
Re-Emergence and Spread of Haemorrhagic Septicaemia in Germany: The Wolf as a Vector?

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Isolates
2.2. Spatiotemporal Analysis of HS Outbreaks
2.3. Whole Genome Sequencing (WGS)
2.4. Multilocus Sequence Typing and Phylogenetic Analysis
2.5. Determination of Virulence-Associated Genes
3. Results
3.1. Determination of P. multocida Capsular Type and HS Specific DNA Fragment KTT72/KTSP61
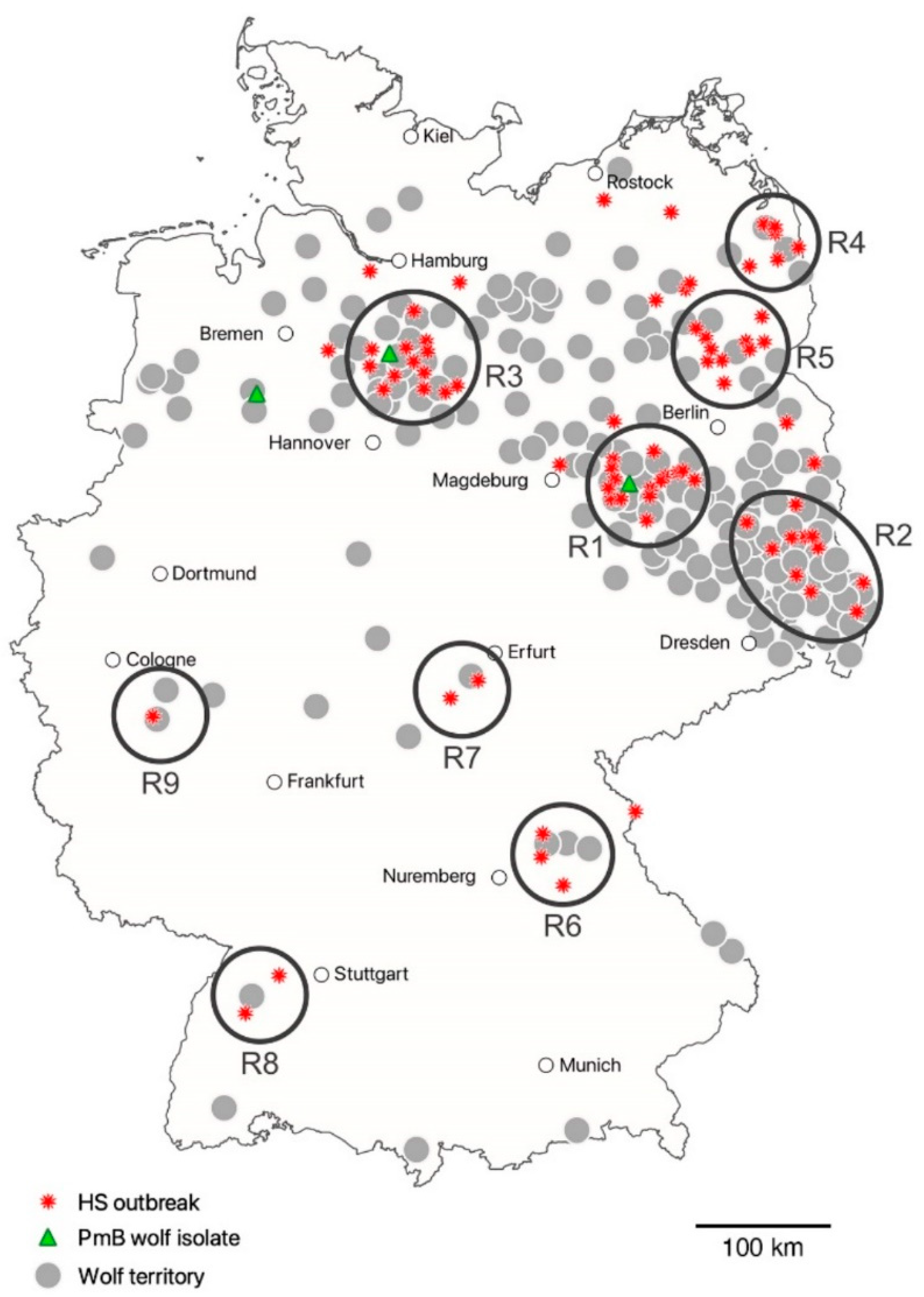
3.2. Spatiotemporal Analysis of HS Outbreaks in Respect of Proven Wolf Territories
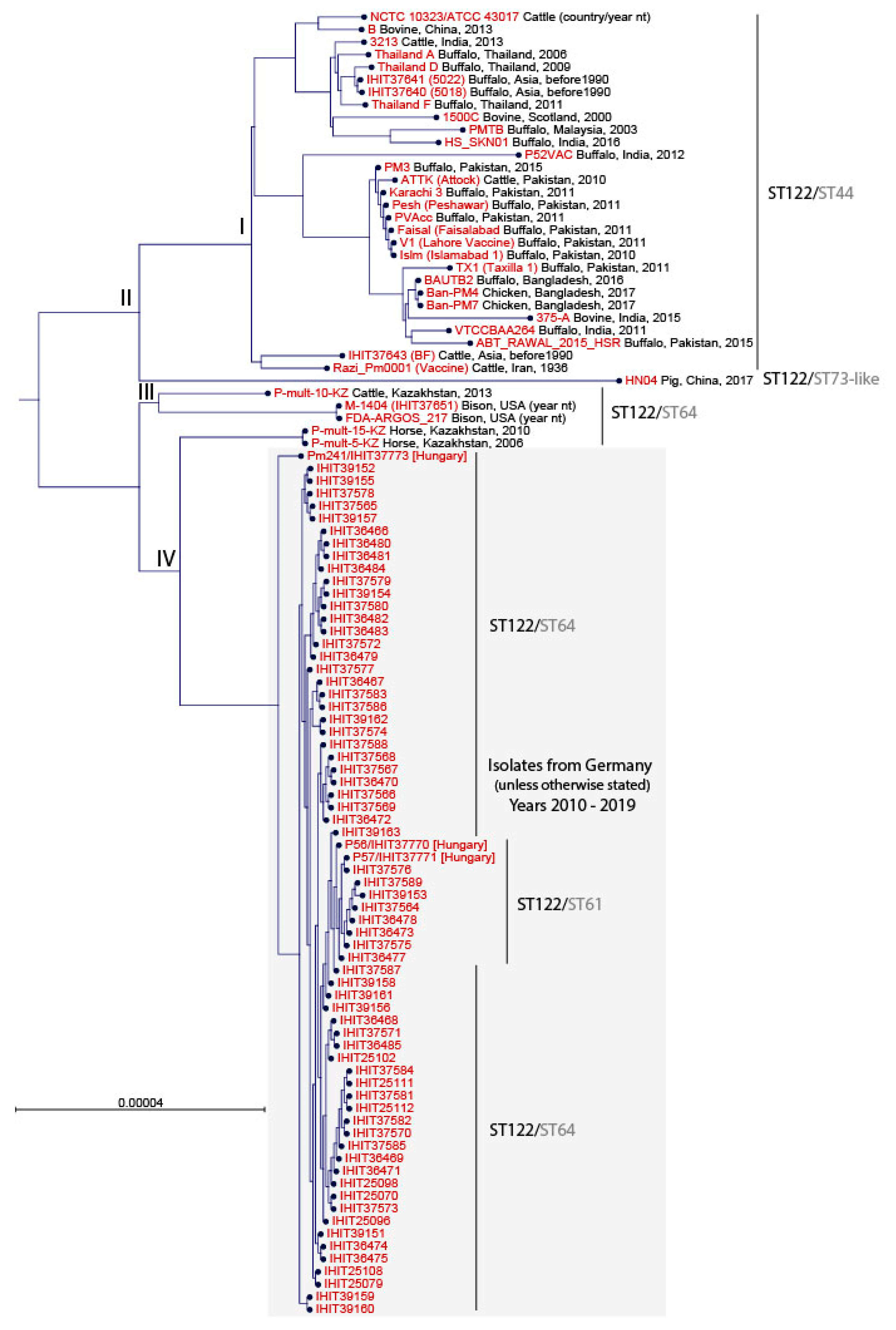
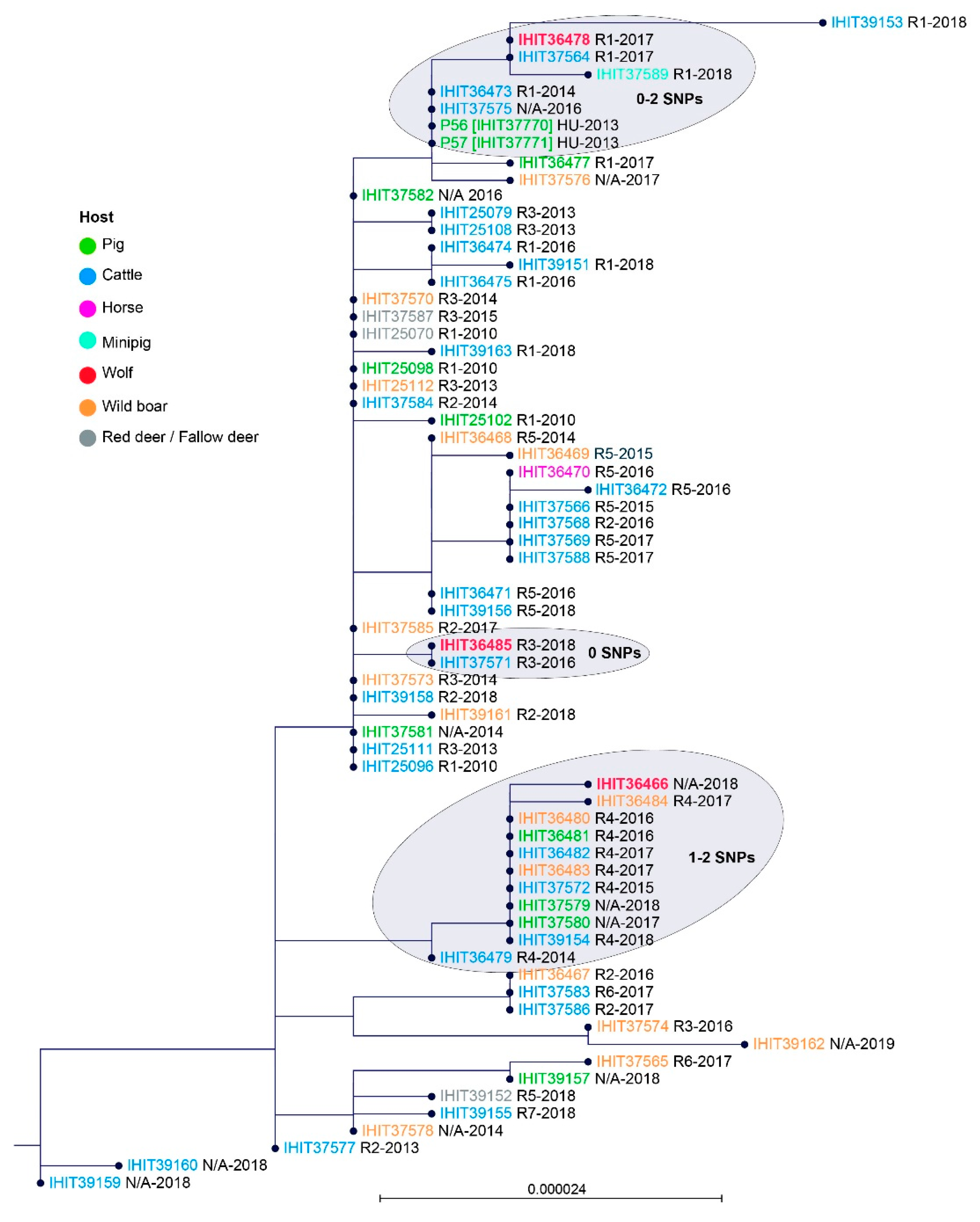
3.3. Multilocus Sequence Typing and Core Genome Comparison
3.4. Distribution of Capsule, LPS, and Virulence-Associated Genes among PmB-ST122 Strains
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Haemorrhagic Septicaemia (Chapter 3.4.10.) in: Manual of Diagnostic Tests and Vaccines for Terrestrial Animals 2021. Available online: https://www.oie.int/en/what-we-do/standards/codes-and-manuals/terrestrial-manual-online-access/ (accessed on 12 July 2021).
- Cuevas, I.; Carbonero, A.; Cano-Terriza, D.; Pacheco, I.L.; Marín, J.C.; Borge, C. First outbreak of bovine haemorrhagic septicaemia caused by Pasteurella multocida type B in Spain – Short communication. Acta Veter. Hung. 2020, 68, 8–11. [Google Scholar] [CrossRef] [PubMed]
- Dunbar, M.R.; Wolcott, M.J.; Rimler, R.B.; Berlowski, B.M. Septicemic Pasteurellosis in Free-ranging Neonatal Pronghorn in Oregon. J. Wildl. Dis. 2000, 36, 383–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eriksen, L.; Aalbaek, B.; Leifsson, P.S.; Basse, A.; Christiansen, T.; Eriksen, E.; Rimler, R.B. Hemorrhagic septicemia in fallow deer (Dama dama) caused by Pasteurella multocida multocida. J. Zoo Wildl. Med. 1999, 30, 285–292. [Google Scholar] [PubMed]
- Franson, J.C.; Smith, B.L. Septicemic Pasteurellosis in Elk (Cervus elaphus) on the United States National Elk Refuge, Wyoming. J. Wildl. Dis. 1988, 24, 715–717. [Google Scholar] [CrossRef] [PubMed]
- Orynbayev, M.; Sultankulova, K.; Sansyzbay, A.; Rystayeva, R.; Shorayeva, K.; Namet, A.; Fereidouni, S.; Ilgekbayeva, G.; Barakbayev, K.; Kopeyev, S.; et al. Biological characterization of Pasteurella multocida present in the Saiga population. BMC Microbiol. 2019, 19, 1–10. [Google Scholar] [CrossRef]
- Soike, D.; Schulze, C.; Kutzer, P.; Ewert, B.; van der Grinten, E.; Schliephake, A.; Ewers, C.; Bethe, A.; Rau, J. Acute pas-teurellosis in fallow deer, cattle and pigs in a region of Eastern Germany. Berl. Munch. Tierarztl. Wochenschr. 2012, 125, 122–128. [Google Scholar] [CrossRef] [PubMed]
- Mackie, J. Haemorrhagic septicaemia in pigs. Aust. Veter. J. 1996, 74, 399–400. [Google Scholar] [CrossRef]
- Risco, D.; Fernández-Llario, P.; Cuesta, J.M.; García-Jiménez, W.L.; Gil Molino, M.; Gonçalves, P.; Martínez, R.; Gómez, L.; Garcia-Sanchez, A.; Rey, J.; et al. Fatal outbreak of systemic pasteurellosis in a wild boar (Sus scrofa) population from southwest Spain. J. Veter. Diagn. Investig. 2013, 25, 791–794. [Google Scholar] [CrossRef] [Green Version]
- Townsend, K.M.; Frost, A.J.; Lee, C.W.; Papadimitriou, J.M.; Dawkins, H.J.S. Development of PCR Assays for Species- and Type-Specific Identification of Pasteurella multocida Isolates. J. Clin. Microbiol. 1998, 36, 1096–1100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ujvari, B.; Szeredi, L.; Pertl, L.; Tóth, G.; Erdélyi, K.; Jánosi, S.; Molnár, T.; Magyar, T. First detection of Pasteurella multocida type B:2 in Hungary associated with systemic pasteurellosis in backyard pigs. Acta Veter. Hung. 2015, 63, 141–156. [Google Scholar] [CrossRef] [Green Version]
- Townsend, K.M.; Boyce, J.D.; Chung, J.Y.; Frost, A.J.; Adler, B. Genetic Organization of Pasteurella multocida cap Loci and Development of a Multiplex Capsular PCR Typing System. J. Clin. Microbiol. 2001, 39, 924–929. [Google Scholar] [CrossRef] [Green Version]
- Brogden, K.A.; Rhoades, K.R.; Heddleston, K.L. A New Serotype of Pasteurella multocida Associated with Fowl Cholera. Avian Dis. 1978, 22, 185. [Google Scholar] [CrossRef] [PubMed]
- Heddleston, K.L.; E Gallagher, J.; A Rebers, P. Fowl cholera: Gel diffusion precipitin test for serotyping Pasteruella multocida from avian species. Avian Dis. 1972, 16, 925–936. [Google Scholar] [CrossRef] [PubMed]
- Harper, M.; John-White, M.; Turni, C.; Edmunds, M.; Michael, F.S.; Adler, B.; Blackall, P.; Cox, A.; Boyce, J.D. Development of a Rapid Multiplex PCR Assay To Genotype Pasteurella multocida Strains by Use of the Lipopolysaccharide Outer Core Biosynthesis Locus. J. Clin. Microbiol. 2014, 53, 477–485. [Google Scholar] [CrossRef] [Green Version]
- Moustafa, A.M.; Bennett, M.D.; Edwards, J.; Azim, K.; Mesaik, M.A.; Choudhary, M.I.; Pathanasophon, P.; Worarach, A.; Ali, Q.; Abubakar, M.; et al. Molecular typing of haemorrhagic septicaemia-associated Pasteurella multocida isolates from Pakistan and Thailand using multilocus sequence typing and pulsed-field gel electrophoresis. Res. Veter. Sci. 2013, 95, 986–990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, Z.; Wang, X.; Zhou, R.; Chen, H.; Wilson, B.A.; Wu, B. Pasteurella multocida: Genotypes and Genomics. Microbiol. Mol. Biol. Rev. 2019, 83. [Google Scholar] [CrossRef] [PubMed]
- Sting, R.; Blazey, B.; Schwabe, I.; Schwalm, A.K.; Müller, S.; Sprague, L.D.; Rau, J. Haemorrhagic septicaemia (septicaemic pasteurellosis) in cattle in Baden-Wuerttemberg (Germany). Berl. Munch. Tierarztl. Wochenschr. 2020, 133. [Google Scholar] [CrossRef]
- Annas, S.; Zamri-Saad, M.; Jesse, F.F.A.; Zunita, Z. New sites of localisation of Pasteurella multocida B:2 in buffalo surviving experimental haemorrhagic septicaemia. BMC Veter. Res. 2014, 10, 88. [Google Scholar] [CrossRef] [Green Version]
- Shivachandra, S.B.; Viswas, K.N.; Kumar, A.A. A review of hemorrhagic septicemia in cattle and buffalo. Anim. Heal. Res. Rev. 2011, 12, 67–82. [Google Scholar] [CrossRef]
- Kędrak, A.; Borkowska-Opacka, B. Phenotypic characteristics of Pasteurella multocida strains isolated from cattle affected with haemorrhagic septicaemia. Bull. Vet. Inst. Pulawy 2001, 45, 171–176. [Google Scholar]
- Magyar, T.; Ujvári, B.; Szeredi, L.; Virsinger, N.; Albert, E.; Német, Z.; Csuka, E.; Biksi, I. Re-emergence of bovine haemorrhagic septicaemia in Hungary. Acta Veter. Hung. 2017, 65, 41–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rohkohl, J.; Schulze, C.; Bilk, S. Hemorrhagic septicemia on a dairy farm in Lower Saxony. Prakt Tierarzt. 2015, 96, 598–608. [Google Scholar]
- World Animal Health Information Database (WAHIS) Interface. Available online: https://www.oie.int/en/animal-health-in-the-world/the-world-animal-health-information-system/data-before-2005-handistatus (accessed on 12 July 2021).
- DBBW. Dokumentations- und Beratungsstelle des Bundes zum Thema Wolf. Available online: https://www.dbb-wolf.de/wolf-occurrence (accessed on 12 July 2021).
- QGIS Association; QGIS.org. QGIS Geographic Information System. Available online: https://qgis.org (accessed on 11 November 2020).
- Roehr, J.T.; Dieterich, C.; Reinert, K. Flexbar 3.0 – SIMD and multicore parallelization. Bioinformatics 2017, 33, 2941–2942. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Seemann, T. Prokka: Rapid Prokaryotic Genome Annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [PubMed]
- Page, A.J.; Cummins, C.A.; Hunt, M.; Wong, V.K.; Reuter, S.; Holden, M.T.G.; Fookes, M.; Falush, D.; Keane, J.A.; Parkhill, J. Roary: Rapid large-scale prokaryote pan genome analysis. Bioinformatics 2015, 31, 3691–3693. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Peng, Z.; Liang, W.; Wang, F.; Xu, Z.; Xie, Z.; Lian, Z.; Hua, L.; Zhou, R.; Chen, H.; Wu, B. Genetic and Phylogenetic Characteristics of Pasteurella multocida Isolates From Different Host Species. Front. Microbiol. 2018, 9, 1408. [Google Scholar] [CrossRef]
- Elliott, D.; Wilson, M.; Buckley, C.M.F.; Spratt, D.A. Cultivable Oral Microbiota of Domestic Dogs. J. Clin. Microbiol. 2005, 43, 5470–5476. [Google Scholar] [CrossRef] [Green Version]
- Peng, Z.; Wang, H.; Liang, W.; Chen, Y.; Tang, X.; Chen, H.; Wu, B. A capsule/lipopolysaccharide/MLST genotype D/L6/ST11 of Pasteurella multocida is likely to be strongly associated with swine respiratory disease in China. Arch. Microbiol. 2017, 200, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Kramer-Schadt, S.; Wenzler, M.; Gras, P.; Knauer, F. Habitatmodellierung und Abschätzung der potenziellen Anzahl von Wolfsterritorien in Deutschland; Bundesamt für Naturschutz: Bonn, Germany, 2020; ISBN 978-3-89624-294-5. [Google Scholar] [CrossRef]
- Hindrikson, M.; Remm, J.; Pilot, M.; Godinho, R.; Stronen, A.V.; Baltrūnaité, L.; Czarnomska, S.; Leonard, J.; Randi, E.; Nowak, C.; et al. Wolf population genetics in Europe: A systematic review, meta-analysis and suggestions for conservation and management. Biol. Rev. 2017, 92, 1601–1629. [Google Scholar] [CrossRef]
- Czarnomska, S.D.; Jędrzejewska, B.; Borowik, T.; Niedziałkowska, M.; Stronen, A.V.; Nowak, S.; Mysłajek, R.; Okarma, H.; Konopiński, M.K.; Pilot, M.; et al. Concordant mitochondrial and microsatellite DNA structuring between Polish lowland and Carpathian Mountain wolves. Conserv. Genet. 2013, 14, 573–588. [Google Scholar] [CrossRef] [Green Version]
- Steyer, C.D. Erster Wolf in der Schorfheide entdeckt. Der Tagesspiegel, 23 March 2007. Available online: https://www.tagesspiegel.de(accessed on 9 August 2021).
- Fechter, D.; Storch, I. How Many Wolves (Canis lupus) Fit into Germany? The Role of Assumptions in Predictive Rule-Based Habitat Models for Habitat Generalists. PLoS ONE 2014, 9, e101798. [Google Scholar] [CrossRef]
- Stier, N.; Meißner-Hylanová, V.; Roth, M. Wolfstelemetrie in Mecklenburg-Vorpommern, Zwischenbericht 2016; Technical University Dresden: Tharandt, Germany, 2016. [Google Scholar]
- Jędrzejewski, W.; Schmidt, K.; Theuerkauf, J.; Jędrzejewska, B.; Kowalczyk, R. Territory size of wolves Canis lupus: Linking local (Białowieża Primeval Forest, Poland) and Holarctic-scale patterns. Ecography 2007, 30, 66–76. [Google Scholar] [CrossRef]
- Kittle, A.M.; Anderson, M.; Avgar, T.; Baker, J.A.; Brown, G.S.; Hagens, J.; Iwachewski, E.; Moffatt, S.; Mosser, A.; Patterson, B.R.; et al. Wolves adapt territory size, not pack size to local habitat quality. J. Anim. Ecol. 2015, 84, 1177–1186. [Google Scholar] [CrossRef] [PubMed]
- Wagner, C.; Holzapfel, M.; Kluth, G.; Reinhardt, I.; Ansorge, H. Wolf (Canis lupus) feeding habits during the first eight years of its occurrence in Germany. Mamm. Biol. 2012, 77, 196–203. [Google Scholar] [CrossRef]
- Reinhardt, I.; Kluth, G. Terrritorial and dispersal behaviour of wolves (Canis lupus) in Germany: Results of a first telemetry study. Nat. Landsch. 2016, 91, 262–271. [Google Scholar] [CrossRef]
- Andersen, L.W.; Harms, V.; Caniglia, R.; Czarnomska, S.D.; Fabbri, E.; Jędrzejewska, B.; Kluth, G.; Madsen, A.B.; Nowak, C.; Pertoldi, C.; et al. Long-distance dispersal of a wolf, Canis lupus, in northwestern Europe. Mammal Res. 2015, 60, 163–168. [Google Scholar] [CrossRef]
- Wolfsmonitoring.com. Wolfsterritorien in Niedersachsen - Territorium Barnstorf. Available online: https://www.wolfsmonitoring.com/monitoring/wolfsterritorien (accessed on 12 July 2021).
- De Alwis, M.C. Haemorrhagic Septicaemia, ACIAR Monograph No. 57; Australian Center for International Agricultural Research: Canberra, Australia, 1999. [Google Scholar]
- De Alwis, M.C.L.; Wijewardana, T.G.; Gomis, A.I.U.; Vipulasiri, A.A. Persistence of the carrier status in haemorrhagic septicaemia (Pasteurella multocida serotype 6:B infection) in Buffaloes. Trop. Anim. Heal. Prod. 1990, 22, 185–194. [Google Scholar] [CrossRef]
- Mech, L.D. The Wolves of Isle Royale. Fauna of the National Parks of the United States, Fauna Series 7; U.S. Government Printing Office: Washington, DC, USA, 1966. [Google Scholar]
- Niedersächsisches Ministerium für Umwelt, Energie, Bauen und Klimaschutz. Infoportal Wolf. Available online: https://www.umwelt.niedersachsen.de/startseite/themen_im_fokus/der_wolf_in_niedersachsen/nutztierrisse-in-niedersachsen-174833.html (accessed on 23 August 2021).
- Landesamt für Umweltschutz Sachsen-Anhalt. Nutztierrisszahlen in Sachsen-Anhalt. Available online: https://lau.sachsen-anhalt.de/naturschutz/das-wolfskompetenzzentrum-wzi/nutztierrisse/rissstatistik-st/ (accessed on 23 August 2021).
- Talan, D.A.; Citron, D.M.; Abrahamian, F.M.; Moran, G.J.; Goldstein, E.J. Bacteriologic Analysis of Infected Dog and Cat Bites. N. Engl. J. Med. 1999, 340, 85–92. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Gene Category and Name | PmB Isolates Germany/Hungary (n = 68) | Global PmB Strains (n = 35) |
|---|---|---|
| Positive strains (%) | ||
| Adhesins/colonization factors | ||
| ptfA | 100 | 100 |
| fimA | 100 | 97.1 |
| hsf-1 | 0 | 0 |
| hsf-2 | 100 | 97.1 |
| pfhB-igB | 100 | 94.3 |
| pfhB1 | 0 | 0 |
| pfhB2 | 100 | 100 |
| tad locus | 0 | 0 |
| Toxin | ||
| toxA | 0 | 0 |
| Iron regulation/acquisition | ||
| afuCBA | 100 | 100 |
| ccmABCDEF | 100 | 97.1 |
| exbBD-tonB | 100 | 97.1 |
| fecBCDE | 100 | 100 |
| fbpABC | 100 | 100 |
| fur | 100 | 100 |
| hgbA | 0 | 0 |
| hgbB | 100 | 94.3 |
| tbpA | 0 | 77.1 |
| Extracellular enzymes | ||
| nanB | 0 | 2.9 |
| nanH | 100 | 97.1 |
| neuA | 100 | 100 |
| nanATEK | 100 | 100 |
| nanR | 0 | 0 |
| pmHAS | 0 | 0 |
| siaPT-nanM | 100 | 100 |
| Oxidative stress | ||
| sodA | 100 | 97.1 |
| sodC | 100 | 97.1 |
| Secretion systems | ||
| T2SS (“DR93_1687”) | 0 | 0 |
| T2SS/T3SS (“DR93_1692”) | 0 | 0 |
| Outer membrane proteins/protectins | ||
| ompA | 100 | 97.1 |
| ompH | 100 | 97.1 |
| oma87 | 100 | 97.1 |
| plpB | 100 | 100 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kutzer, P.; Szentiks, C.A.; Bock, S.; Fritsch, G.; Magyar, T.; Schulze, C.; Semmler, T.; Ewers, C. Re-Emergence and Spread of Haemorrhagic Septicaemia in Germany: The Wolf as a Vector? Microorganisms 2021, 9, 1999. https://doi.org/10.3390/microorganisms9091999
Kutzer P, Szentiks CA, Bock S, Fritsch G, Magyar T, Schulze C, Semmler T, Ewers C. Re-Emergence and Spread of Haemorrhagic Septicaemia in Germany: The Wolf as a Vector? Microorganisms. 2021; 9(9):1999. https://doi.org/10.3390/microorganisms9091999
Chicago/Turabian StyleKutzer, Peter, Claudia A. Szentiks, Sabine Bock, Guido Fritsch, Tibor Magyar, Christoph Schulze, Torsten Semmler, and Christa Ewers. 2021. "Re-Emergence and Spread of Haemorrhagic Septicaemia in Germany: The Wolf as a Vector?" Microorganisms 9, no. 9: 1999. https://doi.org/10.3390/microorganisms9091999