Novel InDels of GHR, GHRH, GHRHR and Their Association with Growth Traits in Seven Chinese Sheep Breeds

Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. DNA Samples and Data Collection
2.2. DNA Isolation and Genomic DNA Pools Construction
2.3. Primer Design and PCR Amplification
2.4. Statistical Analyses
3. Results
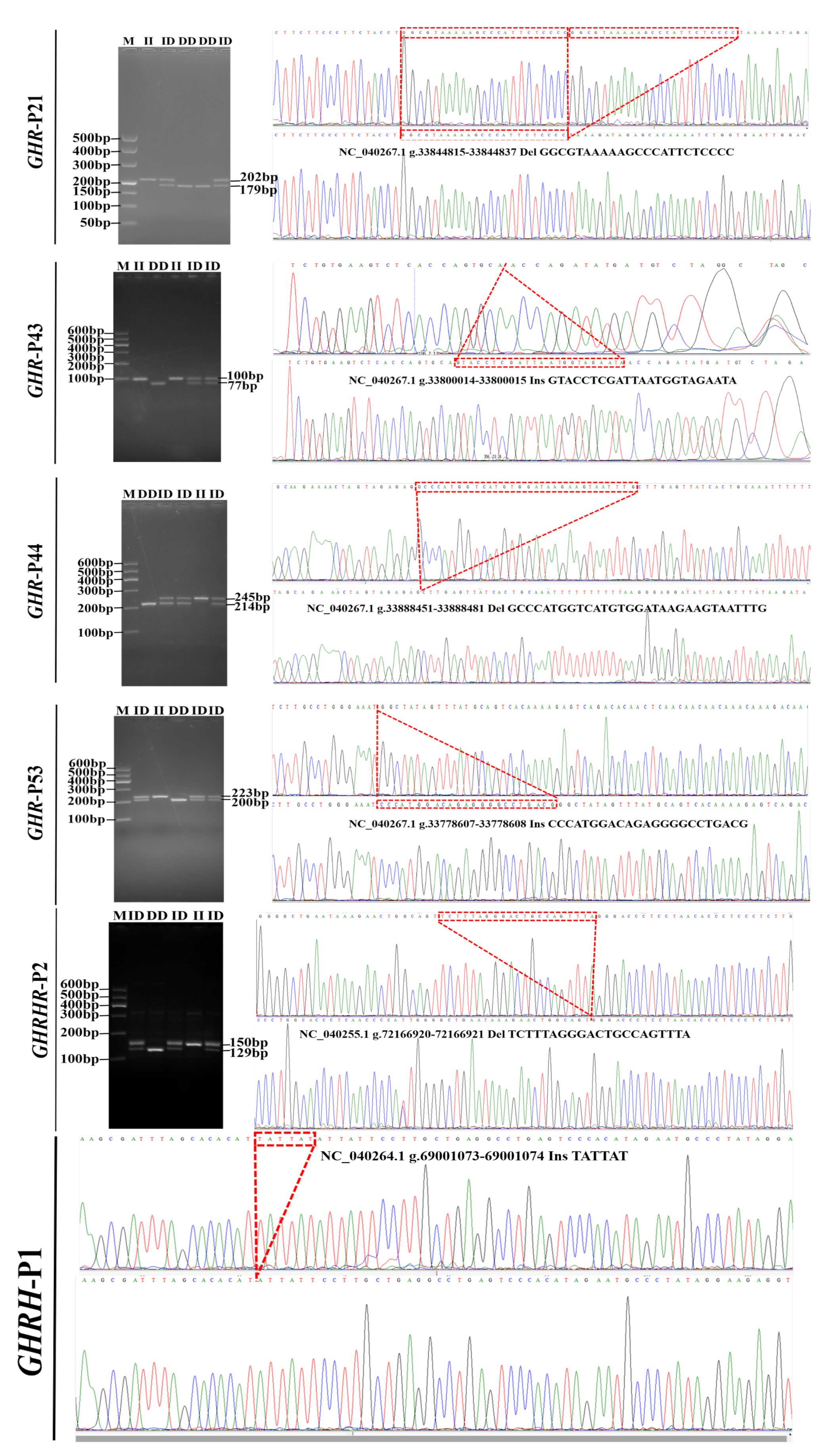
3.1. InDel Identification and Distribution
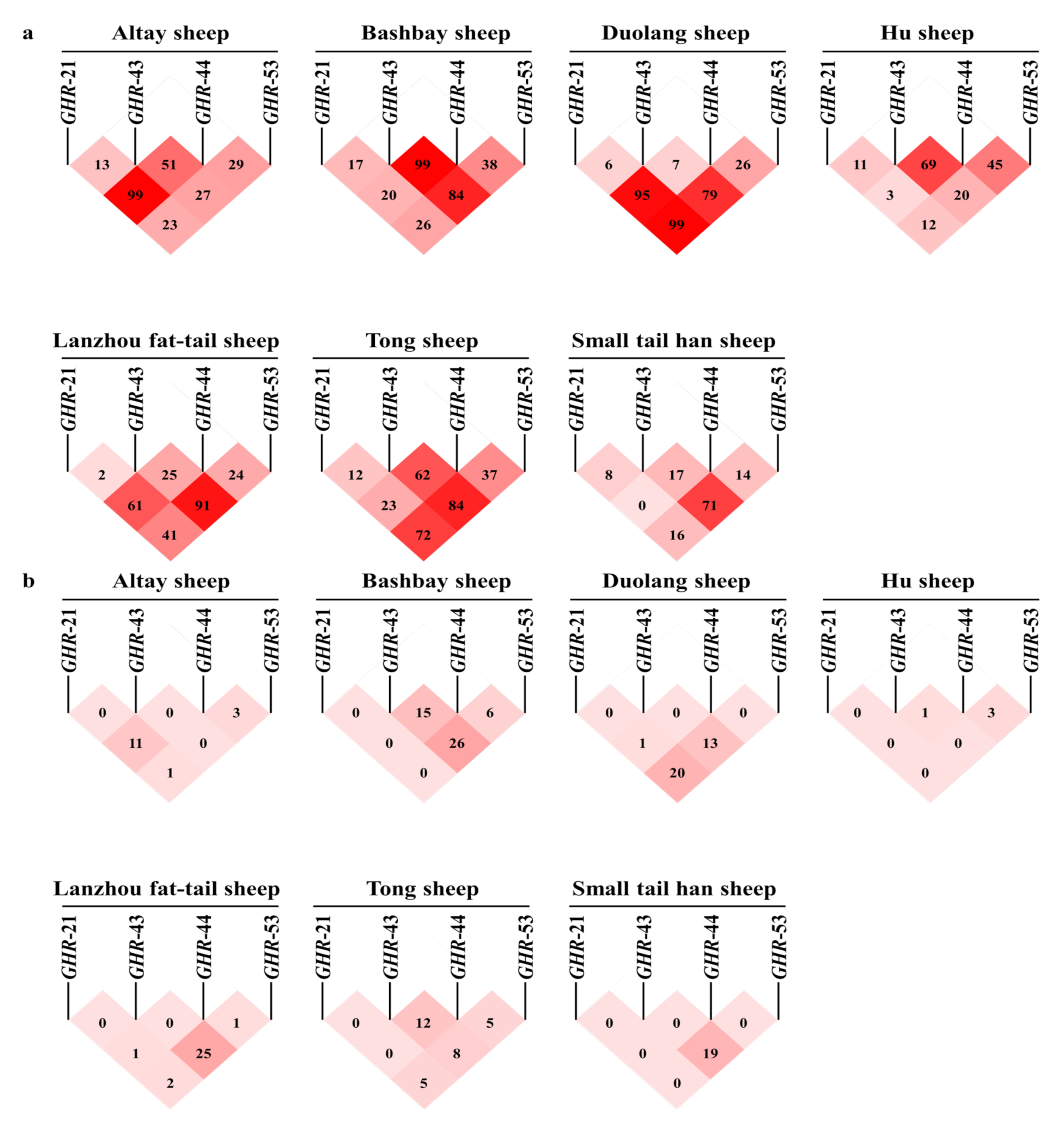
3.2. Population Genetic Analysis of Six Mutation Sites in Seven Varieties
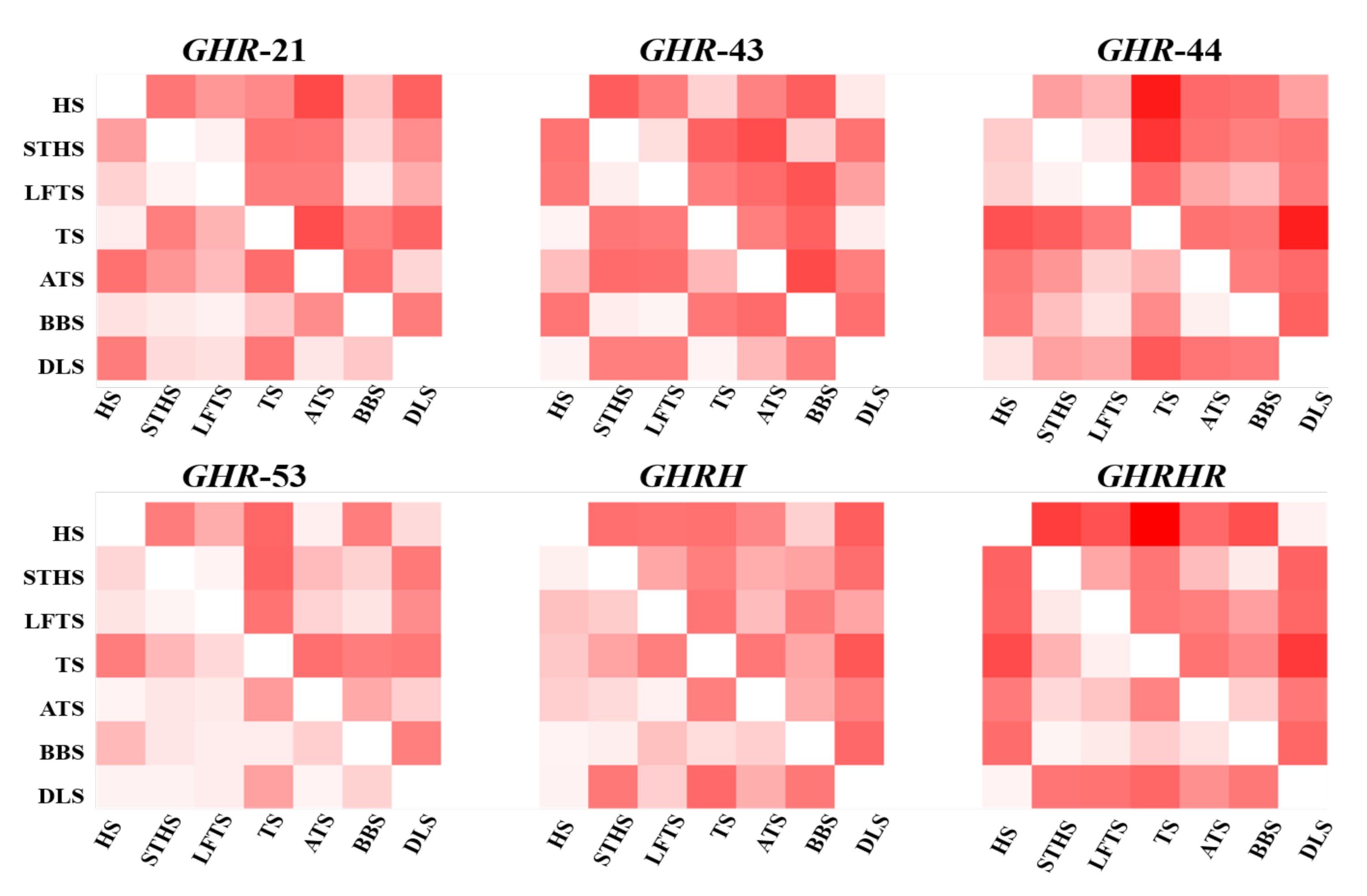
3.3. InDel Association Analyses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Yang, J.; He, J.; Wang, D.; Shi, E.; Yang, W.; Geng, Q.; Wang, Z. Progress in research and application of InDel markers. Sheng Wu Duo Yang Xing 2016, 24, 237–243. [Google Scholar] [CrossRef] [Green Version]
- Hyten, D.; Cannon, S.; Song, Q.; Weeks, N.; Fickus, E.; Shoemaker, R.; Specht, J.; Farmer, A.; May, G.; Cregan, P. High-throughput SNP discovery through deep rese-quencing of a reduced representation library to anchor and orient scaffolds in the soybean whole genome sequence. BMC Genom. 2010, 11, 38. [Google Scholar] [CrossRef]
- Fan, Y.; Wang, W.; Ma, G.; Liang, L.; Shi, Q.; Tao, S. Patterns of insertion and deletion in mammalian genomes. Curr. Genom. 2007, 8, 370–378. [Google Scholar]
- Alkan, C.; Coe, B.; Eichler, E. Genome structural variation discovery and genotyping. Nat. Rev. Genet. 2011, 12, 363–376. [Google Scholar] [CrossRef]
- Tuzun, E.; Sharp, A.; Bailey, J.; Kaul, R.; Morrison, V.; Pertz, L.; Haugen, E.; Hayden, H.; Albertson, D.; Pinkel, D.; et al. Fine-scale structural variation of the human genome. Nat. Genet. 2005, 37, 727–732. [Google Scholar] [CrossRef]
- Zhang, K. Function and Application of Intron. China Anim. Husb. Vet. Med. 2012, 39, 80–83. [Google Scholar]
- Lv, H.; Yang, L.; Kang, J.; Wang, Q.; Wang, X.; Fang, Z.; Liu, Y.; Zhuang, M.; Zhang, Y.; Lin, Y.; et al. Development of InDel markers linked to Fusarium wilt resistance in cabbage. Mol. Breed. 2013, 32, 961–967. [Google Scholar] [CrossRef]
- Vasemägi, A.; Gross, R.; Palm, D.; Paaver, T.; Primmer, C. Discovery and application of insertion-deletion (INDEL) polymorphisms for QTL mapping of early life-history traits in Atlantic salmon. BMC Genom. 2010, 11, 156. [Google Scholar] [CrossRef] [Green Version]
- Santos, N.; Ribeiro-Rodrigues, E.; Ribeiro-Dos-Santos, A.; Pereira, R.; Gusmão, L.; Amorim, A.; Guerreiro, J.; Zago, M.; Matte, C.; Hutz, M.; et al. Assessing individual interethnic admixture and population substructure using a 48-insertion-deletion (INSERTION) ancestry-informative marker (AIM) panel. Hum. Mutat. 2010, 31, 184–190. [Google Scholar] [CrossRef]
- Pereira, R.; Phillips, C.; Alves, C.; Amorim, A.; Carracedo, A.; Gusmão, L. A new multiplex for human identification using insertion/deletion polymorphisms. Electrophoresis 2009, 30, 3682–3690. [Google Scholar] [CrossRef]
- Lee, T.; Chafets, D.; Reed, W.; Wen, L.; Yang, Y.; Chen, J.; Utter, G.; Owings, J.; Busch, M. Enhanced ascertainment of microchimerism with real-time quantitative polymerase chain reaction amplification of insertion-deletion polymor-phisms. Transfusion 2006, 46, 1870–1878. [Google Scholar] [CrossRef]
- Wu, M.; Li, S.; Zhang, G.; Fan, Y.; Gao, Y.; Huang, Y.; Lan, X.; Lei, C.; Ma, Y.; Dang, R. Exploring insertions and deletions (indels) of MSRB3 gene and their association with growth traits in four Chinese indigenous cattle breeds. Arch. Anim. Breed. 2019, 62, 465–475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, J.; Xu, J.; Wang, J.; Wen, Y.; Niu, H.; Zheng, L.; He, H.; Peng, K.; He, P.; Shi, S.; et al. A novel SNP of PLAG1 gene and its association with growth traits in Chinese cattle. Gene 2019, 689, 166–171. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Huang, B.; Lai, Z.; Li, S.; Wu, F.; Qu, K.; Jia, Y.; Hou, J.; Liu, J.; Lei, C.; et al. The Distribution Characteristics of a 19-bp Indel of the PLAG1 Gene in Chinese Cattle. Animals 2019, 9, 1082. [Google Scholar] [CrossRef] [Green Version]
- Akhatayeva, Z.; Li, H.; Mao, C.; Cheng, H.; Zhang, G.; Jiang, F.; Meng, X.; Yao, Y.; Lan, X.; Song, E.; et al. Detecting novel Indel variants within the GHR gene and their associations with growth traits in Luxi Blackhead sheep. Anim. Biotechnol. 2020, 31, 1–9. [Google Scholar] [CrossRef]
- Li, J.; Erdenee, S.; Zhang, S.; Wei, Z.; Zhang, M.; Jin, Y.; Wu, H.; Chen, H.; Sun, X.; Xu, H.; et al. Genetic effects of PRNP gene insertion/deletion (indel) on phenotypic traits in sheep. Prion 2018, 12, 42–53. [Google Scholar] [CrossRef] [Green Version]
- Hernández, L.; Lee, P.; Camacho-Hübner, C. Isolated growth hormone deficiency. Pituitary 2007, 10, 351–357. [Google Scholar] [CrossRef] [Green Version]
- Carakushansky, M.; Whatmore, A.J.; Clayton, P.E.; Shalet, S.; Gleeson, H.; Price, D.; Levine, M.; Salvatori, R. A new missense mutation in the growth hormone-releasing hormone receptor gene in familial isolated GH deficiency. Eur. J. Endocrinol. 2003, 148, 25–30. [Google Scholar] [CrossRef] [Green Version]
- Gorlov, I.; Kolosov, Y.; Shirokova, N.; Getmantseva, L.; Slozhenkina, M.; Mosolova, N.; Bakoev, N.; Leonova, M.; Kolosov, A.; Zlobina, E. Association of the growth hormone gene polymorphism with growth traits in Salsk sheep breed. Small Rumin. Res. 2017, 150, 11–14. [Google Scholar] [CrossRef]
- Armstrong, E.; Ciappesoni, G.; Iriarte, W.; Da Silva, C.; Macedo, F.; Navajas, E.A.; Brito, G.; San Julián, R.; Gimeno, D.; Postiglioni, A. Novel genetic polymorphisms associated with carcass traits in grazing Texel sheep. Meat Sci. 2018, 145, 202–208. [Google Scholar] [CrossRef]
- Jia, J.; Zhang, L.; Wu, J.; Ha, Z.; Li, W. Study of the correlation between GH gene polymorphism and growth traits in sheep. Genet. Mol. Res. 2014, 13, 7190–7200. [Google Scholar] [CrossRef]
- Valeh, M.; Tahmoorespour, M.; Ansari, M.; Nassiry, M.R. Association of growth traits with sscp polymorphisms at the growth hormone receptor (ghr) and growth hormone releasing hormone receptor (ghrhr) genes in the baluchi sheep. J. Anim. Vet. Adv. 2012, 8, 1063–1069. [Google Scholar]
- Ferraz, A.; Bortolossi, J.; Curi, R.; Ferro, M.; Ferro, J.; Furlan, L. Identification and characterization of polymorphisms within the 5’ flanking region, first exon and part of first intron of bovine GH gene. J. Anim. Breed. Genet. 2006, 123, 208–212. [Google Scholar] [CrossRef]
- Szmatoła, T.; Gurgul, A.; Jasielczuk, I.; Ząbek, T.; Ropka-Molik, K.; Litwińczuk, Z.; Bugno-Poniewierska, M. A Comprehensive Analysis of Runs of Homozygosity of Eleven Cattle Breeds Representing Different Production Types. Animals 2019, 9, 1024. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Wu, P.; Xuan, C.; Liu, Y.; Wu, J. Advances in body size measurement and conformation appraisal for sheep. Trans. Chin. Soc. Agric. Eng. 2016, 32, 190–197. [Google Scholar]
- Zhao, H.; He, S.; Wang, S.; Zhu, Y.; Xu, H.; Luo, R.; Lan, X.; Cai, Y.; Sun, X. Two New Insertion/Deletion Variants of the PITX2 Gene and their Effects on Growth Traits in Sheep. Anim. Biotechnol. 2018, 29, 276–282. [Google Scholar] [CrossRef]
- Gao, Y.; Huang, B.; Bai, F.; Wu, F.; Zhou, Z.; Lai, Z.; Li, S.; Qu, K.; Jia, Y.; Lei, C.; et al. Two Novel SNPs in RET Gene Are Associated with Cattle Body Measurement Traits. Animals 2019, 9, 836. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; He, S.; Zhu, Y.; Cao, X.; Luo, R.; Cai, Y.; Xu, H.; Sun, X. A novel 29 bp insertion/deletion (indel) variant of the LHX3 gene and its influence on growth traits in four sheep breeds of various fecundity. Arch. Anim. Breed. 2017, 60, 79–85. [Google Scholar] [CrossRef] [Green Version]
- He, L.; Bi, Y.; Wang, R.; Pan, C.; Chen, H.; Lan, X.; Qu, L. Detection of a 4 bp Mutation in the 3’UTR Region of Goat Sox9 Gene and Its Effect on the Growth Traits. Animals 2020, 10, 672. [Google Scholar] [CrossRef] [Green Version]
- Nei, M. Analysis of gene diversity in subdivided populations. Proc. Natl. Acad. Sci. USA 1973, 70, 3321–3323. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Li, B.; Shi, T.; Huang, Y.; Liu, G.; Lan, X.; Lei, C.; Chen, H. Copy number variation of bovine SHH gene is associated with body conformation traits in Chinese beef cattle. J. Appl. Genet. 2019, 60, 199–207. [Google Scholar] [CrossRef]
- Shi, T.; Peng, W.; Yan, J.; Cai, H.; Lan, X.; Lei, C.; Bai, Y.; Chen, H. A novel 17 bp indel in the SMAD3 gene alters transcription level, contributing to phenotypic traits in Chinese cattle. Arch. Anim. Breed. 2016, 59, 151–157. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Liu, S.; Yuan, T.; Sun, X. Genetic effects of FTO gene insertion/deletion (InDel) on fat-tail measurements and growth traits in Tong sheep. Anim. Biotechnol. 2019, 30, 1–11. [Google Scholar] [CrossRef]
- Bi, Y.; Feng, B.; Wang, Z.; Bi, Y.; Feng, B.; Wang, Z.; Zhu, H.; Qu, L.; Lan, X.; Pan, C.; et al. Myostatin (MSTN) Gene Indel Variation and Its Associations with Body Traits in Shaanbei White Cashmere Goat. Animals 2020, 10, 168. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Wang, K.; Liu, J.; Zhu, H.; Qu, L.; Chen, H.; Lan, X.; Pan, C.; Song, X. An 11-bp Indel Polymorphism within the CSN1S1 Gene Is Associated with Milk Performance and Body Measurement Traits in Chinese Goats. Animals 2019, 9, 1114. [Google Scholar] [CrossRef] [Green Version]
- El-Magd, M.; Abbas, H.; El-kattawy, A.; Mokhbatly, A. Novel polymorphisms of the IGF1R gene and their association with average daily gain in Egyptian buffalo (Bubalus bubalis). Domest. Anim. Endocrinol. 2013, 45, 105–110. [Google Scholar] [CrossRef]
- Mullen, M.; Berry, D.; Howard, D.; Diskin, M.; Lynch, C.; Giblin, L.; Kenny, D.; Magee, D.; Meade, K.; Waters, S. Single Nucleotide Polymorphisms in the Insulin-Like Growth Factor 1 (IGF-1) Gene are Associated with Performance in Holstein-Friesian Dairy Cattle. Front. Genet. 2011, 2, 3. [Google Scholar] [CrossRef] [Green Version]
- Maj, A.; Oprządek, J.; Dymnicki, E.; Zwierzchowski, L. Association of the polymorphism in the 5’-noncoding region of the bovine growth hormone receptor gene with meat production traits in Polish Black-and-White cattle. Meat Sci. 2006, 72, 539–544. [Google Scholar] [CrossRef]
- Zhang, C.; Luo, H.; Chen, Y.; Zhang, G.; Jia, Z. Correlation between Polymorphisms of 5′-Flanking Region of GHR Gene and Body Weight of Nanjiang Huang Goats. Chin. J. Anim. Sci. 2007, 43, 4–7. [Google Scholar]
- Wang, Q.; Diao, Y.; Xu, Z.; Li, X.; Luo, X.; Xu, H.; Ouyang, P.; Liu, M.; Hu, Z.; Wang, Q.; et al. Identification of a novel splicing mutation in the growth hormone (GH)-releasing hormone receptor gene in a Chinese family with pituitary dwarfism. Mol. Cell. Endocrinol. 2009, 313, 50–56. [Google Scholar] [CrossRef]
- Dettori, M.; Pazzola, M.; Paschino, P.; Amills, M.; Vacca, G. Association between the GHR, GHRHR, and IGF1 gene polymorphisms and milk yield and quality traits in Sarda sheep. J. Dairy Sci. 2018, 101, 9978–9986. [Google Scholar] [CrossRef] [Green Version]
- Sahu, A.R.; Jeichitra, V.; Rajendran, R.; Raja, A. Polymorphism of growth hormone receptor (GHR) gene in Nilagiri sheep. Trop. Anim. Health Prod. 2017, 49, 281–285. [Google Scholar] [CrossRef]
- Greenwood, T.; Kelsoe, J. Promoter and intronic variants affect the transcriptional regulation of the human dopamine transporter gene. Genomics 2003, 82, 511–520. [Google Scholar] [CrossRef]
- Lu, S.; Cullen, B. Analysis of the stimulatory eddect of splicing on mRNA production and utilization in mammalian cells. RNA 2003, 9, 618–630. [Google Scholar] [CrossRef] [Green Version]
- Chang, K. Critical regulatory domains in intron 2 of a porcine sarcomericmyosin heavy chain gene. J. Muscle Res. Cell Motil. 2000, 21, 451–461. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Breed | Abbreviation | Sampling Location | n | Sample Form |
|---|---|---|---|---|
| Small tail han | STHS | Lanzhou city (Gansu province) | 184 | Musculature of ear margin |
| Tong | TS | Baishui county (Shanxi province) | 268 | Venous blood |
| Lanzhou fat-tail | LFTS | Lanzhou city (Gansu province) | 67 | Musculature of ear margin |
| Hu | HS | Huzhou city (Zhejiang province) | 166 | Venous blood |
| Duolang | DLS | Shihezi city (Xinjiang autonomous region) | 92 | Venous blood |
| Bashbay | BBS | Shihezi city (Xinjiang autonomous region) | 96 | Venous blood |
| Altay | ATS | Altay city (Xinjiang autonomous region) | 96 | Venous blood |
| Total | 969 |
| Gene(Gene ID) | Name | Primer Sequences (5′-3′) | Product Size (bp) | Notes |
|---|---|---|---|---|
| GHR (443333) | GHR-21 | F: CGGTCCAATTCACCAGAT | 202 − 23 | Upstream |
| R: AGAATCCCACGGACAAAG | ||||
| GHR-43 | F: CTGTGAAGTCTCACCAGTGC | 77 + 23 | Upstream | |
| R: GGATAGGCAGAATGCTAAAG | ||||
| GHR-44 | F: GAAATACCCTTGTGGACAGA | 245 − 31 | Upstream | |
| R: CTGGATTTATTGTTATTGTCTATTG | ||||
| GHR-53 | F: ATGTTAGGCAGCCAAAAGG | 200 + 23 | 1st intron | |
| R: GCCCAACCCAATGTTAATAGA | ||||
| GHRH (100101237) | GHRH | F: ACGACTGAAGCGATTTAGCAC | 159 + 6 | Promoter region |
| R: TCCATCTGTAAAATGGGCATG | ||||
| GHRHR (443511) | GHRHR-2 | F: AACCCCTGTCTCAGTTTCTCC | 129 + 21 | 12th intron |
| R: GATCTCAGTCCTCACCTCCAA |
| Locus | Breed | Size | Genotypic Frequency | Allelic Frequency | Population Parameters | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| n | II | ID | DD | I | D | Ho | He | Ne | PIC | ||
| GHR-21 | HS | 184 | 161 | 18 | 5 | 0.924 | 0.076 | 0.859 | 0.141 | 1.164 | 0.131 |
| STHS | 268 | 203 | 57 | 8 | 0.864 | 0.136 | 0.765 | 0.235 | 1.308 | 0.208 | |
| LFTS | 67 | 52 | 13 | 2 | 0.873 | 0.127 | 0.778 | 0.222 | 1.285 | 0.197 | |
| TS | 166 | 146 | 20 | 0 | 0.940 | 0.060 | 0.887 | 0.113 | 1.128 | 0.107 | |
| ATS | 92 | 53 | 36 | 3 | 0.772 | 0.228 | 0.648 | 0.352 | 1.544 | 0.290 | |
| BBS | 96 | 78 | 15 | 3 | 0.891 | 0.109 | 0.805 | 0.195 | 1.242 | 0.176 | |
| DLS | 96 | 63 | 31 | 2 | 0.818 | 0.182 | 0.702 | 0.298 | 1.425 | 0.254 | |
| GHR-43 | HS | 184 | 132 | 47 | 5 | 0.845 | 0.155 | 0.738 | 0.262 | 1.355 | 0.228 |
| STHS | 268 | 133 | 111 | 24 | 0.703 | 0.297 | 0.583 | 0.417 | 1.716 | 0.330 | |
| LFTS | 67 | 37 | 16 | 4 | 0.672 | 0.328 | 0.621 | 0.389 | 1.610 | 0.307 | |
| TS | 165 | 114 | 49 | 2 | 0.839 | 0.161 | 0.730 | 0.270 | 1.369 | 0.233 | |
| ATS | 92 | 76 | 16 | 0 | 0.913 | 0.087 | 0.841 | 0.159 | 1.189 | 0.146 | |
| BBS | 96 | 41 | 47 | 8 | 0.672 | 0.328 | 0.559 | 0.441 | 1.789 | 0.344 | |
| DLS | 96 | 66 | 28 | 2 | 0.833 | 0.167 | 0.722 | 0.278 | 1.385 | 0.239 | |
| GHR-44 | HS | 184 | 142 | 38 | 4 | 0.875 | 0.125 | 0.781 | 0.219 | 1.280 | 0.377 |
| STHS | 268 | 184 | 76 | 8 | 0.828 | 0.172 | 0.716 | 0.284 | 1.397 | 0.244 | |
| LFTS | 67 | 45 | 19 | 3 | 0.813 | 0.187 | 0.696 | 0.304 | 1.436 | 0.257 | |
| TS | 166 | 62 | 83 | 21 | 0.623 | 0.377 | 0.531 | 0.469 | 1.885 | 0.359 | |
| ATS | 92 | 54 | 26 | 12 | 0.728 | 0.272 | 0.604 | 0.396 | 1.655 | 0.317 | |
| BBS | 96 | 53 | 39 | 4 | 0.755 | 0.245 | 0.630 | 0.370 | 1.587 | 0.301 | |
| DLS | 96 | 82 | 11 | 3 | 0.911 | 0.089 | 0.839 | 0.161 | 1.192 | 0.148 | |
| GHR-53 | HS | 184 | 49 | 97 | 38 | 0.530 | 0.470 | 0.502 | 0.498 | 1.993 | 0.374 |
| STHS | 268 | 70 | 114 | 84 | 0.474 | 0.526 | 0.501 | 0.499 | 1.994 | 0.374 | |
| LFTS | 67 | 17 | 29 | 21 | 0.470 | 0.530 | 0.502 | 0.498 | 1.993 | 0.374 | |
| TS | 166 | 16 | 98 | 52 | 0.392 | 0.608 | 0.524 | 0.476 | 1.910 | 0.363 | |
| ATS | 92 | 25 | 46 | 21 | 0.522 | 0.478 | 0.501 | 0.499 | 1.996 | 0.375 | |
| BBS | 96 | 19 | 44 | 33 | 0.427 | 0.573 | 0.511 | 0.489 | 1.958 | 0.301 | |
| DLS | 96 | 21 | 57 | 18 | 0.516 | 0.484 | 0.500 | 0.500 | 1.998 | 0.374 | |
| GHRH | HS | 184 | 6 | 73 | 105 | 0.231 | 0.769 | 0.645 | 0.355 | 1.550 | 0.292 |
| STHS | 268 | 24 | 68 | 176 | 0.216 | 0.784 | 0.661 | 0.339 | 1.513 | 0.282 | |
| LFTS | 67 | 4 | 11 | 52 | 0.142 | 0.858 | 0.757 | 0.243 | 1.322 | 0.408 | |
| TS | 166 | 23 | 53 | 90 | 0.298 | 0.702 | 0.581 | 0.419 | 1.720 | 0.331 | |
| ATS | 92 | 3 | 24 | 65 | 0.163 | 0.837 | 0.727 | 0.273 | 1.375 | 0.236 | |
| BBS | 96 | 6 | 34 | 56 | 0.240 | 0.760 | 0.636 | 0.364 | 1.573 | 0.298 | |
| DLS | 96 | 1 | 13 | 82 | 0.078 | 0.922 | 0.856 | 0.144 | 1.168 | 0.134 | |
| GHRHR-2 | HS | 184 | 167 | 15 | 2 | 0.948 | 0.052 | 0.902 | 0.097 | 1.109 | 0.093 |
| STHS | 268 | 173 | 80 | 15 | 0.795 | 0.205 | 0.673 | 0.326 | 1.484 | 0.273 | |
| LFTS | 67 | 42 | 17 | 8 | 0.754 | 0.246 | 0.502 | 0.498 | 1.993 | 0.374 | |
| TS | 166 | 82 | 76 | 8 | 0.723 | 0.277 | 0.629 | 0.371 | 1.590 | 0.302 | |
| ATS | 92 | 66 | 24 | 2 | 0.848 | 0.152 | 0.742 | 0.258 | 1.348 | 0.225 | |
| BBS | 96 | 61 | 31 | 4 | 0.797 | 0.203 | 0.676 | 0.324 | 1.489 | 0.271 | |
| DLS | 96 | 86 | 9 | 1 | 0.943 | 0.057 | 0.892 | 0.108 | 1.121 | 0.102 | |
| Loci | Breeds | Growth Traits | Observed Genotypes (LSM a ± SE) | p Values | ||
|---|---|---|---|---|---|---|
| II | ID | DD | ||||
| GHR-21 | LFHS | Hip height (cm) | 77.92 a ± 1.27 (n = 25) | 71.50 b ± 2.07 (n = 7) | 84.50 a ± 3.50 (n = 2) | 0.019 |
| GHR-43 | STHS | Body height (cm) | 62.60 b ± 0.45 (n = 90) | 64.26 a ± 0.46 (n = 80) | 62.37 a,b ± 0.85 (n = 20) | 0.023 |
| STHS | Chest depth (cm) | 27.17 b ± 0.29 (n = 90) | 27.89 a,b ± 0.27 (n = 80) | 28.66 a ± 0.61 (n = 20) | 0.036 | |
| TS | Hip width (cm) | 13.88 b ± 0.27 (n = 48) | 15.10 a,b ± 0.30 (n = 27) | 16.50 a ± 0.50 (n = 2) | 0.005 | |
| GHR-44 | HS | Body weight (kg) | 32.73 a ± 0.36 (n = 141) | 31.03 b ± 0.83 (n = 38) | 27.18 b ± 1.94 (n = 4) | 0.009 |
| STHS | Body height (cm) | 62.82 b ± 0.36 (n = 129) | 64.00 a,b ± 0.62 (n = 54) | 66.14 a ± 1.30 (n = 7) | 0.044 | |
| TS | Head depth (cm) | 14.53 b ± 1.32 (n = 26) | 14.94 a ± 0.11 (n = 42) | 14.20 b ± 0.32 (n = 10) | 0.007 | |
| GHR-53 | HS | Cannon girth (cm) | 6.96 b ± 0.09 (n = 49) | 7.12 a,b ± 0.05 (n = 97) | 7.25 a ± 0.10 (n = 38) | 0.049 |
| STHS | Chest circumference (cm) | 72.20 a,b ± 0.78 (n = 52) | 71.00 b ± 0.61 (n = 83) | 73.60 a ± 0.91 (n = 55) | 0.043 | |
| TS | Forehead width (cm) | 12.00 b ± 0.27 (n = 5) | 12.86 a ± 0.11 (n = 47) | 12.90 a ± 0.15 (n = 25) | 0.049 | |
| GHRHR-2 | LFHS | Body weight (kg) | 47.27 b ± 2.96 (n = 23) | 58.95 a,b ± 4.88 (n = 8) | 62.87 a ± 3.68 (n = 3) | 0.050 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, M.; Zhao, H.; Tang, X.; Li, Q.; Yi, X.; Liu, S.; Sun, X. Novel InDels of GHR, GHRH, GHRHR and Their Association with Growth Traits in Seven Chinese Sheep Breeds. Animals 2020, 10, 1883. https://doi.org/10.3390/ani10101883
Wu M, Zhao H, Tang X, Li Q, Yi X, Liu S, Sun X. Novel InDels of GHR, GHRH, GHRHR and Their Association with Growth Traits in Seven Chinese Sheep Breeds. Animals. 2020; 10(10):1883. https://doi.org/10.3390/ani10101883
Chicago/Turabian StyleWu, Mingli, Haidong Zhao, Xiaoqin Tang, Qi Li, Xiaohua Yi, Shirong Liu, and Xiuzhu Sun. 2020. "Novel InDels of GHR, GHRH, GHRHR and Their Association with Growth Traits in Seven Chinese Sheep Breeds" Animals 10, no. 10: 1883. https://doi.org/10.3390/ani10101883