1. Introduction
The intestinal mucosa of piglets has notably high energy demands because of their physiological functions, including the absorption and transport of nutrients, signal transduction, as well as the renewal of epithelial cells and maintenance of the structure [
1]. Furthermore, piglets can make good use of breast milk to provide energy when they are not weaned, because they have a high lipase activity from the moment of birth [
2]. However, its nutritional supply changes from high digestibility breast milk rich in protein, fat and lactose to a starch-based diet with lower digestibility after weaning [
3,
4]. Meanwhile the inadequate feed intake immediately after weaning results in an energy intake that is insufficient for maintaining the structure and function of the intestinal epithelium [
5]. A previous study showed that early weaning downregulated intestinal epithelial energy production in piglets, including the tricarboxylic acid cycle pathway, fatty acid oxidation and glycolysis [
6]. Thus, a normal mucosal energy status is an important guarantee for relieving weaning stress and improving the growth performance of piglets [
7].
Medium-chain fatty acid glyceride is an ester formed from glycerol and fatty acid. Because of its small molecular weight, small volume and strong water solubility, it is easier to absorb. Medium-chain fatty acid glycerides are decomposed into medium-chain fatty acids (MCFAs) in the small intestine, and can then quickly enter the liver through the portal vein for oxidation to provide energy.
Furthermore, compared with long-chain fatty acids (LCFAs), the oxidation of medium-chain fatty acids is independent of the carnitine system and can be activated directly in the mitochondrial matrix [
8]. The unoxidized MCFAs could synthesize triglyceride (TG) by synthesizing LCFAs, after which TG was transported to adipose tissue for storage by forming chylomicron [
9]. According to Dierick et al., medium-chain triacylglycerols have a good antibacterial effect [
10,
11]. Due to the change of feed during weaning, many undigested nutrients in piglets provide a suitable hotbed for the reproduction and development of bacteria [
12]. Caprylic acid and lauric acid have ruined the cytoplasmic structure of bacteria, and caprylic have significantly reduced the number of
Escherichia coli and
salmonella [
13].
Because of the incomplete intestinal development of piglets, piglets are sensitive to the protein level in diets. The level of protein is an important factor affecting the growth performance and intestinal health of piglets [
14,
15]. The undigested protein of piglets will enter the large intestine for fermentation in order to produce toxic metabolites, which will destroy the integrity of the intestinal epithelium and damage the health of the intestinal tract [
16]. Many studies have shown that reducing protein levels can reduce the diarrhea rate and N excretion of piglets, and can reduce protein fermentation in the intestinal tract [
17,
18].
Therefore, the purpose of this study was to investigate the effects of dietary supplementation with medium-chain fatty acid glyceride on the growth performance and intestinal health of weaned piglets, based on a low-protein diet that was beneficial for relieving the nutritional burden [
19,
20]. In this experiment, 21-day-old weaned piglets were fed with two kinds of medium-chain fatty acid triglycerides at a low protein diet level, and the antibiotic group and normal protein group were compared at the same time. The growth performance, intestinal permeability, intestinal morphology, intestinal tight junction proteins and intestinal cytokines of piglets were detected to provide a scientific basis for the application of medium-chain fatty acid triglycerides in low-protein diets.
2. Materials and Methods
This study was conducted in accordance with the guidelines of the Institute of Subtropical Agriculture, Chinese Academy of Sciences. All the experimental schemes were approved by the Animal Ethics Committee of the Institute of Subtropical Agriculture, Chinese Academy of Sciences. The animal experiments project identification code is IACUC#20190615, and the approval date is 15 May 2019.
2.1. Experimental Animal and Sample Collection
A total of 90 healthy Duroc × Landrace × Large Yorkshire piglets weaned at 21 days of age (body weight 6 ± 0.15 kg) were randomly assigned to five treatments, with six pens per treatment and three piglets per pen. The study design is shown in
Table 1: NP (Normal protein treatment, normal-protein diet no antibiotics included); NC (Negative control, low-protein diet no antibiotics included); PC (Positive control, low-protein diet +75 mg/kg quinocetone, 20 mg/kg virginiamycin and 50 mg/kg aureomycin); MCT (low-protein diet + tricaprylin/tricaprin); GML (low-protein diet +glycerol monolaurate). The normal protein basal diet and low-protein basal diet were formulated according to the nutrient requirements for weanling piglets (NRC, 2012) and previous studies [
21] (
Table 2). The medium-chain fatty acid glycerides were obtained from Deyuanshun Biological Technology Co., Ltd. (Beijing, China). The experiment lasted for 14 days, and all piglets were freely fed.
On the second morning after the 14th day, we collected the blood and tissue. After overnight fasting, we collected blood from the jugular vein of piglets in the morning. Approximately 10 mL of blood from the jugular vein was collected in aseptic capped tubes containing 150 U of sodium heparin and an ordinary centrifuge tube. Serum and plasma samples were obtained by centrifugation at 3000× g for 10 min at 4 ℃ and were stored at −80 ℃ for biochemical detection. Six piglets from each treatment were anesthetized with sodium pentobarbital (20 mg/kg BW) and killed by jugular puncture. After slaughtering, the samples of jejunum and ileum were immediately snap-frozen in liquid nitrogen and then transferred to −80 ℃ for further analysis. The jejunum and ileum (around 2 cm) was fixed in 4% formalin to detect the morphology of the intestine. The chyme of the colon was collected in a 50 mL centrifuge tube and then transferred to −80 ℃ for a fatty acid determination.
2.2. Serum Biochemical Indexes Assays
Biochemical indicators (D-LACT: mmol/L, DAO: mmol/L) were measured using an instrument (Biochemical Analytical Instrument, Beckman CX4, Beckman Coulter Inc., Brea, CA, USA) and commercial kits (Sino-German Beijing Leadman Biotech Ltd., Beijing, China).
2.3. Detection of Intestinal Morphology and Structure
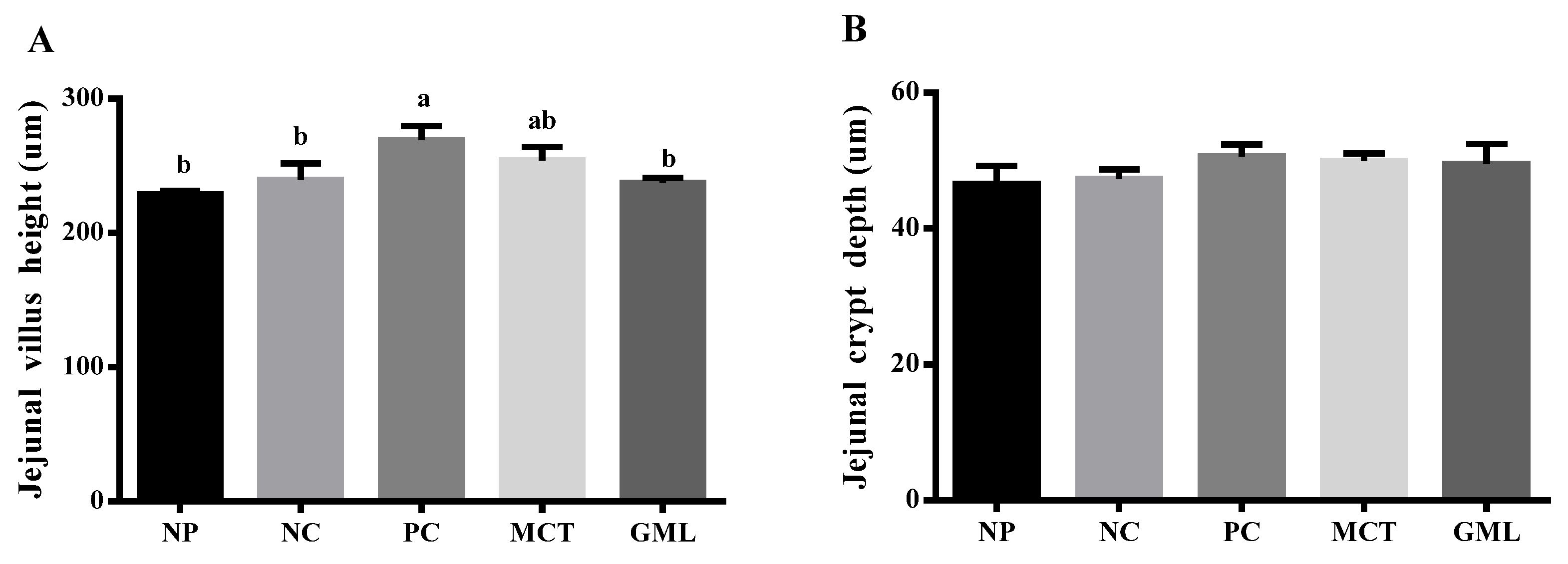
HE staining was used to detect the morphology of the jejunum and ileum. The jejunum and ileum samples were cut open and dehydrated in a tissue-processing machine for 16 h before embedding paraffin. Each section was cut into a thickness of about 4 um, and was then fixed in a glass slide and heated at 57 ℃ until the sample was dry. The sample was stained with hematoxylin and eosin and loaded with cover slides. The villus height and crypt depth of the jejunum and ileum were measured by microscope.
2.4. Determination of Cytokines in the Serum and Intestine
The serum, jejunum and ileum concentrations of SIgA, interleukin-1 beta (IL-1β), interleukin-6 (IL-6), tumor necrosis factor-alpha (TNF-α) and interferon-gamma (IFN-γ) were measured using commercially available swine enzyme-linked immunosorbent assay (ELISA) kits, according to the manufacturer’s instructions (Meimian Industrial Co., Ltd., Nanjing, Jiangsu, China).
2.5. Immunohistochemical Analysis
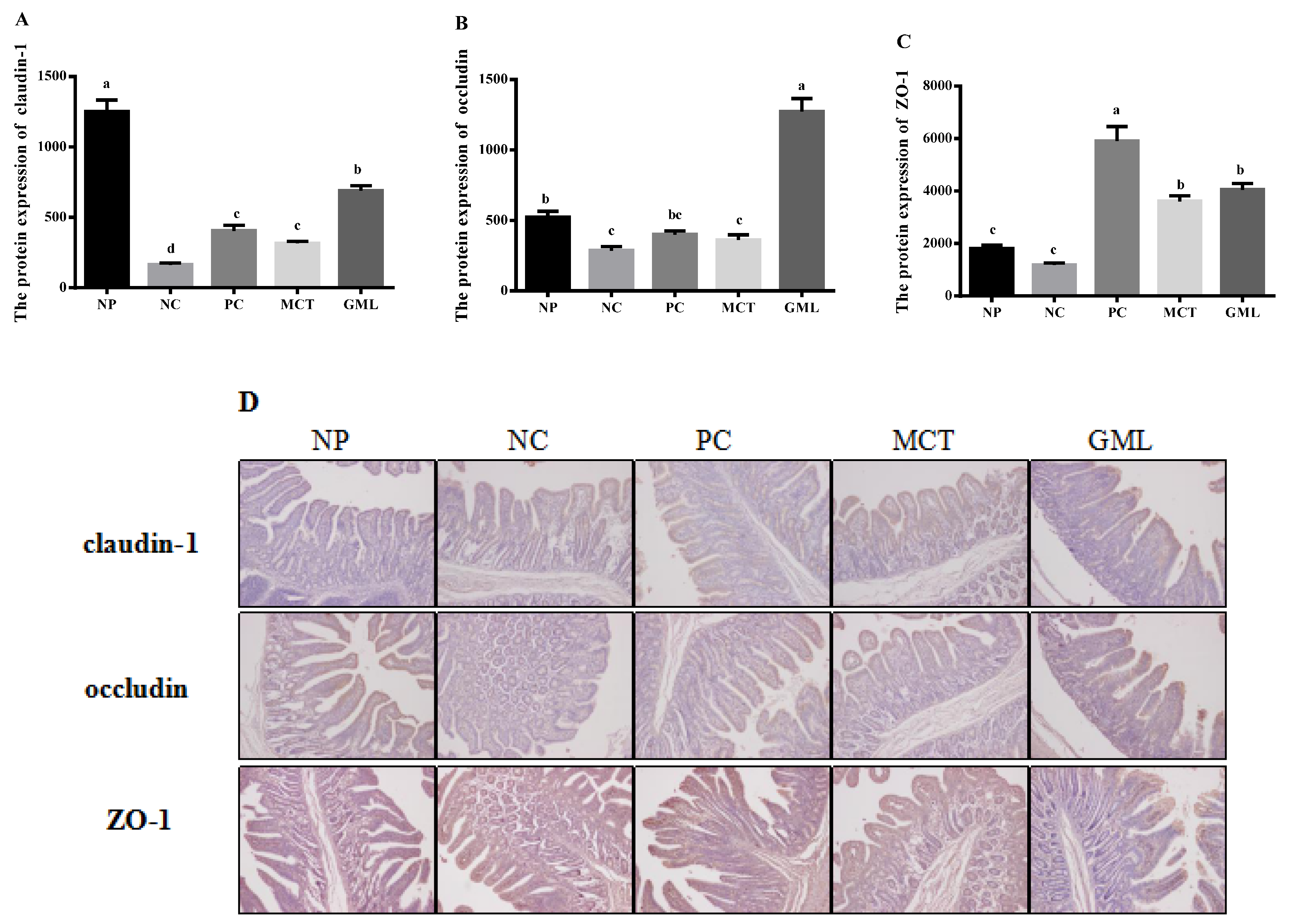
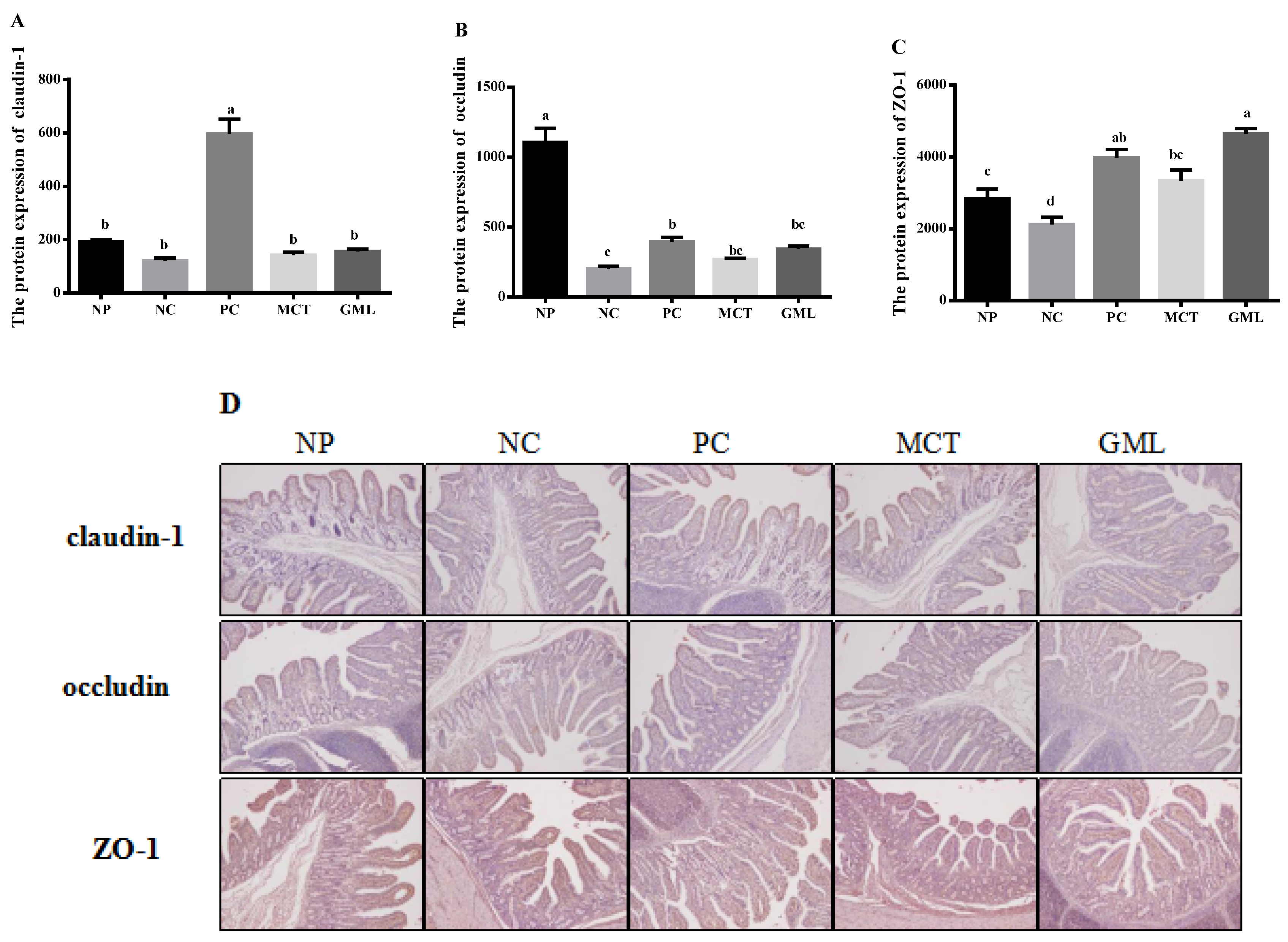
The protein expressions of claudin-1, occluding and ZO-1 in the jejunum and ileum of piglets were determined using an immunohistochemical analysis. The tissue blocks were fixed with 14% paraformaldehyde, and then continuous paraffin sections were made. The paraffin sections were baked in an oven at 65 ℃ for 2 h, washed with PBS three times for 5 min each time, before being repaired by microwave in ethylenediamine tetraacetic acid (EDTA) buffer. And after the medium heat boils, the power is cut off, and then after 10 min, it boils on a low heat. After this, they were washed three times with PBS, as before, before being put in a 3% hydrogen peroxide solution, incubated 10 min away from light at room temperature, washed with PBS three times for 5 min each time, and sealed with 5% Bovine Serum Albumin (BSA) for 20 min after drying. Then, we removed the BSA solution, added about 50 μL diluted LRP6 covering tissue to each slice, which were then left overnight at 4 ℃. After this, they were washed with Phosphate Buffer Saline (PBS) three times, and 50–100 μL of the corresponding species of HRP labeled goat anti-rabbit (purchased from BOSTER company) was added to the slices, which were incubated at 37 ℃ for 50 min. They were PBS-washed three times again, and, except for the PBS solution, 50–100 μL freshly prepared DAB solution was added to each slice, followed by a microscope-controlled color development. After the complete color development, they were rinsed with distilled water or tap water, followed by hematoxylin redyeing, 1% hydrochloric acid alcohol differentiation (about 1 s), a tap water rinse, ammonia return to blue, and a running water rinse. The slices were treated with gradient alcohol for 10 min, dehydrated and dried, transparent xylene was used, and the slices were sealed with neutral gum. Staining sections were independently reviewed and scored by two researchers using a 400-fold magnification microscope (Olympus, Tokyo, Japan).
2.6. Statistical Analyses
Data were analyzed by an analysis of variance, using the General Linear Models procedure of the SPSS 20.0 (SPSS Inc., Chicago, IL, USA). Significant differences between means were determined using the Tukey’s multiple comparison tests. Results were expressed as the mean ± standard error of the mean (SEM). A value of p < 0.05 was considered statistically significant. Mean values and a statistical elaboration were performed by using each pen as the experimental unit (n = 6 per treatment).
4. Discussion
The present study demonstrates the promotion of the intestinal health of medium-chain fatty acids, which is key to maximizing growth performance in livestock. In our study, dietary supplementation with medium-chain fatty acid glycerides in a low-protein and antibiotic-free diet improved the growth performance, intestinal barrier and inflammatory reaction of weaning piglets.
First, the addition of medium-chain fatty acid glycerides in diets might inhibit the increase of the intestinal mucosal permeability of piglets caused by weaning stress. DAO is closely related to the maturity and integrity of intestinal mucosal cells [
22,
23], and D-LACT can enter the bloodstream through the damaged mucosa [
24]. Both of them can be used as an index to detect intestinal permeability. In our study, the D-LACT of serum decreased in the MCT treatment, which may indicate that the intestinal permeability decreased in the piglets. This is consistent with previous studies that showed that MCT could reduce the permeability of CA in Caco-2 cells [
25] and inhibit the increase in intestinal permeability induced by LPS in mice [
26].
Second, medium-chain fatty acid glycerides could promote the intestinal maturation of piglets, reduce the injury of weaning stress and increase the absorption of nutrients in piglets. The epithelial cells near the villous tip have the strongest digestion and absorption ability of nutrients [
27], and thus the increase of epithelial cells and the increase of the villus height can effectively promote the absorption of nutrients [
28]. However, it is easy to cause the villus height of piglets to decrease during weaning [
29,
30]. In our study, the treatment with medium-chain fatty acid glycerides helped increase the villus height and the number of goblet cells, which was consistent with a previous study [
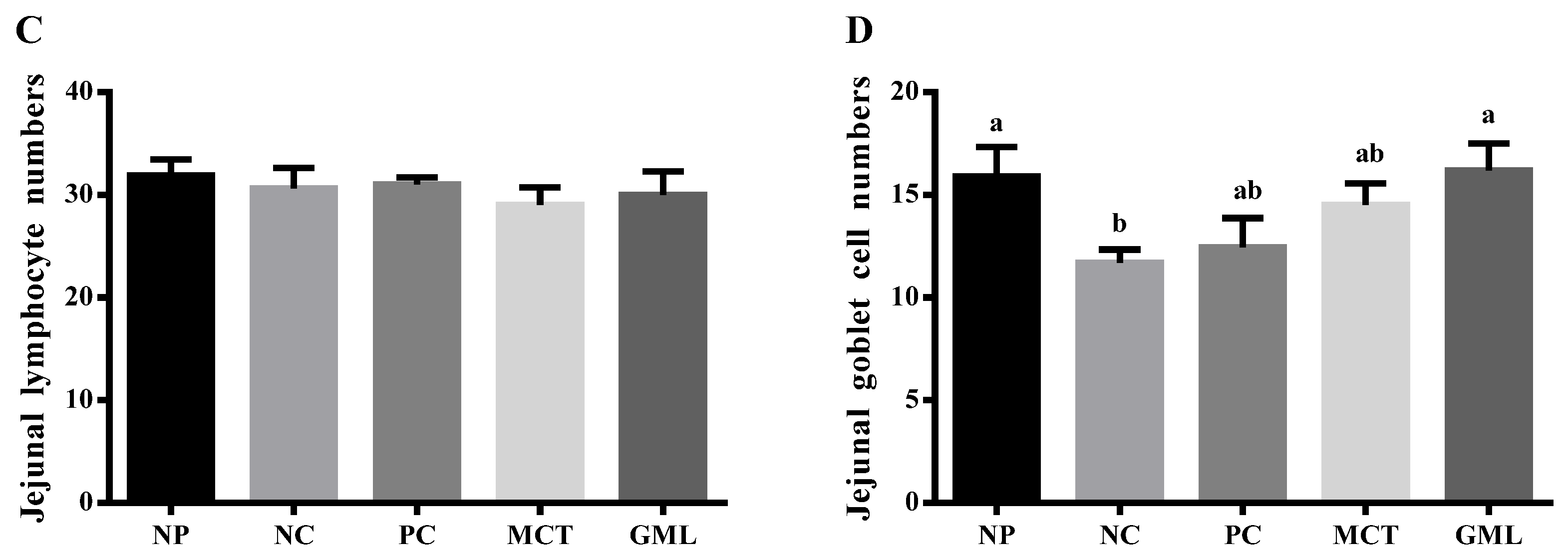
31]. Therefore, it could possibly promote the maturation of intestinal epithelial cells and villi in order to absorb more nutrients by increasing the number of goblet cells and the height of villi.
Third, it was further proven that medium-chain fatty acid glycerides could effectively protect the integrity of the intestinal barrier, given that the protein expressions of claudin-1, occludin and ZO-1 were significantly increased. Claudin-1, occludin and ZO-1 are important components of the intestinal barrier [
32]. The bacteria and toxins can regulate and control the expression of intestinal occludin and ZO-1 by regulating or influencing cytokines and protein kinase C [
33]. By adding medium-chain fatty acid glycerides in our study, the claudin-1, occludin and ZO-1 increased significantly, consistent with a previous study that showed that medium-chain fatty acid glycerides could readjust the distribution of occludin and ZO-1 [
34]. The reason for this may be that they could effectively prevent bacterial endotoxin and toxic macromolecules from entering the body in order to increase the expression of claudin-1, occludin and ZO-1 proteins, thus protecting the health of the intestinal barrier.
In addition, medium-chain glycerides can protect intestinal barrier health by reducing the production of proinflammatory factors. Many studies have shown that weaning stress can easily lead to the production of proinflammatory factors in piglets, including IL-1
β, IL - 6, TNF-
α, IFN-γ [
35,
36]. SIgA is the first protective barrier of the intestinal epithelium and can protect intestinal epithelial cells from enterotoxins and pathogenic microorganisms [
37]. In our study, MCT significantly decreased the level of IL-6 in the ileum when compared with the NC treatment, and both MCT and GML significantly increased the level of SIgA in the ileum when compared with the NC treatment. This agrees with a previous study, according to which medium-chain glycerides could significantly reduce the level of proinflammatory cytokines, such as IL-6, IL-1, and relieve the inflammation of colitis in rats [
38,
39]. These data show that medium-chain fatty acid glycerides can, to a certain extent, alleviate the intestinal inflammatory response caused by weaning stress. The reduction of inflammation is more conducive to the protection of the intestinal barrier and plays a positive role in the growth of piglets.

 ,
, 







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}