
Effects of Mixing Volatile Fatty Acids as Carbon Sources on Rhodospirillum rubrum Carbon Metabolism and Redox Balance Mechanisms

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strain, Culture Medium, and Growth Conditions
2.2. Volatile Fatty Acid Consumption Monitoring by HPLC
2.3. Polyhydroxyalkanoates Quantification
2.4. Proteomic Analysis
2.5. Isoleucine Quantification
2.6. H2 Quantification
2.7. Acetolactate Synthase Specific Activity
3. Results
3.1. Proteomic Analysis of Rs. rubrum Cultivated on a Propionate and Butyrate Mixture
3.2. Propionate and Butyrate Photoassimilation by Rs. rubrum When Present as the Sole Carbon Source
3.3. Photoassimilation of Butyrate and Propionate by Rs. rubrum When Present as a Mixture
3.3.1. Methylcitrate Cycle
3.3.2. EMC and MBC Pathways
3.3.3. Redox Homeostasis in Rs. rubrum When Butyrate and Propionate Are Used as Sole Organic Carbon Sources or as a Mixture
4. Discussion
4.1. Mixing VFAS Leads to Synergy in Their Assimilation
4.2. Mixing VFAs Abolishes the Need for Bicarbonate Supplementation for the Assimilation of Reduced Substrates
4.3. Mixing VFAs Changes the Mechanism Involved in the Redox Balance of the Cells
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Garrity, G. Bergey’s Manual® of Systematic Bacteriology: Volume Two: The Proteobacteria, Part A Introductory Essays; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2006; Volume 2, ISBN 0387280219. [Google Scholar]
- Bengtsson, S.; Werker, A.; Christensson, M.; Welander, T. Production of polyhydroxyalkanoates by activated sludge treating a paper mill wastewater. Bioresour. Technol. 2008, 99, 509–516. [Google Scholar] [CrossRef]
- Kim, D.-H.H.; Lee, J.-H.H.; Hwang, Y.; Kang, S.; Kim, M.-S.S. Continuous cultivation of photosynthetic bacteria for fatty acids production. Bioresour. Technol. 2013, 148, 277–282. [Google Scholar] [CrossRef] [Green Version]
- Fradinho, J.C.; Oehmen, A.; Reis, M.A.M.M. Photosynthetic mixed culture polyhydroxyalkanoate (PHA) production from individual and mixed volatile fatty acids (VFAs): Substrate preferences and co-substrate uptake. J. Biotechnol. 2014, 185, 19–27. [Google Scholar] [CrossRef]
- Alloul, A.; Wuyts, S.; Lebeer, S.; Vlaeminck, S.E. Volatile fatty acids impacting phototrophic growth kinetics of purple bacteria: Paving the way for protein production on fermented wastewater. Water Res. 2019, 152, 138–147. [Google Scholar] [CrossRef] [PubMed]
- Capson-Tojo, G.; Batstone, D.J.; Grassino, M.; Vlaeminck, S.E.; Puyol, D.; Verstraete, W.; Kleerebezem, R.; Oehmen, A.; Ghimire, A.; Pikaar, I. Purple phototrophic bacteria for resource recovery: Challenges and opportunities. Biotechnol. Adv. 2020, 43, 107567. [Google Scholar] [CrossRef] [PubMed]
- Alloul, A.; Cerruti, M.; Adamczyk, D.; Weissbrodt, D.; Vlaeminck, S. Control tools to selectively produce purple bacteria for microbial protein in raceway reactors. BioRxiv 2020. [Google Scholar] [CrossRef] [Green Version]
- Anterrieu, S.; Quadri, L.; Geurkink, B.; Dinkla, I.; Bengtsson, S.; Arcos-Hernandez, M.; Alexandersson, T.; Morgan-Sagastume, F.; Karlsson, A.; Hjort, M.; et al. Integration of biopolymer production with process water treatment at a sugar factory. New Biotechnol. 2014, 31, 308–323. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.-J.; Kim, B.J.; Jeong, C.-M.; Ahn, Y.H.; Chang, H.N. Anaerobic organic acid production of food waste in once-a-day feeding and drawing-off bioreactor. Bioresour. Technol. 2008, 99, 7866–7874. [Google Scholar] [CrossRef] [PubMed]
- Komemoto, K.; Lim, Y.G.; Nagao, N.; Onoue, Y.; Niwa, C.; Toda, T. Effect of temperature on VFA’s and biogas production in anaerobic solubilization of food waste. Waste Manag. 2009, 29, 2950–2955. [Google Scholar] [CrossRef]
- Zhang, P.; Chen, Y.; Zhou, Q. Waste activated sludge hydrolysis and short-chain fatty acids accumulation under mesophilic and thermophilic conditions: Effect of pH. Water Res. 2009, 43, 3735–3742. [Google Scholar] [CrossRef]
- Jiang, J.; Zhang, Y.; Li, K.; Wang, Q.; Gong, C.; Li, M. Volatile fatty acids production from food waste: Effects of pH, temperature, and organic loading rate. Bioresour. Technol. 2013, 143, 525–530. [Google Scholar] [CrossRef]
- Lee, W.S.; Chua, A.S.M.; Yeoh, H.K.; Ngoh, G.C. A review of the production and applications of waste-derived volatile fatty acids. Chem. Eng. J. 2014, 235, 83–99. [Google Scholar] [CrossRef]
- Dahiya, S.; Sarkar, O.; Swamy, Y.V.; Mohan, S.V. Acidogenic fermentation of food waste for volatile fatty acid production with co-generation of biohydrogen. Bioresour. Technol. 2015, 182, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Erb, T.J.; Berg, I.A.; Brecht, V.; Müller, M.; Fuchs, G.; Alber, B.E. Synthesis of C5-dicarboxylic acids from C2-units involving crotonyl-CoA carboxylase/reductase: The ethylmalonyl-CoA pathway. Proc. Natl. Acad. Sci. USA 2007, 104, 10631–10636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leroy, B.; De Meur, Q.; Moulin, C.; Wegria, G.; Wattiez, R. New insight into the photoheterotrophic growth of the isocytrate lyase-lacking purple bacterium Rhodospirillum rubrum on acetate. Microbiology 2015, 161, 1061–1072. [Google Scholar] [CrossRef] [PubMed]
- Schneider, K.; Peyraud, R.; Kiefer, P.; Christen, P.; Delmotte, N.; Massou, S.; Portais, J.-C.; Vorholt, J.A. The ethylmalonyl-CoA pathway is used in place of the glyoxylate cycle by Methylobacterium extorquens AM1 during growth on acetate. J. Biol. Chem. 2012, 287, 757–766. [Google Scholar] [CrossRef] [Green Version]
- Knight, M. The photometabolism of propionate by Rhodospirillum rubrum. Biochem. J. 1962, 84, 170. [Google Scholar] [CrossRef] [Green Version]
- De Meur, Q.; Deutschbauer, A.; Koch, M.; Bayon-Vicente, G.; Cabecas Segura, P.; Wattiez, R.; Leroy, B. New perspectives on butyrate assimilation in Rhodospirillum rubrum S1H under photoheterotrophic conditions. BMC Microbiol. 2020, 20, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Cabecas Segura, P.; De Meur, Q.; Alloul, A.; Tanghe, A.; Vlaeminck, S.E.O.R.; Wouwer, V.; Wattiez, R.A.; Dewasme, L.; Leroy, B. Experimental Study and Dynamic Modeling of Preferential Photoassimilation in Volatile Fatty Acid Mixtures by Purple Non-Sulfur Bacteria; Laboratory of Proteomics and Microbiology, University of Mons: Mons, Belgium, 2021; to be Submitt. [Google Scholar]
- Segers, L.; Verstraete, W. Conversion of organic acids to H2 by Rhodospirillaceae grown with glutamate or dinitrogen as nitrogen source. Biotechnol. Bioeng. 1983, 25, 2843–2853. [Google Scholar] [CrossRef]
- Suhaimi, M.; Liessens, J.; Verstraete, W. NH+/4-N assimilation by Rhodobacter capsulatus ATCC 23782 grown axenically and non-axenically in N and C rich media. J. Appl. Bacteriol. 1987, 62, 53–64. [Google Scholar] [CrossRef]
- Snell, K.D.; Feng, F.; Zhong, L.; Martin, D.; Madison, L.L. YfcX enables medium-chain-length poly (3-hydroxyalkanoate) formation from fatty acids in recombinant Escherichia coli fadB strains. J. Bacteriol. 2002, 184, 5696–5705. [Google Scholar] [CrossRef] [Green Version]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Muhitch, M.J. Acetolactate synthase activity in developing maize (Zea mays L.) kernels. Plant Physiol. 1988, 86, 23–27. [Google Scholar] [CrossRef] [PubMed]
- Westerfeld, W.W. A colorimetric determination of blood acetoin. J. Biol. Chem. 1945, 161, 495–502. [Google Scholar] [CrossRef]
- Brock, M.; Maerker, C.; Schütz, A.; Völker, U.; Buckel, W. Oxidation of propionate to pyruvate in Escherichia coli: Involvement of methylcitrate dehydratase and aconitase. Eur. J. Biochem. 2002, 269, 6184–6194. [Google Scholar] [CrossRef]
- Bayon-Vicente, G.; Marchand, E.; Ducrotois, J.; Dufrasne, F.E.; Hallez, R.; Wattiez, R.; Leroy, B. Analysis of the involvement of the isoleucine biosynthesis pathway in photoheterotrophic metabolism of Rhodospirillum rubrum. Front. Microbiol. 2021, 12, 731976. [Google Scholar] [CrossRef]
- Gordon, G.C.; McKinlay, J.B. Calvin cycle mutants of photoheterotrophic purple nonsulfur bacteria fail to grow due to an electron imbalance rather than toxic metabolite accumulation. J. Bacteriol. 2014, 196, 1231–1237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, M.C.; Buchanan, B.B.; Arnon, D.I. A new ferredoxin-dependent carbon reduction cycle in a photosynthetic bacterium. Proc. Natl. Acad. Sci. USA 1966, 55, 928–934. [Google Scholar] [CrossRef] [Green Version]
- Buchanan, B.B.; Arnon, D.I. A reverse KREBS cycle in photosynthesis: Consensus at last. Photosynth. Res. 1990, 24, 47–53. [Google Scholar] [CrossRef]
- Bond-Watts, B.B.; Bellerose, R.J.; Chang, M.C.Y. Enzyme mechanism as a kinetic control element for designing synthetic biofuel pathways. Nat. Chem. Biol. 2011, 7, 222–227. [Google Scholar] [CrossRef] [PubMed]
- Bayon-Vicente, G.; Wattiez, R.; Leroy, B. Global Proteomic Analysis Reveals High Light Intensity Adaptation Strategies and Polyhydroxyalkanoate Production in Rhodospirillum rubrum Cultivated With Acetate as Carbon Source. Front. Microbiol. 2020, 11, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Zhang, Y.; Welch, E.; Li, J.; Roberts, G.P. Elimination of Rubisco alters the regulation of nitrogenase activity and increases hydrogen production in Rhodospirillum rubrum. Int. J. Hydrogen Energy 2010, 35, 7377–7385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brämer, C.O.; Silva, L.F.; Gomez, J.G.C.; Priefert, H.; Steinbüchel, A. Identification of the 2-methylcitrate pathway involved in the catabolism of propionate in the polyhydroxyalkanoate-producing strain Burkholderia sacchari IPT101T and analysis of a mutant accumulating a copolyester with higher 3-hydroxyvalerate content. Appl. Environ. Microbiol. 2002, 68, 271–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, L.F.; Gomez, J.G.C.; Oliveira, M.S.; Torres, B.B. Propionic acid metabolism and poly-3-hydroxybutyrate-co-3-hydroxyvalerate (P3HB-co-3HV) production by Burkholderia sp. J. Biotechnol. 2000, 76, 165–174. [Google Scholar] [CrossRef]
- Hädicke, O.; Grammel, H.; Klamt, S. Metabolic network modeling of redox balancing and biohydrogen production in purple nonsulfur bacteria. BMC Syst. Biol. 2011, 5, 150. [Google Scholar] [CrossRef] [Green Version]
- Ormerod, J.G. The use of radioactive carbon dioxide in the measurement of carbon dioxide fixation in Rhodospirillum rubrum. Biochem. J. 1956, 64, 373. [Google Scholar] [CrossRef] [Green Version]
- De Meur, Q.; Deutschbauer, A.; Koch, M.; Wattiez, R.; Leroy, B. Genetic plasticity and ethylmalonyl coenzyme A pathway during acetate assimilation in Rhodospirillum rubrum S1H under photoheterotrophic conditions. Appl. Environ. Microbiol. 2018, 84, e02038-17. [Google Scholar] [CrossRef] [Green Version]
- Bayon-Vicente, G.; Zarbo, S.; Deutschbauer, A.; Wattiez, R.; Leroy, B. Photoheterotrophic Assimilation of Valerate and Associated Polyhydroxyalkanoate Production by Rhodospirillum rubrum. Appl. Environ. Microbiol. 2020, 86, e00901-20. [Google Scholar] [CrossRef]
- McCully, A.L.; Onyeziri, M.C.; LaSarre, B.; Gliessman, J.R.; McKinlay, J.B. The reverse TCA cycle and reductive amino acid synthesis pathways contribute to electron balance in a Rhodospirillum rubrum Calvin cycle mutant. bioRxiv 2019, 614065. [Google Scholar]

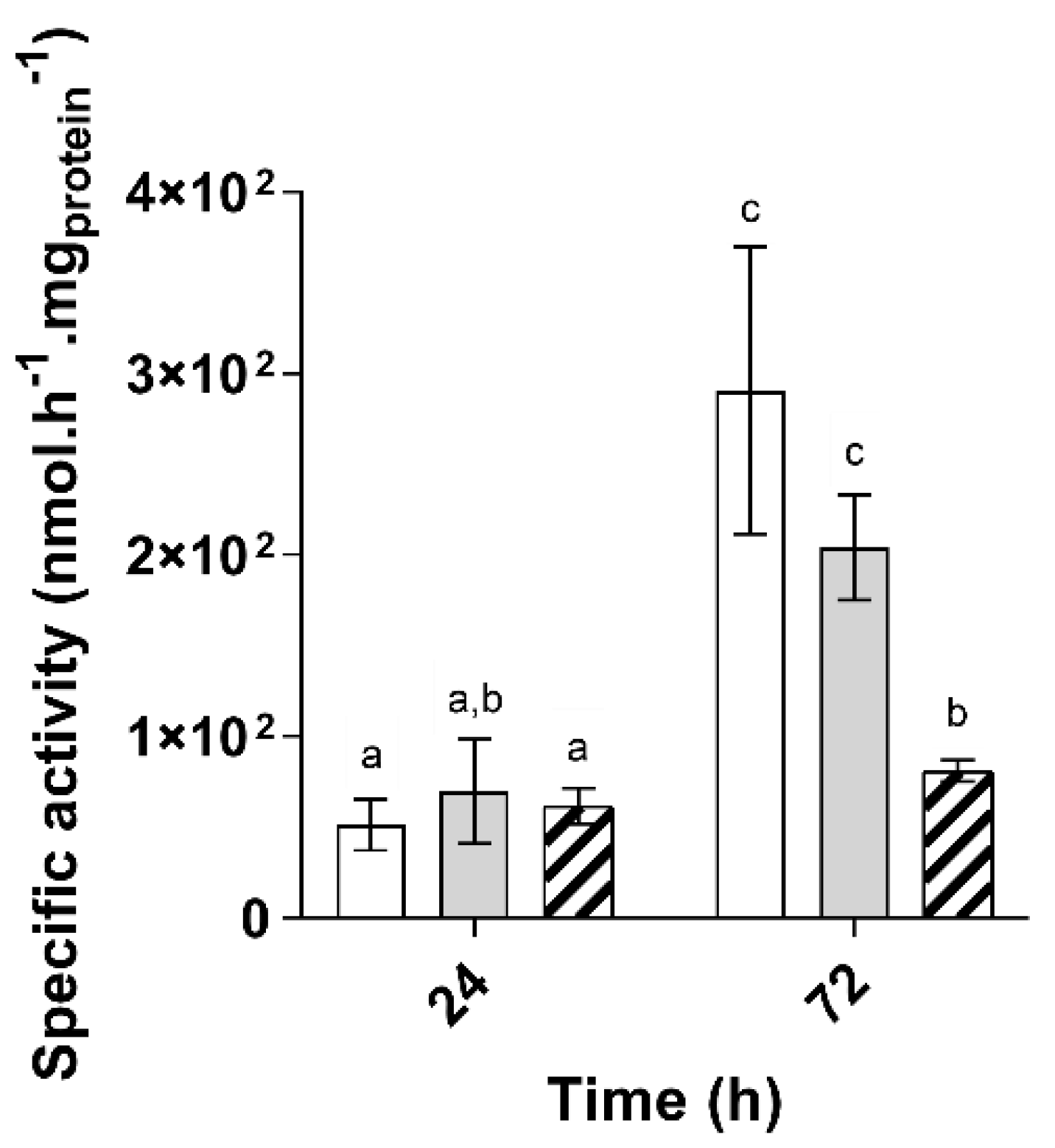
 ) as the carbon source (n = 5). The letters a–d indicate statistically significant difference.
) as the carbon source (n = 5). The letters a–d indicate statistically significant difference.
) as the carbon source (n = 5). The letters a–d indicate statistically significant difference.
) as the carbon source (n = 5). The letters a–d indicate statistically significant difference.

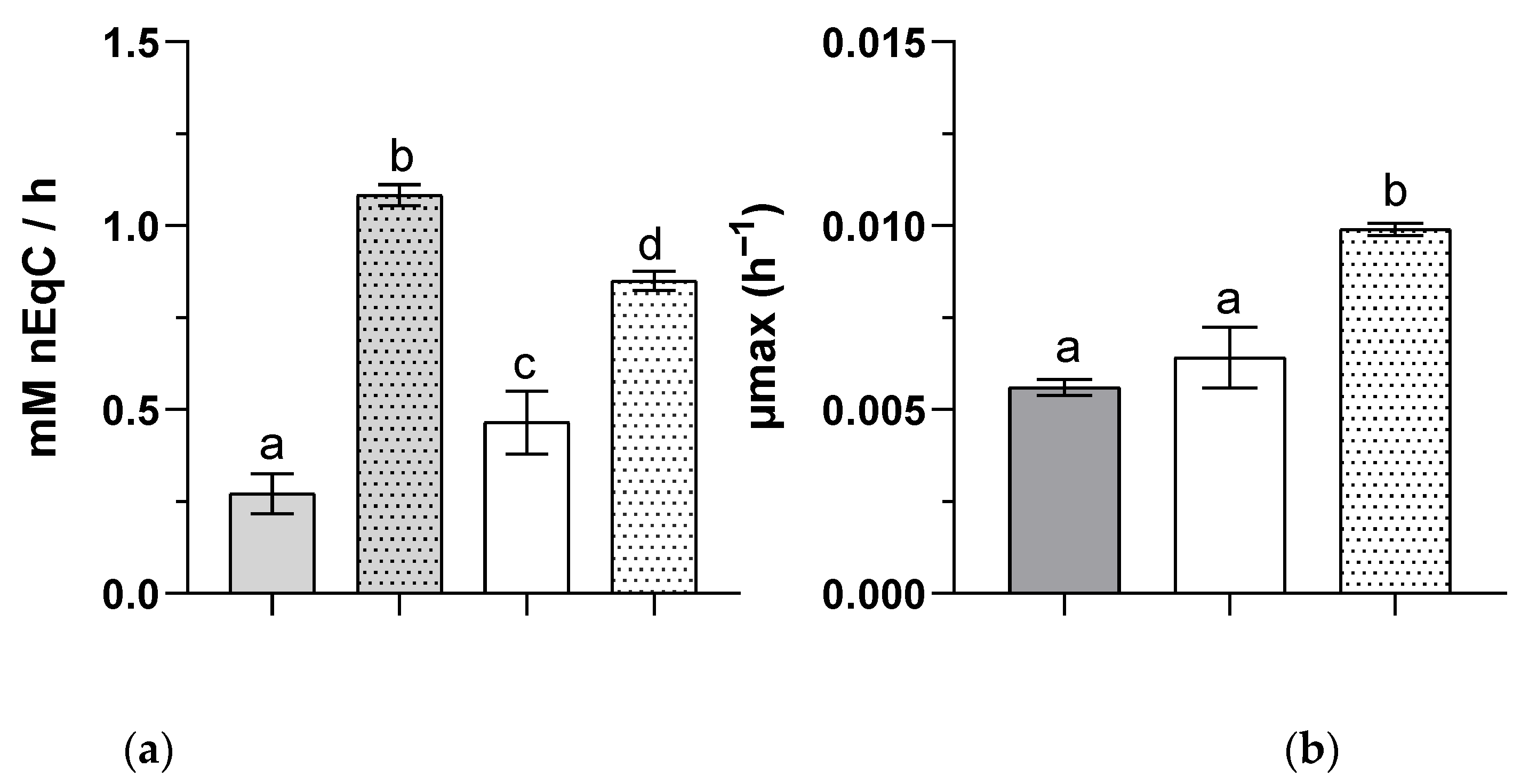
 ) (n = 5). The letters a–c indicate statistically significant difference.
) (n = 5). The letters a–c indicate statistically significant difference.
) (n = 5). The letters a–c indicate statistically significant difference.
) (n = 5). The letters a–c indicate statistically significant difference.
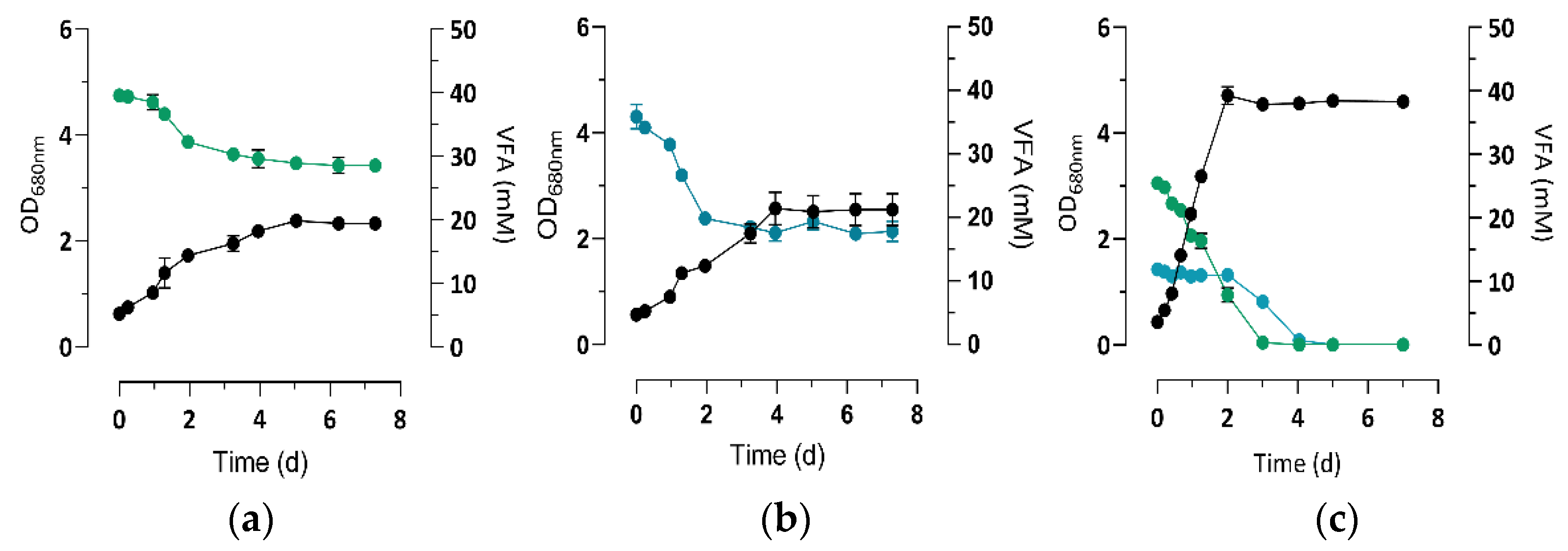
 ) or 3 mM of bicarbonate (
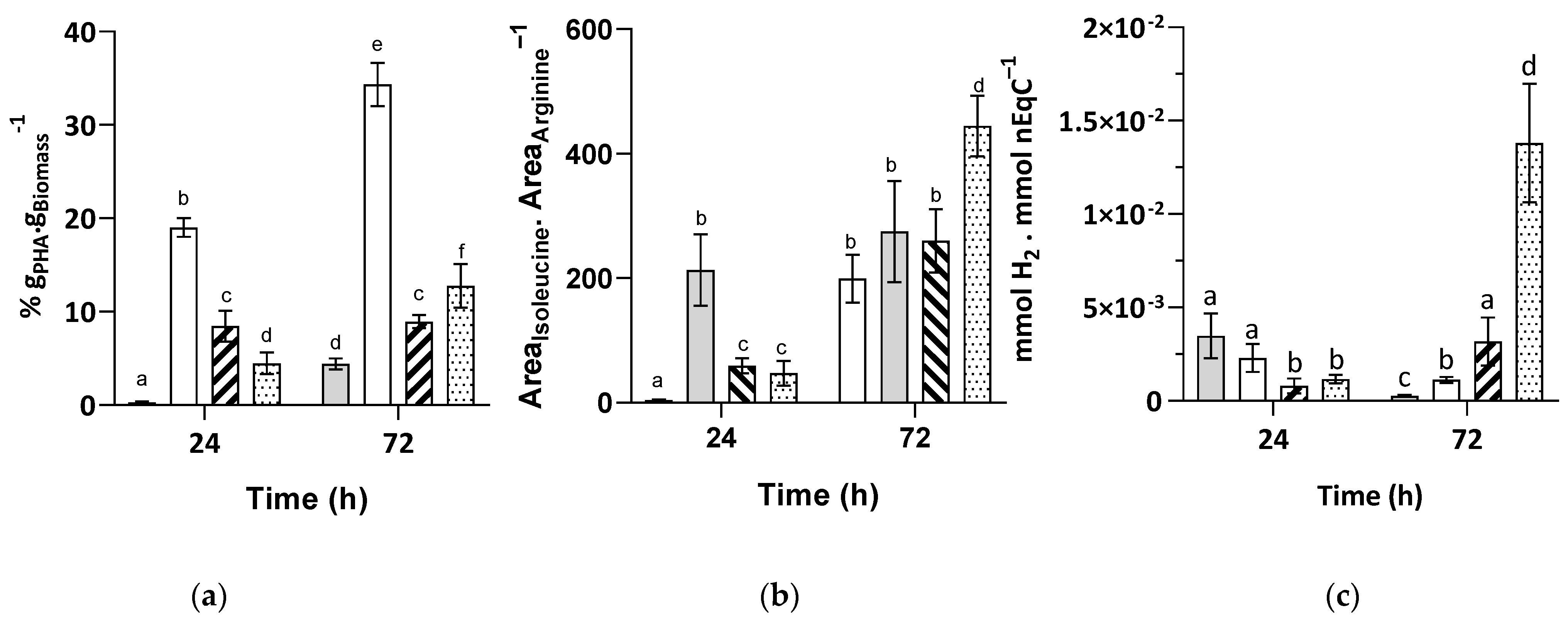
) or 3 mM of bicarbonate (  ) (n = 5). The letters a–f indicate statistically significant difference.
) or 3 mM of bicarbonate ( ) (n = 5). The letters a–f indicate statistically significant difference.
) (n = 5). The letters a–f indicate statistically significant difference.
) or 3 mM of bicarbonate ( ) (n = 5). The letters a–f indicate statistically significant difference.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
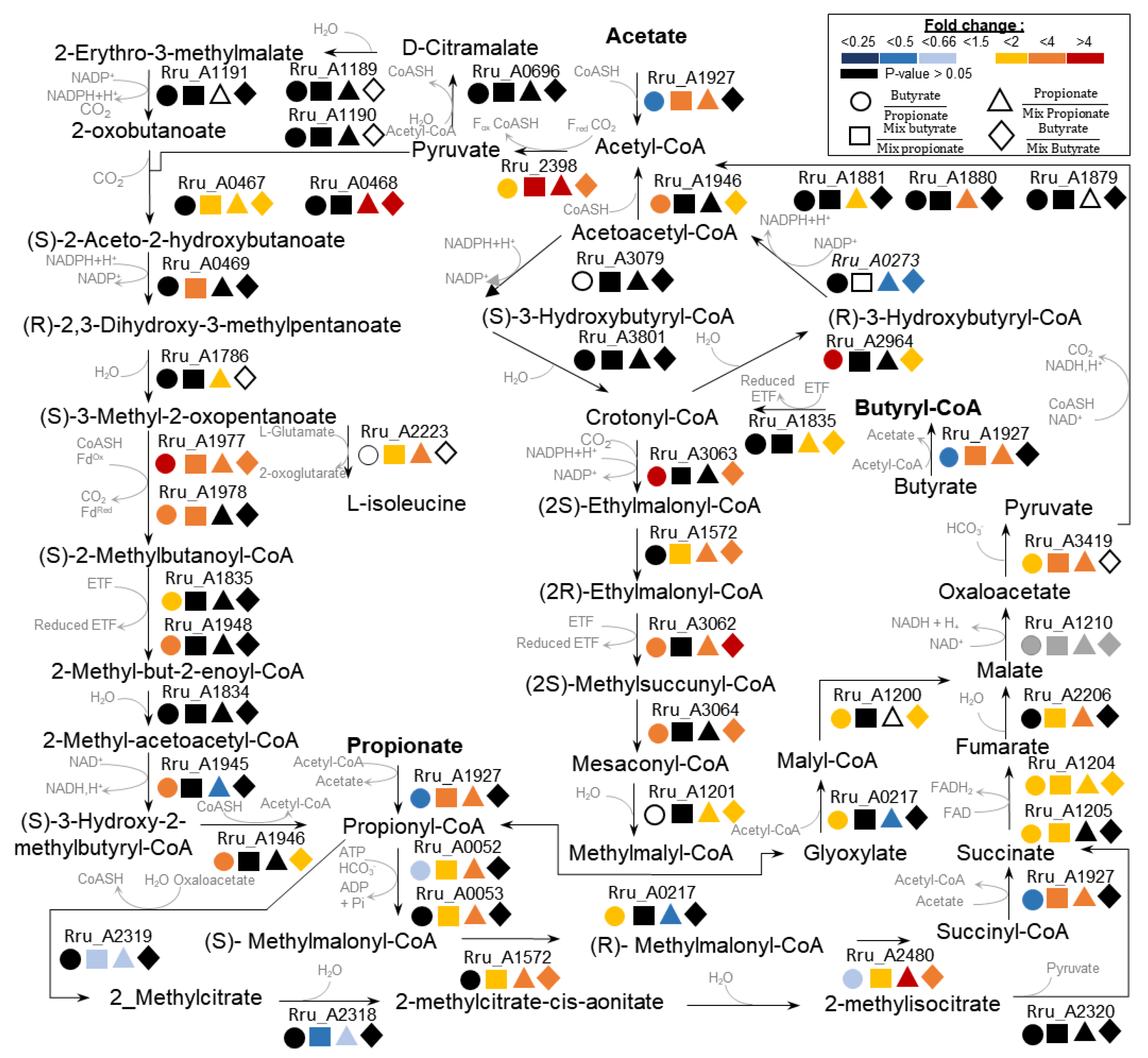
| Uniprot Accession Number | Locus Tag | Fold Change (Butyrate/Propioanate) * | P-Value | Fold Change (Mix Butyrate/Mix Propionate) ** | P-Value | No. of Identified Peptides *** | Description |
|---|---|---|---|---|---|---|---|
| Methylmalonyl-CoA pathway | |||||||
| tr|Q2RYD8 | Rru_A0052 | 0.63 | 3.41 × 10−2 | 1.81 | 9.50 × 10−3 | 5 | Biotin carboxylase |
| tr|Q2RYD7 | Rru_A0053 | 0.83 | 8.70 × 10−2 | 1.81 | 4.95 × 10−2 | 6 | Carboxyl transferase |
| tr|Q2RU23 | Rru_A1572 | 1.30 | 3.49 × 10−1 | 1.67 | 1.00 × 10−2 | 5 | Methylmalonyl-CoA epimerase |
| tr|Q2RRG6 | Rru_A2479 | 0.72 | 8.21 × 10−2 | 1.75 | 9.56 × 10−2 | 6 | Methylmalonyl-CoA mutase |
| tr|Q2RRG5 | Rru_A2480 | 0.61 | 3.43 × 10−2 | 1.56 | 1.37 × 10−3 | 6 | Methylmalonyl-CoA mutase |
| Methilcitrate cycle | |||||||
| tr|Q2RRX7 | Rru_A2318 | 0.76 | 4.60 × 10−1 | 0.47 | 1.22 × 10−2 | 4 | 2-methylcitrate dehydratase |
| tr|Q2RRX6 | Rru_A2319 | 0.25 | 1.61 × 10−1 | 0.53 | 3.96 × 10−2 | 5 | Citrate synthase |
| tr|Q2RRX5 | Rru_A2320 | 1.07 | 8.75 × 10−1 | 0.66 | 3.42 × 10−1 | 2 | 2.3-dimethylmalate lyase |
| Ethylmalonyl-CoA pathway | |||||||
| tr|Q2RXX3 | Rru_A0217 | 1.97 | 1.38 × 10−2 | 0.72 | 4.46 × 10−1 | 6 | Citrate lyase |
| tr|Q2RXR7 | Rru_A0273 | 0.556 | 1.00 × 10−1 | 0.74 | 7.70 × 10−3 | 5 | 3-oxoacyl-eductase |
| tr|Q2RV43 | Rru_A1201 | 1.48 | 8.73 × 10−4 | 1.34 | 3.48 × 10−1 | 6 | MaoC-like dehydratase |
| tr|Q2RTB0 | Rru_A1835 | 1.97 | 1.43 × 10−2 | 1.49 | 1.41 × 10−1 | 4 | Butyryl-CoA dehydrogenase |
| tr|Q2RT18 | Rru_A1927 | 0.35 | 1.60 × 10−2 | 2.33 | 9.63 × 10−3 | 6 | Acetyl-CoA hydrolase |
| tr|Q2RPS1 | Rru_A3079 | 1.49 | 7.00 × 10−3 | 1.05 | 3.24 × 10−1 | 6 | 3-hydroxyacyl-CoA dehydrogenase |
| tr|Q2RQ36 | Rru_A2964 | 1.03 | 8.16 × 10−1 | 0.87 | 3.28 × 10−1 | 2 | MaoC-like dehydratase |
| tr|Q2RPT8 | Rru_A3062 | 2.62 | 2.59 × 10−2 | 1.59 | 1.68 × 10−1 | 5 | Methylmalonyl-CoA mutase |
| tr|Q2RPT7 | Rru_A3063 | 6.17 | 4.78 × 10−2 | 0.76 | 2.34 × 10−1 | 6 | Crotonyl-CoA reductase |
| tr|Q2RPT6 | Rru_A3064 | 2.23 | 4.96 × 10−2 | 1.22 | 5.03 × 10−1 | 6 | Isovaleryl-CoA dehydrogenase |
| tr|Q2RSZ9 | Rru_A1946 | 2.63 | 5.35 × 10−3 | 1.18 | 3.40 × 10−1 | 5 | Acetyl-CoA C-acetyltransferase |
| tr|Q2RMQ0 | Rru_A3801 | 1.05 | 5.90 × 10−1 | 0.93 | 6.26 × 10−1 | 6 | Short chain enoyl-CoA hydratase |
| Methylbutanoyl-CoA pathway | |||||||
| tr|Q2RX73 | Rru_A0467 | 1.25 | 3.69 × 10−1 | 1.66 | 1.51 × 10−2 | 6 | Acetolactate synthase. large subunit |
| tr|Q2RX72 | Rru_A0468 | 0.99 | 8.96 × 10−1 | 1.05 | 7.30 × 10−1 | 3 | Acetolactate synthase. small subunit |
| sp|Q2RX71 | Rru_A0469 | 1.58 | 6.27 × 10−2 | 2.02 | 4.40 × 10−3 | 6 | Ketol-acid reductoisomerase |
| tr|Q2RX33 | Rru_A0508 | 1.40 | 4.91 × 10−1 | 1.07 | 9.34 × 10−1 | 1 | Aminotransferase |
| tr|Q2RWJ5 | Rru_A0696 | 0.47 | 9.37 × 10−2 | 1.10 | 8.65 × 10−1 | 1 | RNA methyltransferase |
| sp|Q2RV55 | Rru_A1189 | 0.92 | 4.49 × 10−1 | 1.12 | 8.59 × 10−2 | 5 | 3-isopropylmalate dehydratase large subunit |
| sp|Q2RV54 | Rru_A1190 | 1.06 | 5.53 × 10−1 | 1.49 | 2.54 × 10−1 | 6 | 3-isopropylmalate dehydratase small subunit |
| sp|Q2RV53 | Rru_A1191 | 0.83 | 8.25 × 10−2 | 0.87 | 1.96 × 10−1 | 6 | 3-isopropylmalate dehydrogenase |
| sp|Q2RTF9 | Rru_A1786 | 0.92 | 4.73 × 10−1 | 1.04 | 6.44 × 10−1 | 6 | Dihydroxy-acid dehydratase |
| tr|Q2RTB1 | Rru_A1834 | 1.16 | 4.20 × 10−1 | 1.26 | 9.54 × 10−2 | 2 | Enoyl-CoA hydratase/isomerase |
| tr|Q2RT00 | Rru_A1945 | 2.17 | 4.24 × 10−3 | 1.11 | 5.24 × 10−1 | 5 | Short-chain dehydrogenase/reductase |
| tr|Q2RSZ7 | Rru_A1948 | 3.14 | 3.47 × 10−2 | 0.91 | 7.65 × 10−1 | 2 | Isovaleryl-CoA dehydrogenase |
| tr|Q2RSW8 | Rru_A1977 | 4.86 | 3.21 × 10−2 | 3.13 | 1.81 × 10−2 | 2 | Pyruvate ferredoxin/flavodoxin oxidoreductase |
| tr|Q2RSW7 | Rru_A1978 | 3.32 | 2.41 × 10−2 | 2.02 | 4.90 × 10−2 | 6 | Indolepyruvate ferredoxin oxidoreductase |
| tr|Q2RS72 | Rru_A2223 | 0.79 | 2.10 × 10−1 | 1.87 | 4.90 × 10−2 | 6 | 2-keto-4-methylthiobutyrate aminotransferase |
| sp|Q53046 | Rru_A2398 | 1.79 | 4.72 × 10−2 | 11.31 | 4.22 × 10−2 | 6 | Pyruvate-flavodoxin oxidoreductase |
| Tricarboxylic carbon cycle | |||||||
| tr|Q2RV44 | Rru_A1200 | 1.87 | 2.59 × 10−3 | 1.53 | 1.06 × 10−1 | 5 | Citrate lyase |
| tr|Q2RV40 | Rru_A1204 | 1.82 | 1.58 × 10−2 | 1.55 | 4.84 × 10−3 | 6 | MaoC-like dehydratase |
| tr|Q2RV39 | Rru_A1205 | 1.98 | 3.64 × 10−2 | 1.54 | 4.01 × 10−2 | 5 | MaoC-like dehydratase |
| tr|Q2RT66 | Rru_A1879 | 0.82 | 2.98 × 10−1 | 1.03 | 7.28 × 10−1 | 6 | Dihydrolipoamide acetyltransferase |
| tr|Q2RT65 | Rru_A1880 | 0.85 | 3.22 × 10−1 | 1.08 | 3.59 × 10−1 | 6 | Pyruvate dehydrogenase beta subunit |
| tr|Q2RT64 | Rru_A1881 | 1.00 | 9.92 × 10−1 | 0.98 | 9.21 × 10−1 | 4 | Pyruvate dehydrogenase |
| tr|Q2RS89 | Rru_A2206 | 0.79 | 1.46 × 10−1 | 1.72 | 1.14 × 10−3 | 6 | Fumarase |
| tr|Q2RNT2 | Rru_A3419 | 1.88 | 6.88 × 10−3 | 3.02 | 1.35 × 10−3 | 6 | Phosphoenolpyruvate carboxykinase |
| Calvin-Benson-Bassham cycle | |||||||
| sp|Q2RRP5 | Rru_A2400 | 4.20 | 3.45 × 10−2 | 2.68 | 1.86 × 10−1 | 6 | Ribulose bisphosphate carboxylase |
| Reverse Tricarboxylic carbon cycle | |||||||
| tr|Q2RQS7 | Rru_A2721 | 0.96 | 9.26 × 10−1 | 1.52 | 1.48 × 10−1 | 2 | 2-oxoglutarate synthase. alpha subunit |
| tr|Q2RQS6 | Rru_A2722 | 0.99 | 9.75 × 10−1 | 1.50 | 2.75 × 10−2 | 5 | 2-oxoglutarate synthase, beta subunit |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cabecas Segura, P.; De Meur, Q.; Tanghe, A.; Onderwater, R.; Dewasme, L.; Wattiez, R.; Leroy, B. Effects of Mixing Volatile Fatty Acids as Carbon Sources on Rhodospirillum rubrum Carbon Metabolism and Redox Balance Mechanisms. Microorganisms 2021, 9, 1996. https://doi.org/10.3390/microorganisms9091996
Cabecas Segura P, De Meur Q, Tanghe A, Onderwater R, Dewasme L, Wattiez R, Leroy B. Effects of Mixing Volatile Fatty Acids as Carbon Sources on Rhodospirillum rubrum Carbon Metabolism and Redox Balance Mechanisms. Microorganisms. 2021; 9(9):1996. https://doi.org/10.3390/microorganisms9091996
Chicago/Turabian StyleCabecas Segura, Paloma, Quentin De Meur, Audrey Tanghe, Rob Onderwater, Laurent Dewasme, Ruddy Wattiez, and Baptiste Leroy. 2021. "Effects of Mixing Volatile Fatty Acids as Carbon Sources on Rhodospirillum rubrum Carbon Metabolism and Redox Balance Mechanisms" Microorganisms 9, no. 9: 1996. https://doi.org/10.3390/microorganisms9091996