Effect of Parity on Reproductive Performance and Composition of Sow Colostrum during First 24 h Postpartum


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
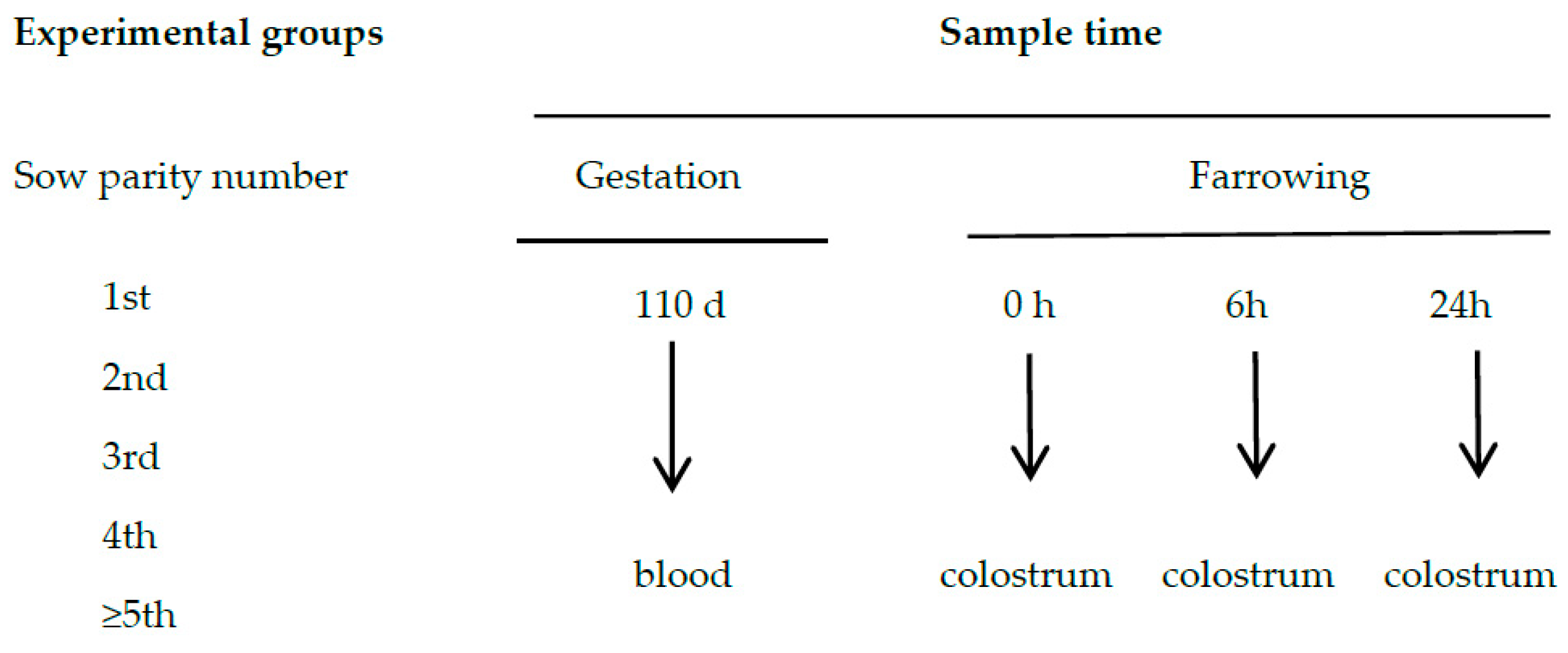
2.1. Animals, Measurements and Experimental Design
2.2. Samples Collection
2.3. Chemical Composition
2.4. Immunological Parameters
2.5. Data Analysis
3. Results
3.1. Reproductive Performance of Sow
3.2. Colostrum Chemical Composition
3.3. Immunological Parameters
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Hurley, W.L. Composition of Sow Colostrum. In The Gestating and Lactating Sow; Farmer, C., Ed.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2015; pp. 193–230. [Google Scholar]
- Theil, P.K.; Hurley, W.L. The Protein Component of Sow Colostrum and Milk. In Milk Proteins—From Structure to Biological Properties and Health Aspect; Gigli, I., Ed.; IntechOpen Publishe: London, UK, 2016. [Google Scholar] [CrossRef] [Green Version]
- Theil, P.K.; Lauridsen, C.; Quesnel, H. Neonatal piglet survival: Impact of sow nutrition around parturition on fetal glycogen deposition and production and composition of colostrum and transient milk. Animal 2014, 8, 1021–1030. [Google Scholar] [CrossRef] [PubMed]
- Frenyo, V.; Pethes, G.; Antal, T.; Szabo, I. Changes in colostral and serum IgG content in swine in relation to time. Vet. Res. Commun. 1980, 4, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Klobasa, F.; Werhahn, E.; Butler, J.E. Composition of sow milk during lactation. J. Anim. Sci. 1987, 64, 1458–1466. [Google Scholar] [CrossRef] [PubMed]
- Picone, G.; Zappaterra, M.; Luise, D.; Trimigno, A.; Capozzi, F.; Motta, V.; Davoli, R.; Nanni Costa, L.; Bosi, P.; Trevisi, P. Metabolomics characterization of colostrum in three sow breeds and its influences on piglets’ survival and litter growth rates. J. Anim. Sci. Biotechnol. 2018, 9, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nuntapaitoon, M.; Juthamanee, P.; Theil, P.K.; Tummaruk, P. Impact of sow parity on yield and composition of colostrum and milk in Danish Landrace × Yorkshire crossbred sows. Prev. Vet. Med. 2020, 181, 105085. [Google Scholar] [CrossRef] [PubMed]
- Nuntapaitoon, M.; Suwimonteerabutr, J.; Am-In, N.; Tienthai, P.; Chuesiri, P.; Kedkovid, R.; Tummaruk, P. Impact of parity and housing conditions on concentration of immunoglobulin G in sow colostrum. Trop. Anim. Health Prod. 2019, 51, 1239–1246. [Google Scholar] [CrossRef]
- Bourne, F.J.; Curtis, J. The transfer of immunoglobins IgG, IgA and IgM from serum to colostrum and milk in the sow. Immunology 1973, 24, 157–162. [Google Scholar]
- Quesnel, H. Colostrum production by sows: Variability of colostrum yield and immunoglobulin G concentrations. Animal 2011, 5, 1546–1553. [Google Scholar] [CrossRef] [Green Version]
- De Blas, C.; Gasa, J.; Mateos, G.G.; López-Bote, C.; Gorrachategui, M.; Aguilera, J.; Fructuoso, G. Necesidades Nutricionales Para el Ganado Porcino Normas FEDNA, 2nd ed.; Fundación Española para el Desarrollo de la Nutrición Animal: Madrid, Spain, 2013. [Google Scholar]
- Maes, D.G.D.; Janssens, G.P.J.; Delputte, P.; Lammertyn, A.; De Kruif, A. Back fat measurements in sows from three commercial pig herds: Relationship with reproductive efficiency and correlation with visual body condition scores. Livest. Prod. Sci. 2004, 91, 57–67. [Google Scholar] [CrossRef]
- Devillers, N.; Farmer, C.; Le Dividich, J.; Prunier, A. Variability of colostrum yield and colostrum intake in pigs. Animal 2007, 1, 1033–1041. [Google Scholar] [CrossRef] [Green Version]
- Arco-Pérez, A.; Ramos-Morales, E.; Yáñez-Ruiz, D.R.; Abecia, L.; Martín-García, A.I. Nutritive evaluation and milk quality of including of tomato or olive by-products silages with sunflower oil in the diet of dairy goats. Anim. Feed Sci. Technol. 2017, 232, 57–70. [Google Scholar] [CrossRef]
- ISO 8968-1/ IDF Standard 20-1 Part 1: Kjeldahl Principle and Crude Protein Calculation. In Milk and Milk Products: Determination of Nitrogen Content, 2nd ed.; International Organization for Standardization: Geneva, Switzerland, 2014.
- Marcelo, P.A.; Rizvi, S.S.H. Physicochemical properties of liquid virgin whey protein isolate. Int. Dairy J. 2008, 18, 236–246. [Google Scholar] [CrossRef]
- Le Dividich, J.; Rooke, J.A.; Herpin, P. Nutritional and immunological importance of colostrum for the new-born pig. J. Agric. Sci. 2005, 143, 469–485. [Google Scholar] [CrossRef] [Green Version]
- Quesnel, H.; Farmer, C.; Devillers, N. Colostrum intake: Influence on piglet performance and factors of variation. Livest. Sci. 2012, 146, 105–114. [Google Scholar] [CrossRef]
- Lavery, A.; Lawlor, P.G.; Magowan, E.; Miller, H.M.; O’driscoll, K.; Berry, D.P. An association analysis of sow parity, live-weight and back-fat depth as indicators of sow productivity. Animal 2019, 13, 622–630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milligan, B.N.; Fraser, D.; Kramer, D.L. Within-litter birth weight variation in the domestic pig and its relation to pre-weaning survival, weight gain, and variation in weaning weights. Livest. Prod. Sci. 2002, 76, 181–191. [Google Scholar] [CrossRef] [Green Version]
- Carney-Hinkle, E.E.; Tran, H.; Bundy, J.W.; Moreno, R.; Miller, P.S.; Burkey, T.E. Effect of dam parity on litter performance, transfer of passive immunity, and progeny microbial ecology. J. Anim. Sci. 2013, 91, 2885–2893. [Google Scholar] [CrossRef] [Green Version]
- Gama, L.L.; Johnson, R.K. Changes in ovulation rate, uterine capacity, uterine dimensions, and parity effects with selection for litter size in swine. J. Anim. Sci. 1993, 71, 608–617. [Google Scholar] [CrossRef] [Green Version]
- Szyndler-Nȩdza, M.; Rózycki, M.; Eckert, R.; Mucha, A.; Koska, M.; Szulc, T. Relationships between chemical composition of colostrum and milk and rearing performance of piglets during a 21-day lactation. Ann. Anim. Sci. 2013, 13, 771–781. [Google Scholar] [CrossRef] [Green Version]
- Csapó, J.; Martin, T.G.; Csapó-Kiss, Z.S.; Házas, Z. Protein, fats, vitamin and mineral concentrations in porcine colostrum and milk from parturition to 60 days. Int. Dairy J. 1996, 6, 881–902. [Google Scholar] [CrossRef]
- Beyer, M.; Jentsch, W.; Kuhla, S.; Wittenburg, H.; Kreienbring, F.; Scholze, H.; Rudolph, P.E.; Metges, C.C. Effects of dietary energy intake during gestation and lactation on milk yield and composition of first, second and fourth parity sows. Arch. Anim. Nutr. 2007, 61, 452–468. [Google Scholar] [CrossRef] [PubMed]
- Mahan, D.C.; Peters, J.C. Long-term effects of dietary organic and inorganic selenium sources and levels on reproducing sows and their progeny. J. Anim. Sci. 2004, 82, 1343–1358. [Google Scholar] [CrossRef] [PubMed]
- Luise, D.; Cardenia, V.; Zappaterra, M.; Motta, V.; Bosi, P.; Rodriguez-Estrada, M.T.; Trevisi, P. Evaluation of breed and parity order effects on the lipid composition of porcine colostrum. J. Agric. Food Chem. 2018, 66, 12911–12920. [Google Scholar] [CrossRef]
- Declerck, I.; Dewulf, J.; Piepers, S.; Decaluwé, R.; Maes, D. Sow and litter factors influencing colostrum yield and nutritional composition. J. Anim. Sci. 2015, 93, 1309–1317. [Google Scholar] [CrossRef] [Green Version]
- Craig, J.R.; Dunshea, F.R.; Cottrell, J.J.; Wijesiriwardana, U.A.; Pluske, J.R. Primiparous and multiparous sows have largely similar colostrum and milk composition profiles throughout lactation. Animals 2019, 9, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foisnet, A.; Farmer, C.; David, C.; Quesnel, H. Relationships between colostrum production by primiparous sows and sow physiology around parturition. J. Anim. Sci. 2010, 88, 1672–1683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Theil, P.K.; Flummer, C.; Hurley, W.L.; Kristensen, N.B.; Labouriau, R.L.; Sørensen, M.T. Mechanistic model to predict colostrums intake based on deuterium oxide dilution technique data and impact of gestation and prefarrowing diets on piglet intake and sow yield of colostrums. J. Anim. Sci. 2014, 92, 5507–5519. [Google Scholar] [CrossRef] [PubMed]
- Decaluwé, R.; Maes, D.; Declerck, I.; Cools, A.; Wuyts, B.; De Smet, S.; Janssens, G.P.J. Changes in back fat thickness during late gestation predict colostrum yield in sows. Animal 2013, 7, 1999–2007. [Google Scholar] [CrossRef] [Green Version]
- Quesnel, H.; Farmer, C. Review: Nutritional and endocrine control of colostrogenesis in swine. Animal 2019, 13, S26–S34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clowes, E.J.; Williams, I.H.; Baracos, V.E.; Pluske, J.R.; Cegielski, A.C.; Zak, L.J.; Aherne, F.X. Feeding lactating primiparous sows to establish three divergent metabolic states: II. effect on nitrogen partitioning and skeletal muscle composition 1,2. J. Anim. Sci. 1998, 76, 1154–1164. [Google Scholar] [CrossRef] [Green Version]
- Trevisi, P.; Luise, D.; Won, S.; Salcedo, J.; Bertocchi, M.; Barile, D.; Bosi, P. Variations in porcine colostrum oligosaccharide composition between breeds and in association with sow maternal performance. J. Anim. Sci. Biotechnol. 2020, 11, 1–11. [Google Scholar] [CrossRef]
- Wagstrom, E.A.; Yoon, K.; Zimmerman, J.J. Immune components in porcine mammary secretions. Viral Immunol. 2000, 13, 383–397. [Google Scholar] [CrossRef] [PubMed]
- Klobasa, F.; Habe, F.; Werhahn, E.; Butler, J.E. Changes in the concentrations of serum IgG, IgA and IgM of sows throughout the reproductive cicle. Vet. Immunol. Immunopathol. 1985, 10, 341–353. [Google Scholar] [CrossRef]
- Farmer, C.; Quesnel, H. Nutritional, hormonal, and environmental effects on colostrum in sows. J. Anim. Sci. 2009, 87, 56–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curtis, J.; Bourne, F.J. Immunoglobulin quantitation in sow serum, colostrum and milk and the serum of young pigs. BBA Protein Struct. 1971, 236, 319–332. [Google Scholar] [CrossRef]
- Hurley, W.L.; Theil, P.K. Perspectives on immunoglobulins in colostrum and milk. Nutrients 2011, 442–474. [Google Scholar] [CrossRef] [PubMed]
- Markowska-Daniel, I.; Pomorska-Mól, M.; Pejsak, Z. The influence of age and maternal antibodies on the postvaccinal response against swine influenza viruses in pigs. Vet. Immunol. Immunopathol. 2011, 142, 81–86. [Google Scholar] [CrossRef]
- Bernabucci, U.; Basiricó, L.; Morera, P. Impact of hot environment on colostrum and milk composition. Cell. Mol. Biol. 2013, 59, 67–83. [Google Scholar] [CrossRef]
- Rooke, J.A.; Bland, I.M. The acquisition of passive immunity in the new-born piglet. Livest. Prod. Sci. 2002, 78, 13–23. [Google Scholar] [CrossRef]


{kind=link}
| Item | Paritynumber | SEM | p-Value | ||||
|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | ≥5 | |||
| Backfat thickness 1, mm | 16.5 | 21.6 | 20.4 | 15.9 | 16.1 | 0.818 | 0.086 |
| Loin depth 1, mm | 46.9 | 42.7 | 44.5 | 41.9 | 40.1 | 1.118 | 0.321 |
| Body condition score 1 | 2.8 | 2.7 | 3.1 | 2.8 | 2.6 | 0.086 | 0.588 |
| NBA 2 | 13.0 | 11.0 | 12.5 | 13.3 | 12.7 | 0.451 | 0.541 |
| NBD 3 | 0.7 | 2.2 | 2.0 | 0.9 | 1.8 | 0.237 | 0.157 |
| LBW 4, kg | 17.4 | 19.4 | 18.0 | 20.8 | 19.6 | 0.629 | 0.443 |
| PBW 5, kg | 1.3 b | 1.6 a | 1.4 ab | 1.5 a | 1.4 ab | 0.031 | 0.034 |
| Gestation length, days | 114.5 | 115.0 | 114.5 | 114.8 | 114.4 | 0.128 | 0.538 |
| Item | Parity Number | Time (h) 1 | SEM | p-Value | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | ≥5 | 0 1 | 6 | 24 | Parity | h | P × h | ||
| DM 2, % | 28.1 a | 25.5 b | 23.9 b | 24.3 b | 25.2 b | 29.3 a | 24.0 b | 22.9 b | 0.351 | 0.001 | <0.001 | 0.454 |
| Protein, % | 14.6 | 13.6 | 12.8 | 12.5 | 14.0 | 18.3 a | 13.5 b | 8.7 c | 0.293 | 0.150 | <0.001 | 0.625 |
| Fat, % | 8.0 a | 7.0 ab | 6.1 b | 6.9 ab | 6.1 b | 6.1 b | 5.5 b | 8.8 a | 0.223 | 0.029 | <0.001 | 0.505 |
| Lactose, % | 3.9 a | 3.3 b | 3.5 b | 3.6 ab | 3.5 b | 3.0 c | 3.5 b | 4.2 a | 0.065 | 0.024 | <0.001 | 0.441 |
| NFS 3, % | 20.1 a | 18.5 b | 17.8 b | 17.4 b | 19.0 ab | 23.1 a | 18.5 b | 14.1 c | 0.268 | 0.015 | <0.001 | 0.469 |
| SCC 4, (×103/mL) | 5217 | 5718 | 3398 | 9397 | 2117 | 4229 | 5132 | 6139 | 1.069 | 0.323 | 0.742 | 0.447 |
| Item | Parity Number | SEM | p-Value | ||||
|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | ≥5 | |||
| IgG (mg/mL) | 18.4 | 20.6 | 23.4 | 18.9 | 27.9 | 1.361 | 0.195 |
| IgM (mg/mL) | 10.4 | 9.6 | 10.4 | 8.6 | 9.6 | 0.416 | 0.610 |
| IgA (mg/mL) | 1.5 ab | 1.3 ab | 1.3 ab | 1.0 b | 2.3 a | 0.123 | 0.045 |
| Item | Paritynumber | Time (h) 1 | SEM | p-Value | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | ≥5 | 0 | 6 | 24 | P | H | P × h | ||
| IgG (mg/mL) | 63.3 | 62.3 | 76.4 | 63.1 | 71.8 | 94.3 a | 70.6 b | 37.3 c | 1.891 | 0.349 | <0.001 | 0.398 |
| IgM (mg/mL) | 4.6 | 4.4 | 3.7 | 4.9 | 4.9 | 5.0 a | 4.7 a | 3.7 b | 0.159 | 0.201 | 0.005 | 0.947 |
| IgA (mg/mL) | 8.1 | 9.1 | 7.4 | 7.4 | 6.5 | 9.9 a | 8.2 b | 5.1 c | 0.233 | 0.167 | <0.001 | 0.978 |
| Serum IgA | Serum IgM | Colostrum IgG | Colostrum IgA | Colostrum IgM | |
|---|---|---|---|---|---|
| Serum IgG | 0.006 | 0.082 | 0.571 *** | −0.063 | 0.438 ** |
| Serum IgA | 0.072 | 0.087 | 0.159 | 0.098 | |
| Serum IgM | 0.007 | −0.215 | 0.013 | ||
| Colostrum IgG | 0.115 | 0.995 *** | |||
| Colostrum IgA | 0.184 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Segura, M.; Martínez-Miró, S.; López, M.J.; Madrid, J.; Hernández, F. Effect of Parity on Reproductive Performance and Composition of Sow Colostrum during First 24 h Postpartum. Animals 2020, 10, 1853. https://doi.org/10.3390/ani10101853
Segura M, Martínez-Miró S, López MJ, Madrid J, Hernández F. Effect of Parity on Reproductive Performance and Composition of Sow Colostrum during First 24 h Postpartum. Animals. 2020; 10(10):1853. https://doi.org/10.3390/ani10101853
Chicago/Turabian StyleSegura, Mónica, Silvia Martínez-Miró, Miguel José López, Josefa Madrid, and Fuensanta Hernández. 2020. "Effect of Parity on Reproductive Performance and Composition of Sow Colostrum during First 24 h Postpartum" Animals 10, no. 10: 1853. https://doi.org/10.3390/ani10101853