
OXA-900, a Novel OXA Sub-Family Carbapenemase Identified in Citrobacter freundii, Evades Detection by Commercial Molecular Diagnostics Tests

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Description of the Isolate
2.2. Antibiotic Susceptibly Testing
2.3. Detection of Carbapenemases
2.4. WGS Analysis
2.5. Insertion Sequences (IS)
2.6. Carbapenemase Gene Cloning and Functional Confirmation
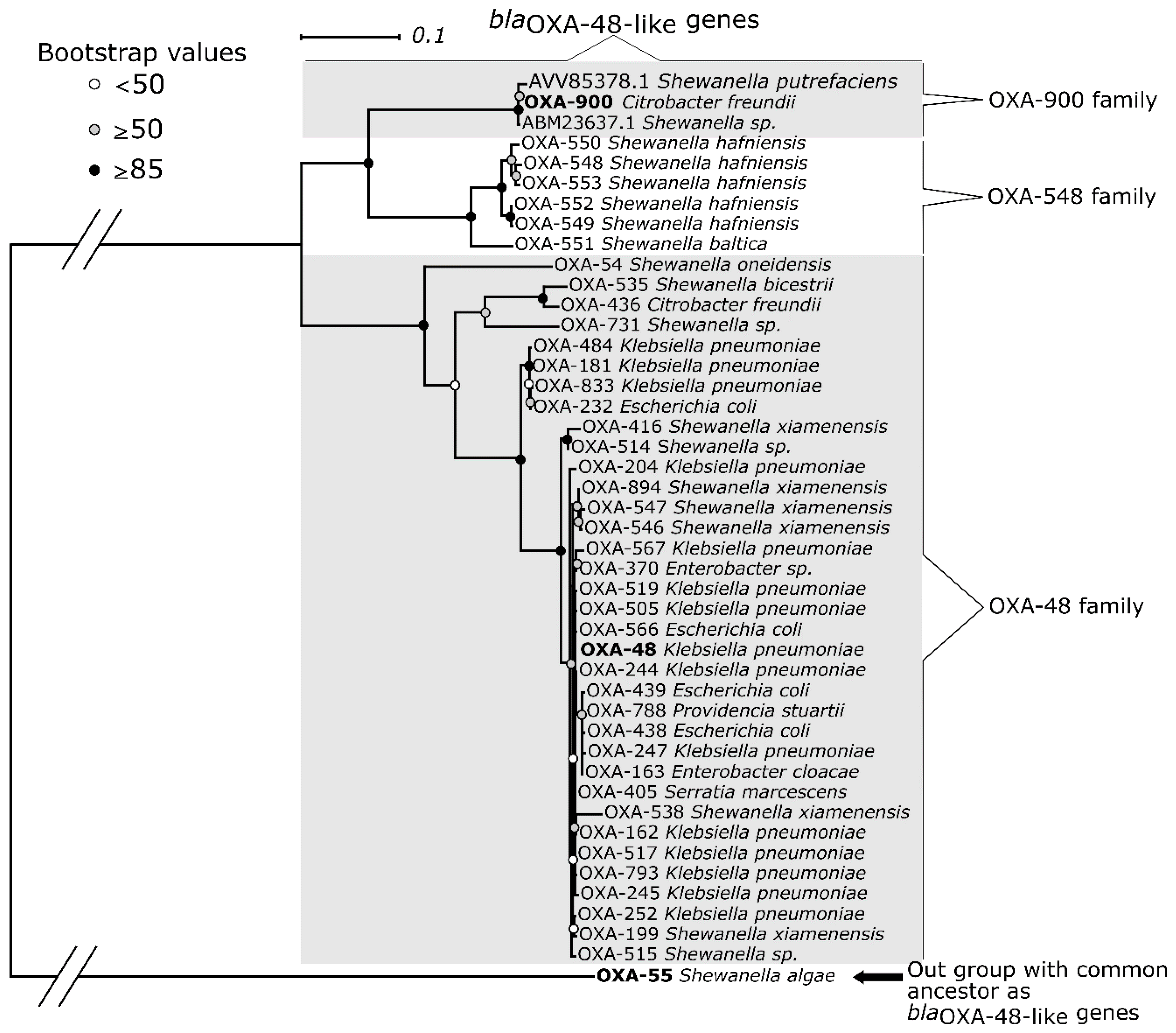
2.7. Phylogenetic Analysis of Selected blaOXA Genes
2.8. Conjugation Efficiency
2.9. Data Availability
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
| Resistance Mechanism | Antibiotic Resistance Gene | Best Hit Identity (%) | Query/Template Length | Genetic Fragment |
|---|---|---|---|---|
| Quinolone resistance protein Class C β-lactamases | qnrB38-like | 99.53 | 645/645 | Chromosome (CP046502) |
| blaCMY-65-like | 99.91 | 1146/1146 | ||
| Efflux pumps | tet(D) | 100 | 1185/1185 | Plasmid (CP046504) |
| Class A β-lactamases | blaCTX-M-3 | 100 | 876/876 | |
| Dihydropteroate synthase | sul2 | 100 | 816/816 | |
| Class A β-lactamases | blaCTX-M-39 | 100 | 876/876 | Plasmid (CP046506) |
| Class A β-lactamases | blaPER-2 | 100 | 927/927 | |
| Class D β-lactamases | blaOXA-900 | 81.5 * | 810/795 |
References
- Poirel, L.; Héritier, C.; Tolün, V.; Nordmann, P. Emergence of oxacillinase-mediated resistance to imipenem in Klebsiella pneumoniae. Antimicrob. Agents Chemother. 2004, 48, 15–22. [Google Scholar] [CrossRef] [Green Version]
- Antunes, N.T.; Lamoureaux, T.L.; Toth, M.; Stewart, N.K.; Frase, H.; Vakulenko, S.B. Class D β-Lactamases: Are they all carbapenemases? Antimicrob. Agents Chemother. 2014, 58, 2119–2125. [Google Scholar] [CrossRef] [Green Version]
- Evans, B.A.; Amyes, S.G.B. OXA β-lactamases. Clin. Microbiol. Rev. 2014, 27, 241–263. [Google Scholar] [CrossRef] [Green Version]
- Clinical and Laboratory Standards Institute (CLSI). Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically, 11th ed.; CLSI Standard M07; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018. [Google Scholar]
- Pierce, V.M.; Simner, P.J.; Lonsway, D.R.; Roe-Carpenter, D.E.; Johnson, J.K.; Brasso, W.B.; Bobenchik, A.M.; Lockett, Z.C.; Charnot-Katsikas, A.; Ferraro, M.J.; et al. Modified carbapenem inactivation method for phenotypic detection of carbapenemase production among Enterobacteriaceae. J. Clin. Microbiol. 2017, 55, 2321–2333. [Google Scholar] [CrossRef] [Green Version]
- Lerner, A.; Solter, E.; Rachi, E.; Adler, A.; Rechnitzer, H.; Miron, D.; Krupnick, L.; Sela, S.; Aga, E.; Ziv, Y.; et al. Detection and characterization of carbapenemase-producing Enterobacteriaceae in wounded Syrian patients admitted to hospitals in northern Israel. Eur. J. Clin. Microbiol. Infect. Dis. 2016, 35, 149–154. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput. Biol. 2017, 13, e1005595. [Google Scholar] [CrossRef] [Green Version]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [PubMed]
- Buchfink, B.; Xie, C.; Huson, D.H. Fast and sensitive protein alignment using DIAMOND. Nat. Methods 2015, 12, 59–60. [Google Scholar] [CrossRef] [PubMed]
- Frenk, S.; Rakovitsky, N.; Temkin, E.; Schechner, V.; Cohen, R.; Kloyzner, S.; Schwaber, M.J.; Solter, E.; Cohen, S.; Stepansky, S.; et al. Investigation of outbreaks of extended-spectrum beta-lactamase-producing Klebsiella pneumoniae in three neonatal intensive care units using whole genome sequencing. Antibiotics 2020, 9, 705. [Google Scholar] [CrossRef] [PubMed]
- Siguier, P.; Perochon, J.; Lestrade, L.; Mahillon, J.; Chandler, M. ISfinder: The reference centre for bacterial insertion sequences. Nucleic Acids Res. 2006, 34, 32–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sullivan, M.J.; Petty, N.K.; Beatson, S.A. Easyfig: A genome comparison visualizer. Bioinformatics 2011, 27, 1009–1010. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Huson, D.H.; Scornavacca, C. Dendroscope 3: An interactive tool for rooted phylogenetic trees and networks. Syst. Biol. 2012, 61, 1061–1067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ambrose, S.J.; Harmer, C.J.; Hall, R.M. Evolution and typing of IncC plasmids contributing to antibiotic resistance in Gram-negative bacteria. Plasmid 2018, 99, 40–55. [Google Scholar] [CrossRef] [PubMed]
- Hancock, S.J.; Phan, M.D.; Luo, Z.; Lo, A.W.; Peters, K.M.; Nhu, N.T.K.; Forde, B.M.; Whitfield, J.; Yang, J.; Strugnell, R.A.; et al. Comprehensive analysis of IncC plasmid conjugation identifies a crucial role for the transcriptional regulator AcaB. Nat. Microbiol. 2020, 5, 1340–1348. [Google Scholar] [CrossRef]
- Smet, A.; Van Nieuwerburgh, F.; Vandekerckhove, T.T.M.; Martel, A.; Deforce, D.; Butaye, P.; Haesebrouck, F. Complete nucleotide sequence of CTX-M-15-plasmids from clinical Escherichia coli isolates: Insertional events of transposons and insertion sequences. PLoS ONE 2010, 5, e11202. [Google Scholar] [CrossRef]
- Partridge, S.R.; Kwong, S.M.; Firth, N.; Jensen, S.O. Mobile genetic elements associated with antimicrobial resistance. Clin. Microbiol. Rev. 2018, 31, e00088-17. [Google Scholar] [CrossRef] [Green Version]
- Roberts, M.C. Antibiotic-resistant environmental bacteria and their role as reservoirs in disease. In Modeling the Transmission and Prevention of Infectious Disease; Hurst, C.J., Ed.; Springer International Publishing: Berlin/Heidelberg, Germany, 2017; pp. 187–212. [Google Scholar]
- Zheng, B.; Tan, S.; Gao, J.; Han, H.; Liu, J.; Lu, G.; Liu, D.; Yi, Y.; Zhu, B.; Gao, G.F. An unexpected similarity between antibiotic-resistant NDM-1 and beta-lactamase II from Erythrobacter litoralis. Protein Cell 2011, 2, 250–258. [Google Scholar] [CrossRef] [Green Version]
- Poirel, L.; Rodriguez-Martinez, J.-M.; Mammeri, H.; Liard, A.; Nordmann, P. Origin of plasmid-mediated quinolone resistance determinant QnrA. Antimicrob. Agents Chemother. 2005, 49, 3523–3525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samuelsen, Ø.; Hansen, F.; Aasnæs, B.; Hasman, H.; Lund, B.A.; Leiros, H.-K.S.; Lilje, B.; Janice, J.; Jakobsen, L.; Littauer, P.; et al. Dissemination and characteristics of a novel plasmid-encoded carbapenem-hydrolyzing Class D β-Lactamase, OXA-436, found in isolates from four patients at six different hospitals in Denmark. Antimicrob. Agents Chemother. 2017, 62, e01260-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tacão, M.; Araújo, S.; Vendas, M.; Alves, A.; Henriques, I. Shewanella species as the origin of blaOXA-48 genes: Insights into gene diversity, associated phenotypes and possible transfer mechanisms. Int. J. Antimicrob. Agents 2018, 51, 340–348. [Google Scholar] [CrossRef] [PubMed]


| MIC in µg/mL (CLSI Breakpoints) | ||||
|---|---|---|---|---|
| Antibiotic Class | Antibiotic | C. freundii Isolate ISCF142 | Transformant DH10β (OXA-900) | DH10β (Negative Control) |
| Carbapenems | Meropenem | 1 (S) | 1 (S) | <0.12 (S) |
| Imipenem | 1 (S) | 2 (S) | <0.5 (S) | |
| Ertapenem | 1 (I) | 1 (I) | <0.12 (S) | |
| Cephalosporins | Cefotaxime | 8 (R) | <0.5 (S) | <0.5 (S) |
| Ceftazidime | >16 (R) | <0.5 (S) | <0.5 (S) | |
| Cephalosporin combinations | Ceftazidime/ avibactam a | <0.5 (S) | <0.5 (S) | <0.5 (S) |
| Ceftolozane/ tazobactam a | >32 (R) | 8 (S) | <0.5 (S) | |
| Monobactams | Aztreonam | >32 (R) | <0.5 (S) | <0.5 (S) |
| Penicillin combinations | Piperacillin/ tazobactam a | >32 (R) | >32 (R) | 2 (S) |
| Amoxicillin/clavulanic acid b | >64 (R) | >64 (R) | <4 (S) | |
| Fluoroquinolones | Ciprofloxacin | 0.25 (S) | 0.12 (S) | 0.12 (S) |
| Polymixins | Colistin | 1 | 0.25 | 0.5 |
| Aminoglycosides | Tobramycin | 1 (S) | <1 (S) | <1 (S) |
| Tetracyclines | Tigecycline | 0.25 (S) | <0.25 (S) | <0.25 (S) |
| Sulfonamide combinations | Trimethoprim/ sulfamethoxazole c | 1 (S) | <1 (S) | <1 (S) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Frenk, S.; Rakovitsky, N.; Kon, H.; Rov, R.; Abramov, S.; Lurie-Weinberger, M.N.; Schwartz, D.; Pinco, E.; Lellouche, J.; Carmeli, Y. OXA-900, a Novel OXA Sub-Family Carbapenemase Identified in Citrobacter freundii, Evades Detection by Commercial Molecular Diagnostics Tests. Microorganisms 2021, 9, 1898. https://doi.org/10.3390/microorganisms9091898
Frenk S, Rakovitsky N, Kon H, Rov R, Abramov S, Lurie-Weinberger MN, Schwartz D, Pinco E, Lellouche J, Carmeli Y. OXA-900, a Novel OXA Sub-Family Carbapenemase Identified in Citrobacter freundii, Evades Detection by Commercial Molecular Diagnostics Tests. Microorganisms. 2021; 9(9):1898. https://doi.org/10.3390/microorganisms9091898
Chicago/Turabian StyleFrenk, Sammy, Nadya Rakovitsky, Hadas Kon, Reut Rov, Shirin Abramov, Mor Nadia Lurie-Weinberger, David Schwartz, Erica Pinco, Jonathan Lellouche, and Yehuda Carmeli. 2021. "OXA-900, a Novel OXA Sub-Family Carbapenemase Identified in Citrobacter freundii, Evades Detection by Commercial Molecular Diagnostics Tests" Microorganisms 9, no. 9: 1898. https://doi.org/10.3390/microorganisms9091898