroxycholecalciferol improves livability in broiler breeder hens. Poult. Sci. 2019, 98, 6108–6166. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.Y.; Chou, P.C.; Chen, Y.H.; Chung, T.K.; Lai, L.S.; Walzem, R.L.; Huang, S.Y.; Chen, S.E. Dietary supplementation of 25-hydroxycholecalciferol improves livability in broiler breeder hens-amelioration of cardiac pathogenesis and hepatopathology. Animals 2019, 9, 770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.Y.; Lin, H.Y.; Chen, Y.W.; Ko, Y.J.; Liu, Y.J.; Chen, Y.H.; Walzem, R.L.; Chen, S.E. Obesity-associated cardiac pathogenesis in broiler breeder hens: Pathological adaption of cardiac hypertrophy. Poult. Sci. 2017, 96, 2428–2437. [Google Scholar] [CrossRef]
- Chen, C.Y.; Huang, Y.F.; Ko, Y.J.; Liu, Y.J.; Chen, Y.H.; Walzem, R.L.; Chen, S.E. Obesity-associated cardiac pathogenesis in broiler breeder hens: Development of metabolic cardiomyopathy. Poult. Sci. 2017, 96, 2438–2446. [Google Scholar] [CrossRef]
- Datar, S.P.; Bhonde, R.R. Cryopreservation of chick islets. CryoLetters 2010, 31, 485–492. [Google Scholar]
- Franssens, L.; Lesuisse, J.; Wang, Y.; De Ketelaere, B.; Willems, E.; Koppenol, A.; Guo, X.; Buyse, J.; Decuypere, E.; Everaert, N. Prenatal tolbutamide treatment alters plasma glucose and insulin concentrations and negatively affects the postnatal performance of chickens. Domest. Anim. Endocrinol. 2015, 52, 35–42. [Google Scholar] [CrossRef]
- Datar, S.P.; Suryavanshi, D.S.; Bhonde, R.R. Chick pancreatic B islets as an alternative in vitro model for screening insulin secretagogues. Poult. Sci. 2006, 85, 2260–2264. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.C.; Xie, Y.L.; Chang, C.J.; Su, C.M.; Chen, Y.H.; Huang, S.Y.; Walzem, R.L.; Chen, S.E. Feed intake alters immune cell functions and ovarian infiltration in broiler hens-implications for reproductive performance. Biol. Reprod. 2014, 90, 134. [Google Scholar] [CrossRef] [Green Version]
- Alarcon, C.; Serna, J.; Perez-Villamil, B.; de Pablo, F. Synthesis and differentially regulated processing of proinsulin in developing chick pancreas, liver and neuroretina. FEBS Lett. 1998, 436, 361–3666. [Google Scholar] [CrossRef] [Green Version]
- Pan, Y.E.; Liu, Z.C.; Chang, C.J.; Xie, Y.L.; Chen, C.Y.; Chen, C.F.; Walzem, R.L.; Chen, S.E. Ceramide accumulation and upregulation of proinflammatory interleukin-1β exemplify lipotoxicity to mediate declines of reproductive efficacy of broiler hens. Domest. Anim. Endocrinol. 2012, 42, 183–194. [Google Scholar] [CrossRef]
- He, X.; Dagan, A.; Gatt, S.; Schuchman, E.H. Simultaneous quantitative analysis of ceramide and sphingosine in mouse blood by naphthalene-2, 3-dicarboxyaldehyde derivatization after hydrolysis with ceramidase. Anal. Biochem. 2005, 340, 113–122. [Google Scholar] [CrossRef]
- Poitout, V.; Hagman, D.; Stein, R.; Artner, I.; Robertson, R.P.; Harmon, J.S. Regulation of the insulin gene by glucose and fatty acids. J. Nutr. 2006, 136, 873–876. [Google Scholar] [CrossRef] [PubMed]
- Randle, P.J.; Priestman, D.A.; Mistry, S.; Halsall, A. Mechanisms modifying glucose oxidation in diabetes mellitus. Diabetologia 1994, 37, S155–S161. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.Q.; Tornheim, K.; Leahy, J.L. Glucose-fatty acid cycle to inhibit glucose utilization and oxidation is not operative in fatty acid-cultured islets. Diabetes 1999, 48, 1747–1753. [Google Scholar] [CrossRef]
- Liu, Y.Q.; Moibi, J.A.; Leahy, J.L. Chronic high glucose lowers pyruvate dehydrogenase activity in islets through enhanced production of long chain acyl-CoA: Prevention of impaired glucose oxidation by enhanced pyruvate recycling through the malate-pyruvate shuttle. J. Biol. Chem. 2004, 279, 7470–7475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leclercq, B.; Whitehead, C.C. Leanness in Domestic Birds, 1st ed.; Butterworth & Co Ltd-INRA: London, UK, 1998.
- Dupont, J.; Dagou, C.; Derouet, M.; Simon, J.; Taouis, M. Early steps of insulin receptor signaling in chicken and rat: Apparent refractoriness in chicken muscle. Domest. Anim. Endocrinol. 2004, 26, 127–142. [Google Scholar] [CrossRef]
- Dupont, J.; Metayer-Coustard, S.; Ji, B.; Rame, C.; Gespach, C.; Voy, B.; Simon, J. Characterization of major elements of insulin signaling cascade in chicken adipose tissue: Apparent insulin refractoriness. Gen. Comp. Endocrinol. 2012, 176, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Proszkowiec-Weglarz, M.; Dupont, J.; Rideau, N.; Gespach, C.; Simon, J.; Porter, T.E. Insulin immuno-neutralization decreases food intake in chickens without altering hypothalamic transcripts involved in food intake and metabolism. Poult. Sci. 2017, 96, 4409–4418. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, Y.-F.; Chang, L.-C.; Chen, C.-Y.; Chen, Y.-H.; Walzem, R.L.; Chen, S.-E. Unrestricted Feed Intake Induces β-Cell Death and Impairs Insulin Secretion in Broiler Breeder Hens. Animals 2020, 10, 1969. https://doi.org/10.3390/ani10111969
Huang Y-F, Chang L-C, Chen C-Y, Chen Y-H, Walzem RL, Chen S-E. Unrestricted Feed Intake Induces β-Cell Death and Impairs Insulin Secretion in Broiler Breeder Hens. Animals. 2020; 10(11):1969. https://doi.org/10.3390/ani10111969
Chicago/Turabian StyleHuang, Yu-Feng, Ling-Chu Chang, Chung-Yu Chen, Yu-Hui Chen, Rosemary L. Walzem, and Shuen-Ei Chen. 2020. "Unrestricted Feed Intake Induces β-Cell Death and Impairs Insulin Secretion in Broiler Breeder Hens" Animals 10, no. 11: 1969. https://doi.org/10.3390/ani10111969