
Unique Tubulin-Based Structures in the Zoonotic Apicomplexan Parasite Cryptosporidium parvum


Abstract
:1. Introduction
2. Materials and Methods
2.1. Antibody Development and Affinity-Purification
2.2. Preparation of Parasite Materials
2.3. qRT-PCR Analysis of CpTubA and CpTubB Gene Transcripts
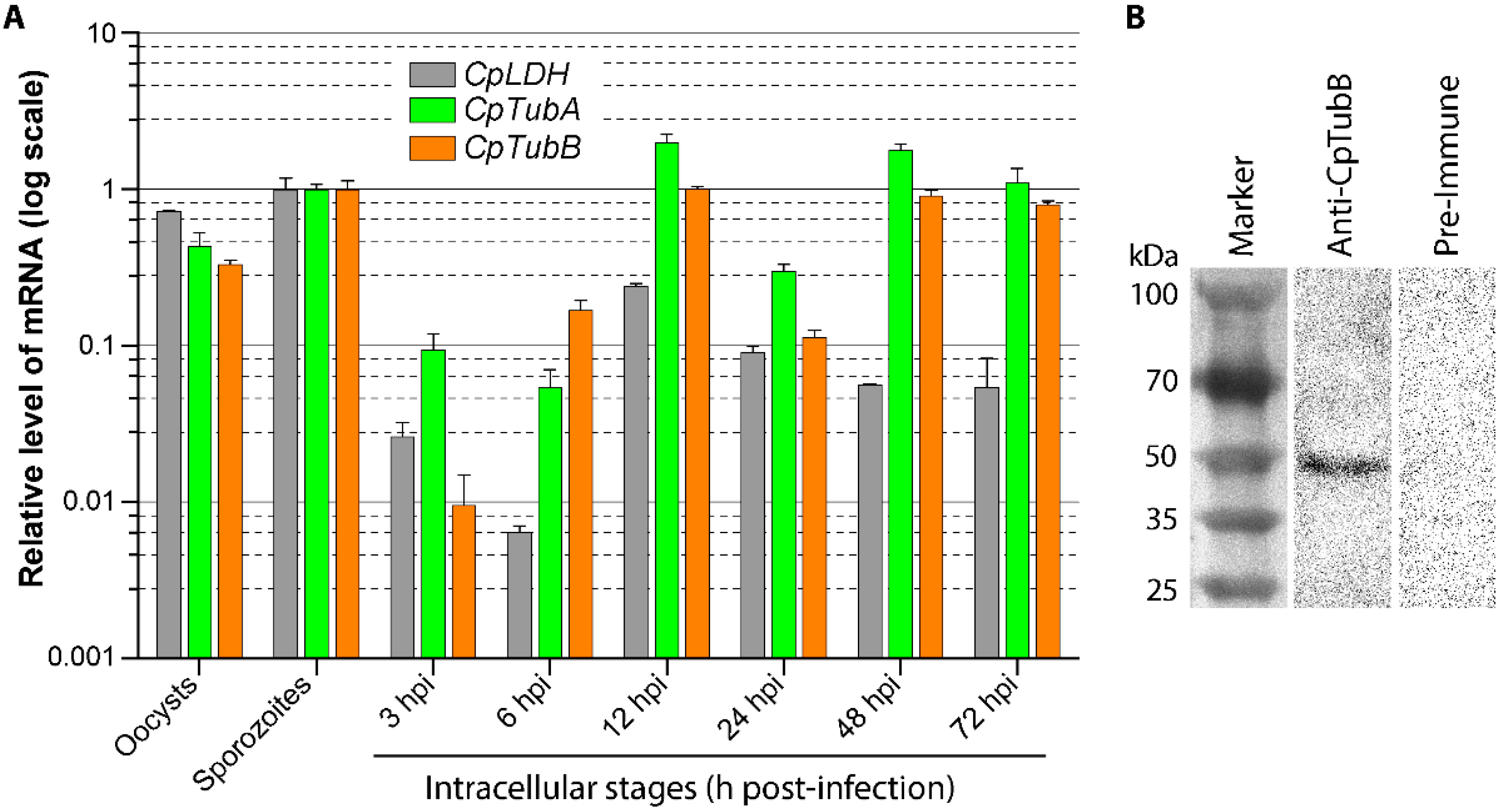
2.4. Western Blot Analysis
2.5. Indirect Immunofluorescence Assay (IFA)
2.6. Phylogenetic Analysis of Apicomplexan α-, β- and γ-tubulin Proteins
3. Results and Discussion
3.1. The α- and β-tubulin Genes in C. Parvum Are Differentially Expressed during the Parasite Life Cycle
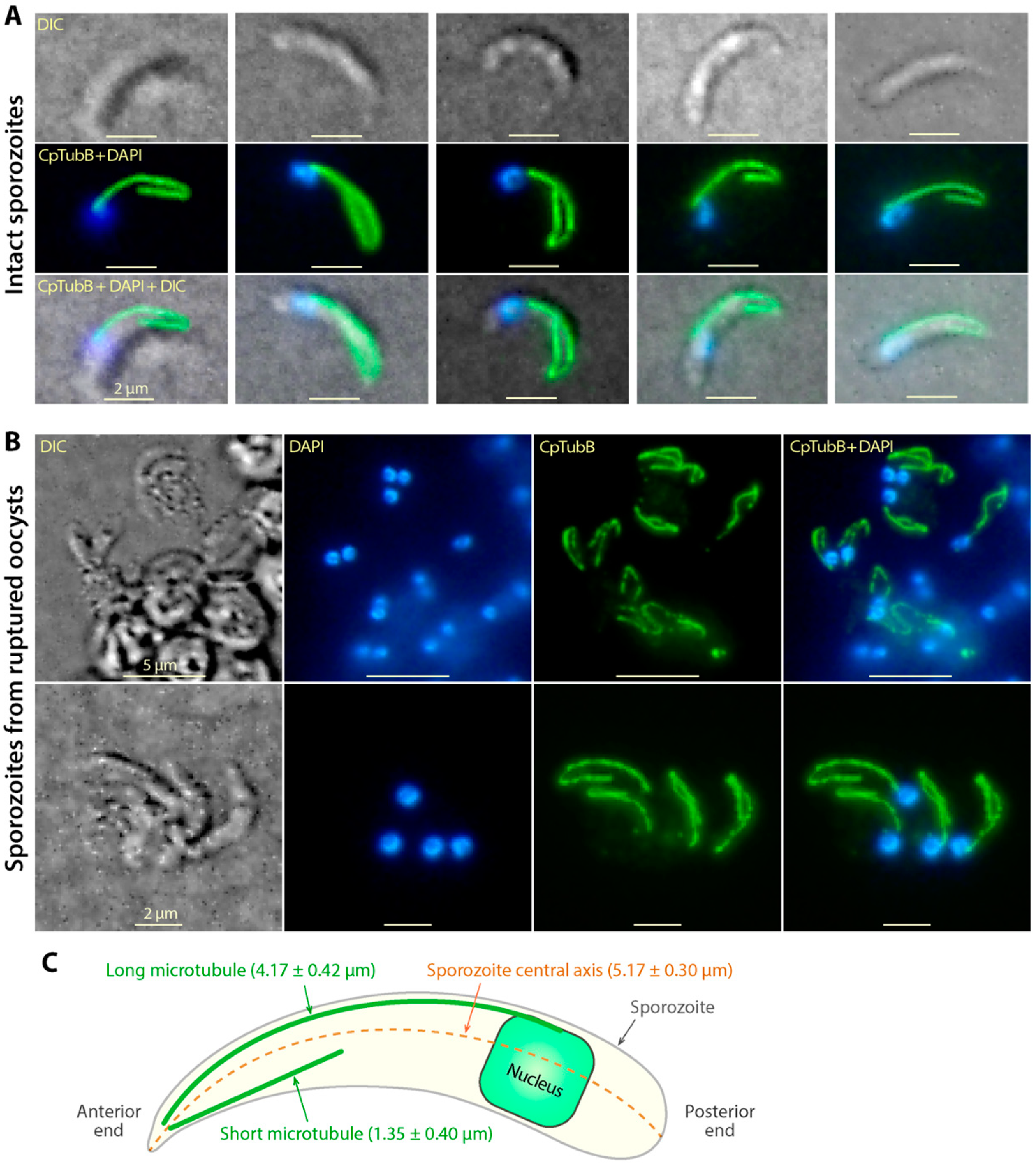
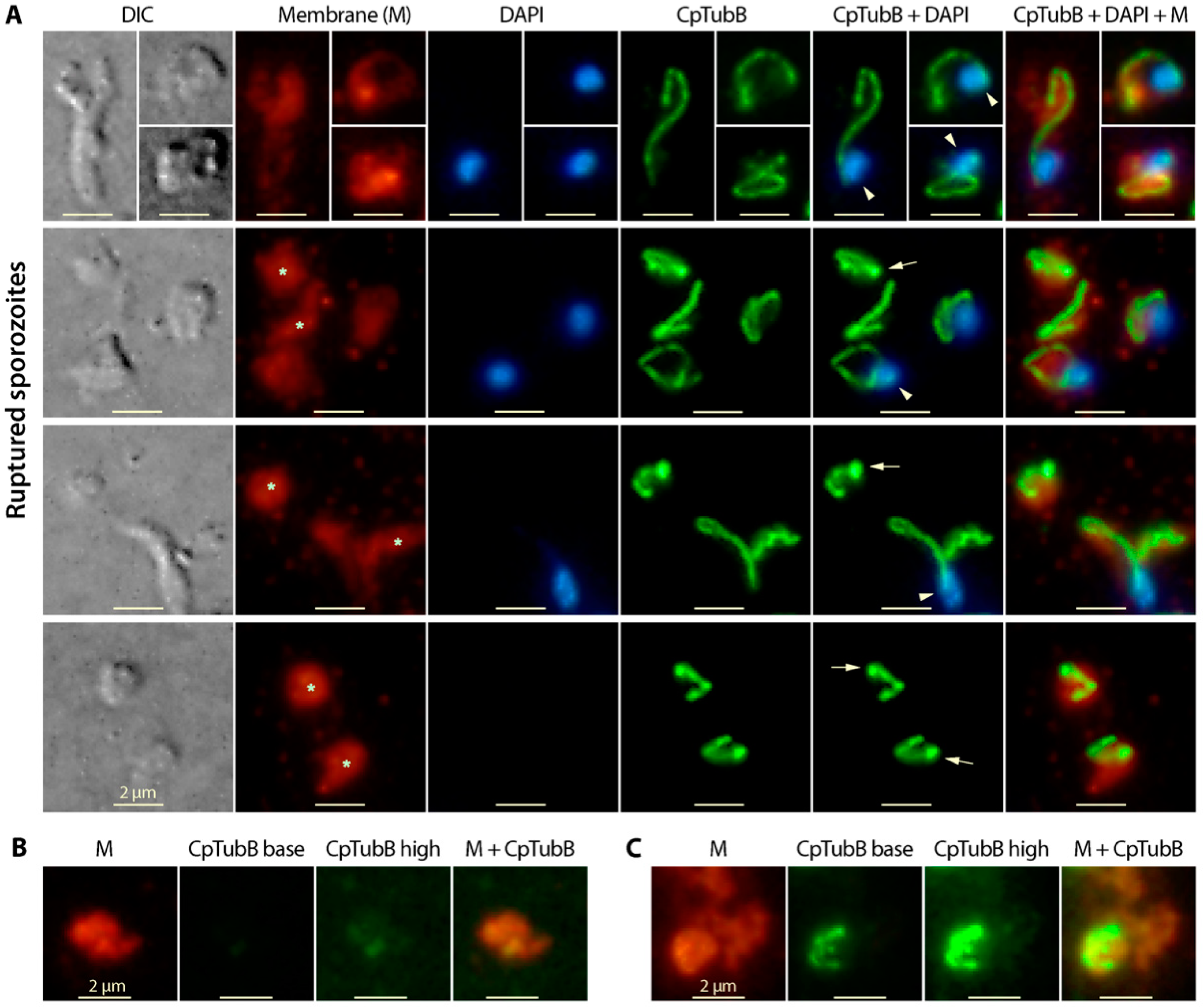
3.2. Tubulin Polymers Form Two Microtubular Filaments in the C. parvum Sporozoites Differing from Those in Other Apicomplexans
3.3. The Two C. Parvum Microtubules in the Sporozoite Stage Are Rigid, and the Long Microtubule Appeared to Be Associated with the Nucleus
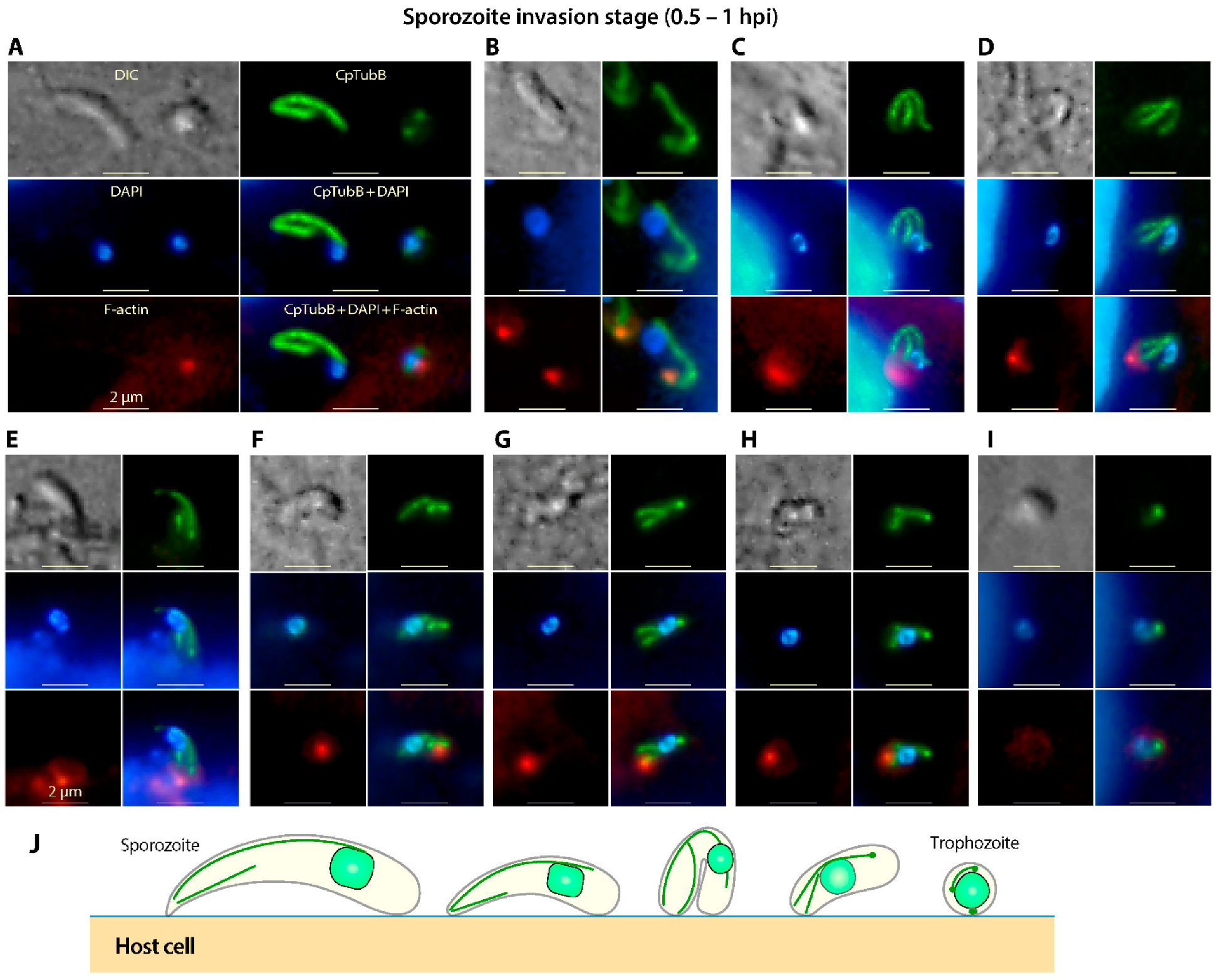
3.4. The Two Microtubules in C. Parvum Are Restructured during the Invasion of Sporozoites and Its Transformation into Trophozoites
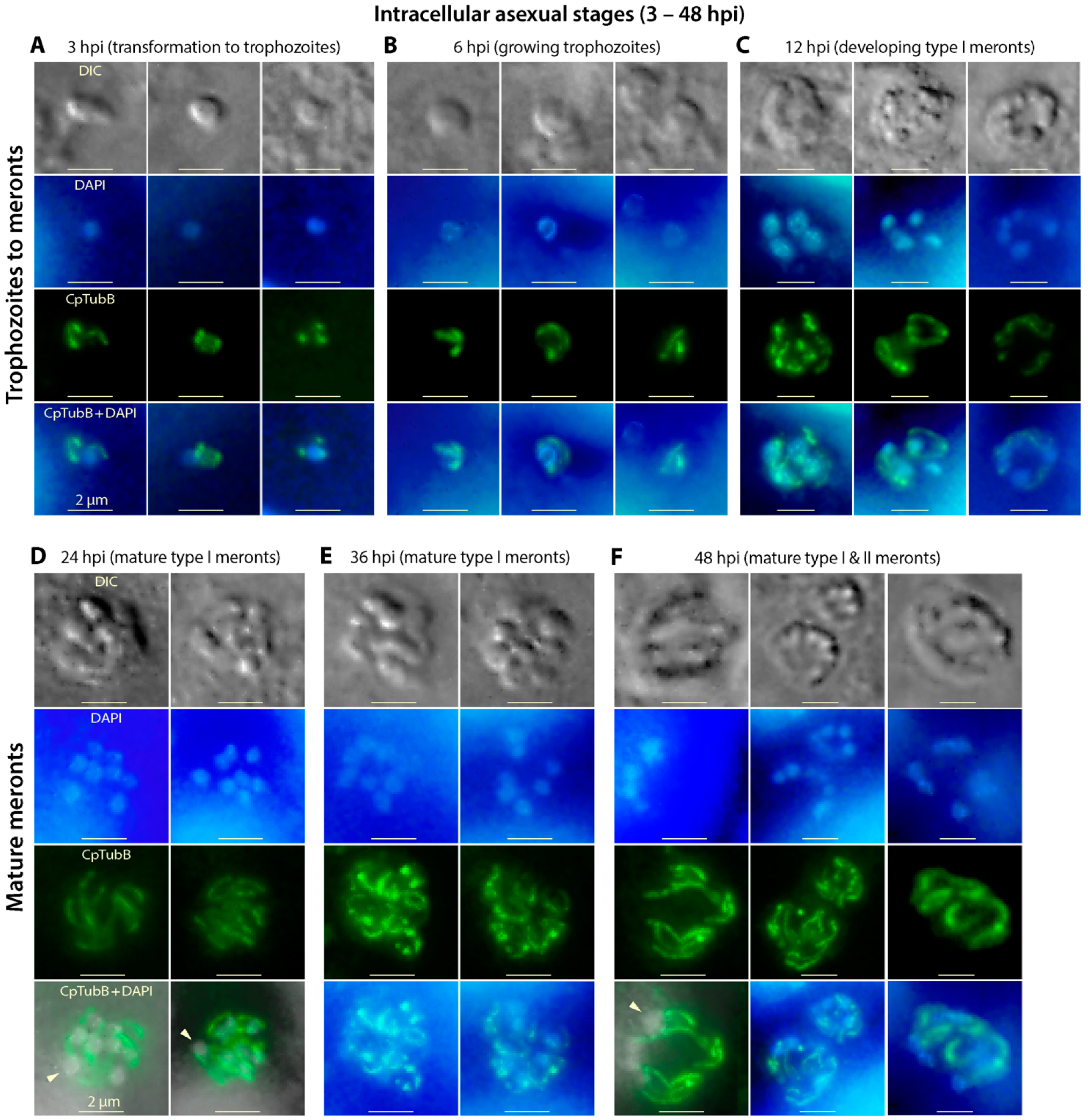
3.5. The Two Microtubules of C. parvum Are Present in the Asexual Developmental Stages (Merogony) and Rearranged during Parasite Development
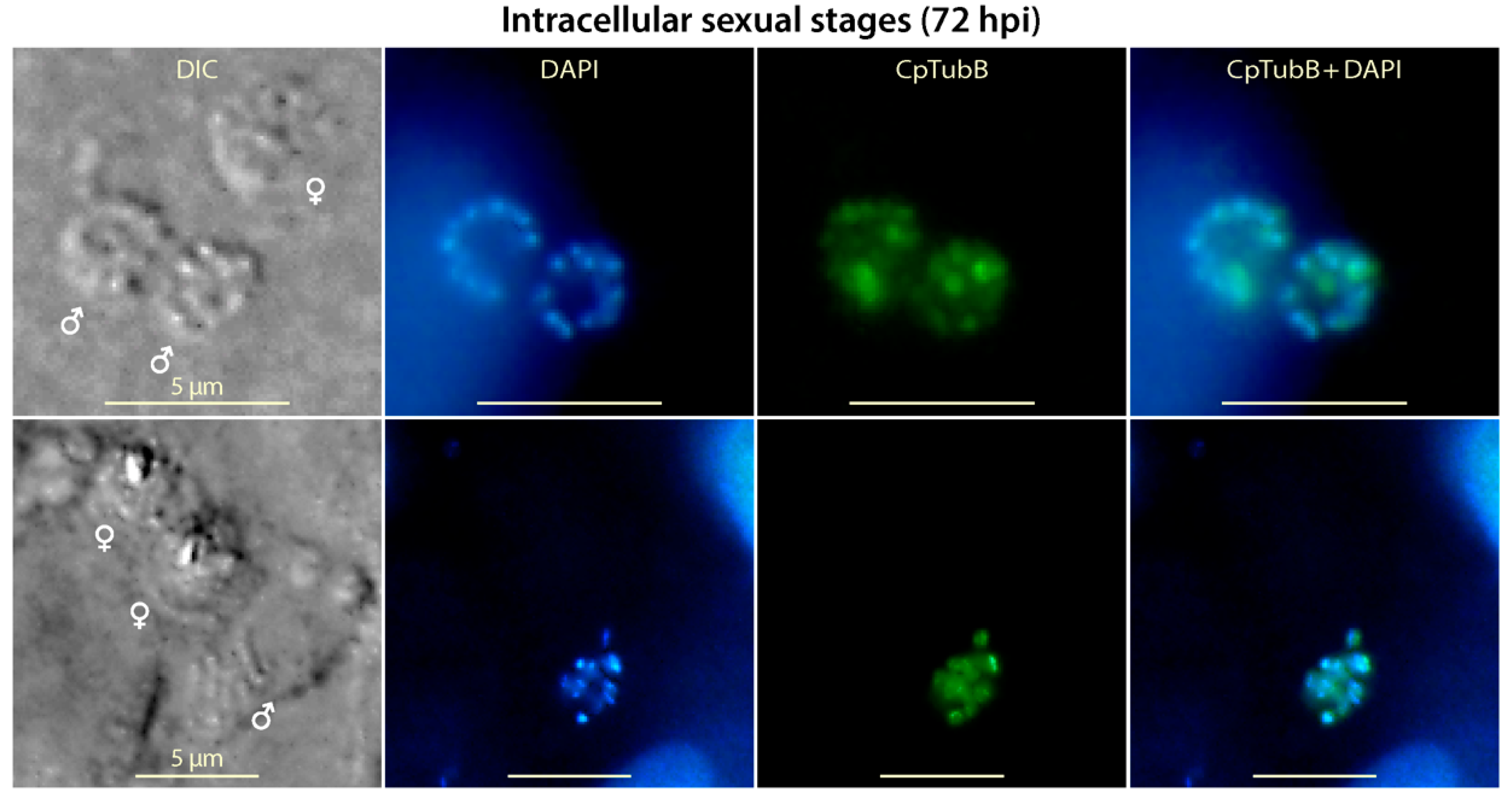
3.6. Tubulin-Based Structures Were Present in Microgametes, but Appear Absent in Macrogametes in the Sexual Development of C. parvum
3.7. Lack of Other α- and β-tubulin Orthologs for Alternative Microtubules in the Cryptosporidium Genomes
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Goodson, H.V.; Jonasson, E.M. Microtubules and Microtubule-Associated Proteins. Cold Spring Harb Perspect. Biol. 2018, 10, a022608. [Google Scholar] [CrossRef]
- Logan, C.M.; Menko, A.S. Microtubules: Evolving roles and critical cellular interactions. Exp. Biol. Med. (Maywood) 2019, 244, 1240–1254. [Google Scholar] [CrossRef]
- Tosetti, N.; Dos Santos Pacheco, N.; Bertiaux, E.; Maco, B.; Bournonville, L.; Hamel, V.; Guichard, P.; Soldati-Favre, D. Essential function of the alveolin network in the subpellicular microtubules and conoid assembly in Toxoplasma gondii. Elife 2020, 9, e56635. [Google Scholar] [CrossRef] [PubMed]
- Harding, C.R.; Frischknecht, F. The Riveting Cellular Structures of Apicomplexan Parasites. Trends Parasitol. 2020, 36, 979–991. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos Pacheco, N.; Tosetti, N.; Koreny, L.; Waller, R.F.; Soldati-Favre, D. Evolution, Composition, Assembly, and Function of the Conoid in Apicomplexa. Trends Parasitol. 2020, 36, 688–704. [Google Scholar] [CrossRef]
- Morrissette, N.S.; Sibley, L.D. Cytoskeleton of apicomplexan parasites. Microbiol. Mol. Biol. Rev. 2002, 66, 21–38. [Google Scholar] [CrossRef] [Green Version]
- Frenal, K.; Dubremetz, J.F.; Lebrun, M.; Soldati-Favre, D. Gliding motility powers invasion and egress in Apicomplexa. Nat. Rev. Microbiol. 2017, 15, 645–660. [Google Scholar] [CrossRef] [Green Version]
- Zhu, G.; Yin, J.; Cuny, G.D. Current status and challenges in drug discovery against the globally important zoonotic cryptosporidiosis. Anim. Dis. 2021, 1, 3. [Google Scholar] [CrossRef]
- Pinto, D.J.; Vinayak, S. Cryptosporidium: Host-Parasite Interactions and Pathogenesis. Curr. Clin. Microbiol. Rep. 2021, 8, 62–67. [Google Scholar] [CrossRef] [PubMed]
- Innes, E.A.; Chalmers, R.M.; Wells, B.; Pawlowic, M.C. A One Health Approach to Tackle Cryptosporidiosis. Trends Parasitol. 2020, 36, 290–303. [Google Scholar] [CrossRef] [Green Version]
- Ryan, U.; Zahedi, A.; Paparini, A. Cryptosporidium in humans and animals-a one health approach to prophylaxis. Parasite Immunol. 2016, 38, 535–547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uni, S.; Iseki, M.; Maekawa, T.; Moriya, K.; Takada, S. Ultrastructure of Cryptosporidium muris (strain RN 66) parasitizing the murine stomach. Parasitol. Res. 1987, 74, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Matsubayashi, M.; Takase, H.; Kimata, I.; Nakagawa, H.; Tani, H.; Sasai, K.; Baba, E. Electron microscopic observation of cytoskeletal frame structures and detection of tubulin on the apical region of Cryptosporidium parvum sporozoites. Parasitology 2008, 135, 295–301. [Google Scholar] [CrossRef]
- O’Hara, S.P.; Huang, B.Q.; Chen, X.M.; Nelson, J.; LaRusso, N.F. Distribution of Cryptosporidium parvum sporozoite apical organelles during attachment to and internalization by cultured biliary epithelial cells. J. Parasitol. 2005, 91, 995–999. [Google Scholar] [CrossRef]
- Guo, F.; Zhang, H.; McNair, N.N.; Mead, J.R.; Zhu, G. The Existing Drug Vorinostat as a New Lead Against Cryptosporidiosis by Targeting the Parasite Histone Deacetylases. J. Infect. Dis. 2018, 217, 1110–1117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lateef, S.S.; Gupta, S.; Jayathilaka, L.P.; Krishnanchettiar, S.; Huang, J.S.; Lee, B.S. An improved protocol for coupling synthetic peptides to carrier proteins for antibody production using DMF to solubilize peptides. J. Biomol. Tech. 2007, 18, 173–176. [Google Scholar] [PubMed]
- Greenfield, E.A. Standard Immunization of Rabbits. Cold Spring Harb Protoc. 2020, 2020, 100305. [Google Scholar] [CrossRef]
- Kurien, B.T. Affinity purification of autoantibodies from an antigen strip excised from a nitrocellulose protein blot. Methods Mol. Biol. 2009, 536, 201–211. [Google Scholar] [CrossRef]
- Truong, Q.; Ferrari, B.C. Quantitative and qualitative comparisons of Cryptosporidium faecal purification procedures for the isolation of oocysts suitable for proteomic analysis. Int. J. Parasitol. 2006, 36, 811–819. [Google Scholar] [CrossRef]
- Zhang, H.; Zhu, G. High-Throughput Screening of Drugs against the Growth of Cryptosporidium parvum In Vitro by qRT-PCR. Methods Mol. Biol. 2020, 2052, 319–334. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Guo, F.; Zhu, G. Involvement of host cell integrin alpha2 in Cryptosporidium parvum infection. Infect. Immun. 2012, 80, 1753–1758. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Zhu, G. Quantitative RT-PCR assay for high-throughput screening (HTS) of drugs against the growth of Cryptosporidium parvum in vitro. Front. Microbiol. 2015, 6, 991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mauzy, M.J.; Enomoto, S.; Lancto, C.A.; Abrahamsen, M.S.; Rutherford, M.S. The Cryptosporidium parvum transcriptome during in vitro development. PLoS ONE 2012, 7, e31715. [Google Scholar] [CrossRef] [Green Version]
- Guo, F.; Zhang, H.; Payne, H.R.; Zhu, G. Differential Gene Expression and Protein Localization of Cryptosporidium parvum Fatty Acyl-CoA Synthetase Isoforms. J. Eukaryot. Microbiol. 2016, 63, 233–246. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Guo, F.; Zhu, G. Cryptosporidium Lactate Dehydrogenase Is Associated with the Parasitophorous Vacuole Membrane and Is a Potential Target for Developing Therapeutics. PLoS Pathog. 2015, 11, e1005250. [Google Scholar] [CrossRef]
- Zhang, H.; Guo, F.; Zhou, H.; Zhu, G. Transcriptome analysis reveals unique metabolic features in the Cryptosporidium parvum Oocysts associated with environmental survival and stresses. BMC Genom. 2012, 13, 647. [Google Scholar] [CrossRef] [Green Version]
- Hu, K.; Roos, D.S.; Murray, J.M. A novel polymer of tubulin forms the conoid of Toxoplasma gondii. J. Cell Biol. 2002, 156, 1039–1050. [Google Scholar] [CrossRef] [Green Version]
- Xiao, H.; El Bissati, K.; Verdier-Pinard, P.; Burd, B.; Zhang, H.; Kim, K.; Fiser, A.; Angeletti, R.H.; Weiss, L.M. Post-translational modifications to Toxoplasma gondii alpha- and beta-tubulins include novel C-terminal methylation. J. Proteome Res. 2010, 9, 359–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guerin, A.; Roy, N.H.; Kugler, E.M.; Berry, L.; Burkhardt, J.K.; Shin, J.B.; Striepen, B. Cryptosporidium rhoptry effector protein ROP1 injected during invasion targets the host cytoskeletal modulator LMO7. Cell Host Microbe 2021, 29, 1407–1420.e5. [Google Scholar] [CrossRef] [PubMed]
- Stephens, D.J. Functional coupling of microtubules to membranes—Implications for membrane structure and dynamics. J. Cell Sci. 2012, 125, 2795–2804. [Google Scholar] [CrossRef] [Green Version]
- Barlan, K.; Gelfand, V.I. Microtubule-Based Transport and the Distribution, Tethering, and Organization of Organelles. Cold Spring Harb Perspect Biol. 2017, 9, a025817. [Google Scholar] [CrossRef]
- Gallardo, P.; Barrales, R.R.; Daga, R.R.; Salas-Pino, S. Nuclear Mechanics in the Fission Yeast. Cells 2019, 8, 1285. [Google Scholar] [CrossRef] [Green Version]
- Janota, C.S.; Calero-Cuenca, F.J.; Gomes, E.R. The role of the cell nucleus in mechanotransduction. Curr. Opin. Cell Biol. 2020, 63, 204–211. [Google Scholar] [CrossRef] [PubMed]
- Francia, M.E.; Striepen, B. Cell division in apicomplexan parasites. Nat. Rev. Microbiol. 2014, 12, 125–136. [Google Scholar] [CrossRef] [PubMed]
- Tandel, J.; English, E.D.; Sateriale, A.; Gullicksrud, J.A.; Beiting, D.P.; Sullivan, M.C.; Pinkston, B.; Striepen, B. Life cycle progression and sexual development of the apicomplexan parasite Cryptosporidium parvum. Nat. Microbiol. 2019, 4, 2226–2236. [Google Scholar] [CrossRef] [Green Version]
- Morrissette, N. Targeting Toxoplasma tubules: Tubulin, microtubules, and associated proteins in a human pathogen. Eukaryot. Cell 2015, 14, 2–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment Group * | Control | Colchicine (50 μM) | Paclitaxel (50 μM) | ||
|---|---|---|---|---|---|
| Mean ± SD (μm) | Mean ± SD (μm) | p-Value † | Mean ± SD (μm) | p-Value † | |
| Long microtubule | 4.13 ± 0.41 | 4.28 ± 0.36 | 0.2079 | 4.33 ± 0.33 | 0.0683 |
| Short microtubule | 1.54 ± 0.38 | 1.47 ± 0.34 | 0.6673 | 1.55 ± 0.33 | 0.9985 |
| Taxonomic Group | Tubulin Subunit | ||
|---|---|---|---|
| Alpha | Beta | Gamma | |
| Cryptosporidia | 1 | 1 | 1 |
| Intestinal coccidia | 1 | 1–2 | 1 |
| Cystic coccidia | 3 | 2 | 1 |
| Plasmodia | 2 | 1 | 1 |
| Piroplasmids | 2 | 1 | 1 |
| Gregarines | 2 | 2 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, C.; Wang, D.; Nie, J.; Gao, X.; Yin, J.; Zhu, G. Unique Tubulin-Based Structures in the Zoonotic Apicomplexan Parasite Cryptosporidium parvum. Microorganisms 2021, 9, 1921. https://doi.org/10.3390/microorganisms9091921
Wang C, Wang D, Nie J, Gao X, Yin J, Zhu G. Unique Tubulin-Based Structures in the Zoonotic Apicomplexan Parasite Cryptosporidium parvum. Microorganisms. 2021; 9(9):1921. https://doi.org/10.3390/microorganisms9091921
Chicago/Turabian StyleWang, Chenchen, Dongqiang Wang, Jiawen Nie, Xin Gao, Jigang Yin, and Guan Zhu. 2021. "Unique Tubulin-Based Structures in the Zoonotic Apicomplexan Parasite Cryptosporidium parvum" Microorganisms 9, no. 9: 1921. https://doi.org/10.3390/microorganisms9091921