
From Water into Sediment—Tracing Freshwater Cyanobacteria via DNA Analyses

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
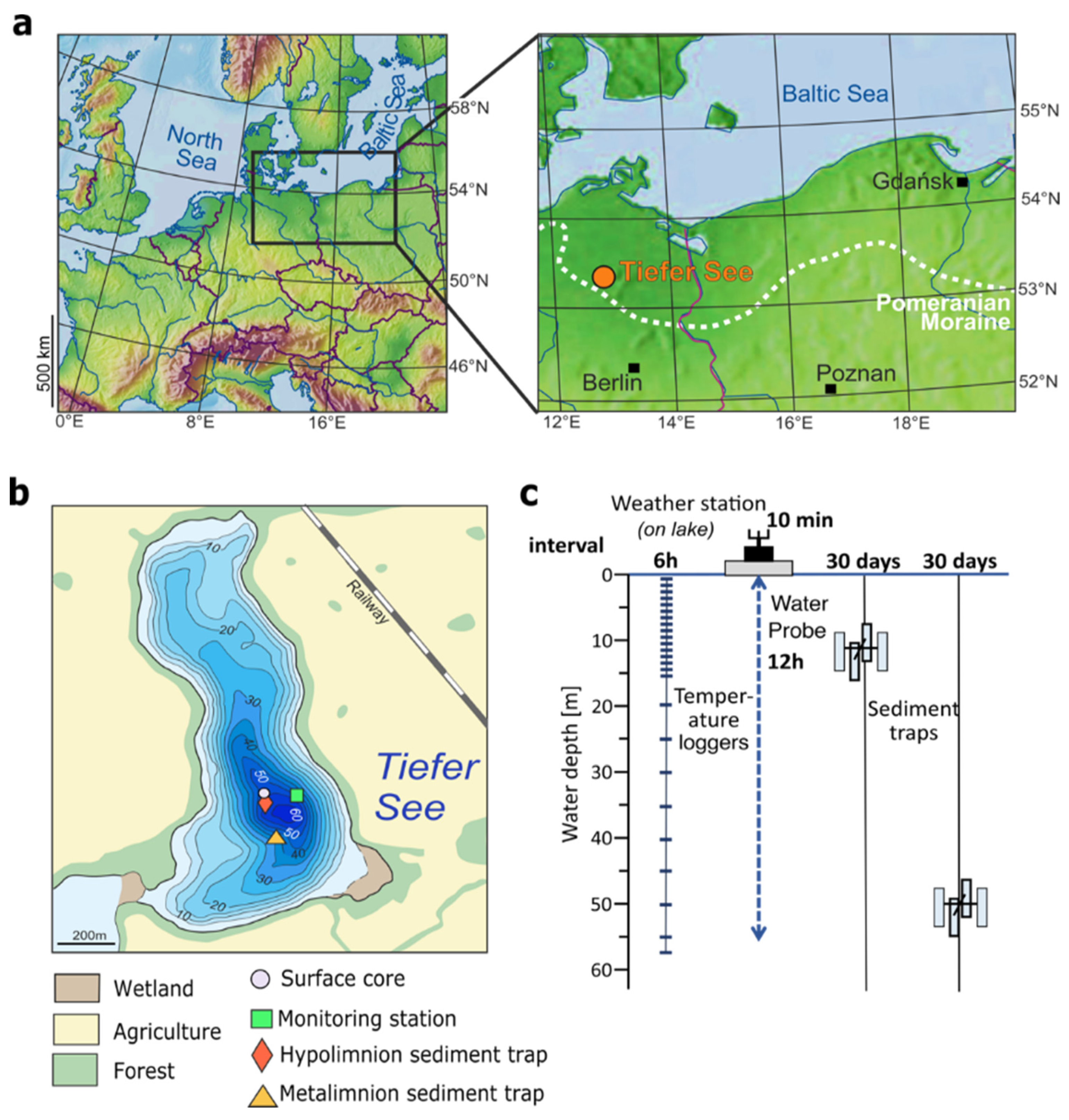
2.1. Study Site
2.2. Collection of Water, Sediment Trap Material, and Sediment Cores
2.3. Lake Physicochemical Properties
2.4. Molecular Analyses
2.5. Bioinformatics and Sequence Processing
2.6. Data Treatment and Statistics
3. Results
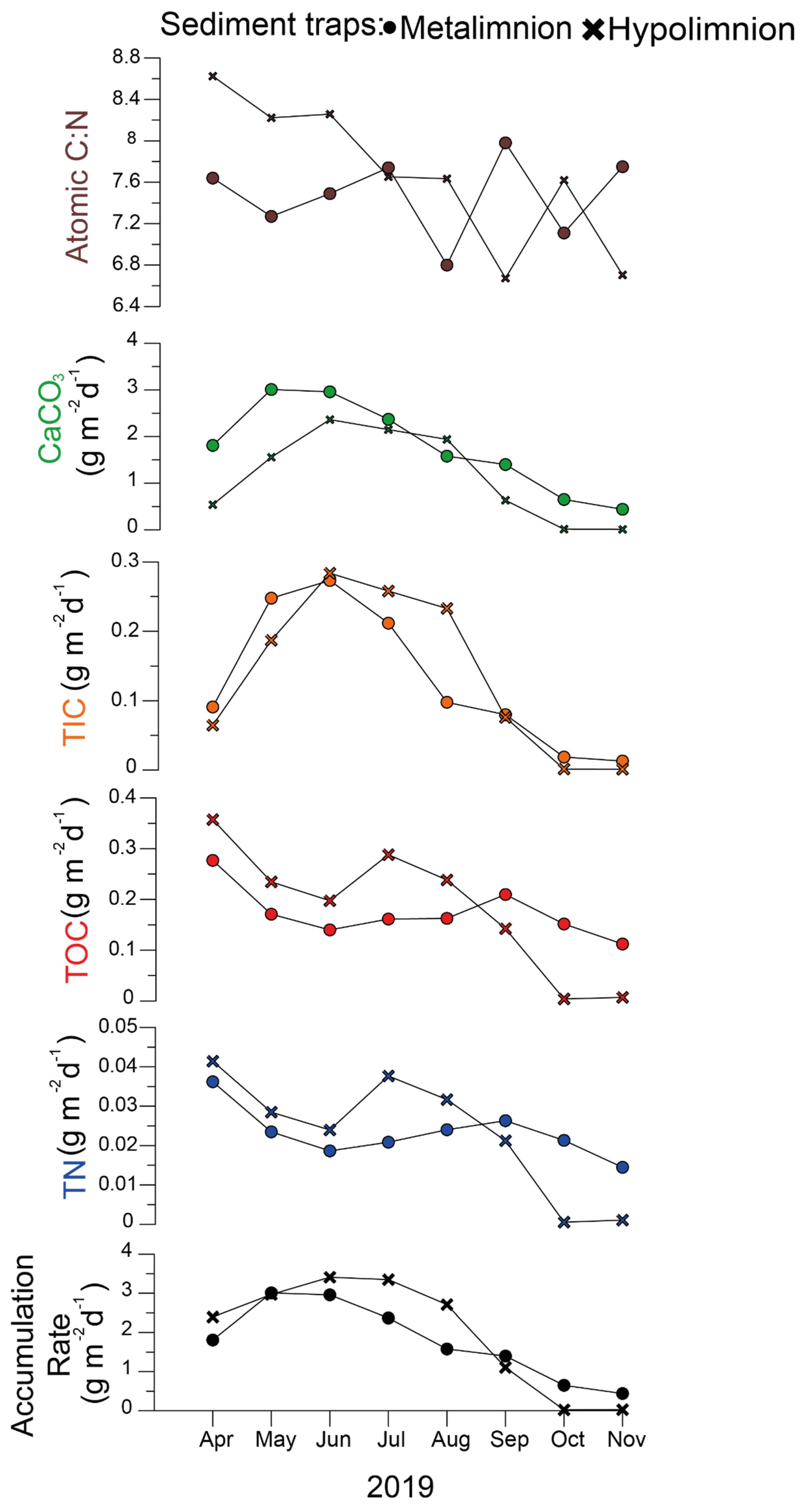
3.1. Physicochemical Properties of Pelagic Water and Trap Material
3.2. Cyanobacteria Community Structure
Cyanobacteria Abundance
3.3. Alpha and Beta Diversity
3.4. Correlation of Biotic with Physicochemical Properties
4. Discussion
4.1. Tracing Cyanobacteria from the Water Column into the Sediment
4.2. Taxa with High Deposition
4.3. Taxa with Poor Deposition
4.4. Linking Cyanobacteria Community Dynamics with Environmental Data
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vitousek, P.M.; Mooney, H.A.; Lubchenco, J.; Melillo, J.M. Human Domination of Earth’ s Ecosystems. Science 1997, 277, 494–499. [Google Scholar] [CrossRef] [Green Version]
- Fischer, W.W.; Hemp, J.; Johnson, J.E. Evolution of Oxygenic Photosynthesis. Annu. Rev. Earth Planet. Sci. 2016, 44, 647–683. [Google Scholar] [CrossRef]
- Posch, T.; Köster, O.; Salcher, M.M.; Pernthaler, J. Harmful filamentous cyanobacteria favoured by reduced water turnover with lake warming. Nat. Clim. Chang. 2012, 2, 809–813. [Google Scholar] [CrossRef] [Green Version]
- Dokulil, M.T.; Teubner, K. Deep living Planktothrix rubescens modulated by environmental constraints and climate forcing. Hydrobiologia 2012, 698, 29–46. [Google Scholar] [CrossRef] [Green Version]
- Huisman, J.; Codd, G.A.; Paerl, H.W.; Ibelings, B.W.; Verspagen, J.M.H.; Visser, P.M. Cyanobacterial blooms. Nat. Rev. Microbiol. 2018, 16, 471–483. [Google Scholar] [CrossRef]
- Cronberg, G. Blue-green algae, green algae and chrysophyceae in sediments. In Handbook of Holocene Palaeoecology and Palaeohydrology; Berglund, B., Ed.; John Wiley and Sons: Chichester, UK, 1986; pp. 507–526. [Google Scholar]
- Livingstone, D.; Jaworski, G.H.M. The viability of akinetes of blue-green algae recovered from the sediments of rostherne mere. Br. Phycol. J. 1980, 15, 357–364. [Google Scholar] [CrossRef]
- Van Geel, B.; Mur, L.R.; Ralska-Jasiewiczowa, M.; Goslar, T. Fossil akinetes of Aphanizomenon and Anabaena as indicators for medieval phosphate-eutrophication of Lake Gosciaz (Central Poland). Rev. Palaeobot. Palynol. 1994, 83, 97–105. [Google Scholar] [CrossRef] [Green Version]
- Sanger, J.E. Fossil pigments in paleoecology and paleolimnology. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1988, 62, 343–359. [Google Scholar] [CrossRef]
- Leavitt, P.R.; Findlay, D.L. Comparison of fossil pigments with 20 years of phytoplankton data from eutrophic Lake 227, Experimental Lakes Area, Ontario. Can. J. Fish. Aquat. Sci. 1994, 51, 2286–2299. [Google Scholar] [CrossRef]
- Hertzberg, S.; Liaaen-Jensen, S.; Siegelman, H.W. The carotenoids of blue-green algae. Phytochemistry 1971, 10, 3121–3127. [Google Scholar] [CrossRef]
- Swain, E.B. Measurement and interpretation of sedimentary pigments. Freshw. Biol. 1985, 15, 53–75. [Google Scholar] [CrossRef]
- Coolen, M.J.L.; Overmann, J. Analysis of subfossil molecular remains of purple sulfur bacteria in a lake sediment. Appl. Environ. Microbiol. 1998, 64, 4513–4521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domaizon, I.; Savichtcheva, O.; Debroas, D.; Arnaud, F.; Villar, C.; Pignol, C.; Alric, B.; Perga, M.E. DNA from lake sediments reveals the long-term dynamics and diversity of Synechococcus assemblages. Biogeosci. Discuss. 2013, 10, 2515–2564. [Google Scholar] [CrossRef]
- Jasser, I.; Królicka, A.; Karnkowska-Ishikawa, A. A novel phylogenetic clade of picocyanobacteria from the Mazurian lakes (Poland) reflects the early ontogeny of glacial lakes. FEMS Microbiol. Ecol. 2011, 75, 85–98. [Google Scholar] [CrossRef] [Green Version]
- Stoof-Leichsenring, K.R.; Epp, L.S.; Trauth, M.H.; Tiedemann, R. Hidden diversity in diatoms of Kenyan Lake Naivasha: A genetic approach detects temporal variation. Mol. Ecol. 2012, 21, 1918–1930. [Google Scholar] [CrossRef] [PubMed]
- Belle, S.; Parent, C. Reconstruction of Past Dynamics of Methane-Oxidizing Bacteria in Lake Sediments Using a Quantitative PCR Method: Connecting Past Environmental Changes and Microbial Community. Geomicrobiol. J. 2019, 36, 570–579. [Google Scholar] [CrossRef]
- Leavitt, P.R. A review of factors that regulate carotenoid and chlorophyll deposition and fossil pigment abundance. J. Paleolimnol. 1993, 9, 109–127. [Google Scholar] [CrossRef]
- Domaizon, I.; Winegardner, A.; Capo, E.; Gauthier, J.; Gregory-Eaves, I. DNA-based methods in paleolimnology: New opportunities for investigating long-term dynamics of lacustrine biodiversity. J. Paleolimnol. 2017, 58, 1–21. [Google Scholar] [CrossRef]
- Capo, E.; Debroas, D.; Arnaud, F.; Domaizon, I. Is Planktonic Diversity Well Recorded in Sedimentary DNA? Toward the Reconstruction of Past Protistan Diversity. Microb. Ecol. 2015, 70, 865–875. [Google Scholar] [CrossRef] [PubMed]
- Capo, E.; Giguet-Covex, C.; Rouillard, A.; Nota, K.; Heintzman, P.D.; Vuillemin, A.; Ariztegui, D.; Arnaud, F.; Belle, S.; Bertilsson, S.; et al. Lake Sedimentary DNA Research on Past Terrestrial and Aquatic Biodiversity: Overview and Recommendations. Quaternary 2021, 4, 6. [Google Scholar] [CrossRef]
- Pedersen, M.W.; Overballe-Petersen, S.; Ermini, L.; Der Sarkissian, C.; Haile, J.; Hellstrom, M.; Spens, J.; Thomsen, P.F.; Bohmann, K.; Cappellini, E.; et al. Ancient and modern environmental DNA. Philos. Trans. R. Soc. B Biol. Sci. 2015, 370, 20130383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giguet-Covex, C.; Ficetola, G.F.; Walsh, K.; Poulenard, J.; Bajard, M.; Fouinat, L.; Sabatier, P.; Gielly, L.; Messager, E.; Develle, A.L.; et al. New insights on lake sediment DNA from the catchment: Importance of taphonomic and analytical issues on the record quality. Sci. Rep. 2019, 9, 14676. [Google Scholar] [CrossRef] [PubMed]
- Ellegaard, M.; Clokie, M.R.J.; Czypionka, T.; Frisch, D.; Godhe, A.; Kremp, A.; Letarov, A.; McGenity, T.J.; Ribeiro, S.; John Anderson, N. Dead or alive: Sediment DNA archives as tools for tracking aquatic evolution and adaptation. Commun. Biol. 2020, 3, 169. [Google Scholar] [CrossRef] [Green Version]
- Gauthier, J.; Walsh, D.; Selbie, D.T.; Bourgeois, A.; Griffiths, K.; Domaizon, I.; Gregory-Eaves, I. Evaluating the congruence between DNA-based and morphological taxonomic approaches in water and sediment trap samples: Analyses of a 36-month time series from a temperate monomictic lake. Limnol. Oceanogr. 2021. [Google Scholar] [CrossRef]
- Monchamp, M.E.; Walser, J.C.; Pomati, F.; Spaak, P. Sedimentary DNA reveals cyanobacterial community diversity over 200 years in two perialpine lakes. Appl. Environ. Microbiol. 2016, 82, 6472–6482. [Google Scholar] [CrossRef] [Green Version]
- Roeser, P.; Dräger, N.; Brykała, D.; Ott, F.; Pinkerneil, S.; Gierszewski, P.; Lindemann, C.; Plessen, B.; Brademann, B.; Kaszubski, M.; et al. Advances in understanding calcite varve formation: New insights from a dual lake monitoring approach in the southern Baltic lowlands. Boreas 2021. [Google Scholar] [CrossRef]
- Dräger, N.; Plessen, B.; Kienel, U.; Słowiński, M.; Ramisch, A.; Tjallingii, R.; Pinkerneil, S.; Brauer, A. Hypolimnetic oxygen conditions influence varve preservation and δ13C of sediment organic matter in Lake Tiefer See, NE Germany. J. Paleolimnol. 2019, 62, 181–194. [Google Scholar] [CrossRef]
- Savichtcheva, O.; Debroas, D.; Kurmayer, R.; Villar, C.; Jenny, J.P.; Arnaud, F.; Perga, M.E.; Domaizon, I. Quantitative PCR enumeration of total/toxic Planktothrix rubescens and total cyanobacteria in preserved DNA isolated from lake sediments. Appl. Environ. Microbiol. 2011, 77, 8744–8753. [Google Scholar] [CrossRef] [Green Version]
- Nübel, U.; Garcia-Pichel, F.; Muyzer, G. PCR primers to amplify 16S rRNA genes from cyanobacteria. Appl. Environ. Microbiol. 1997, 63, 3327–3332. [Google Scholar] [CrossRef] [Green Version]
- Andrews, S.; Krueger, F.; Seconds-Pichon, A.; Biggins, F.; Wingett, S. FastQC: A quality control tool for high throughput sequence data. Available online: https://github.com/s-andrews/FastQC (accessed on 21 July 2021).
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [Green Version]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Hammer, Ø.; Harper, D.A.T.a.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar] [CrossRef]
- Saary, P.; Forslund, K.; Bork, P.; Hildebrand, F. RTK: Efficient rarefaction analysis of large datasets. Bioinformatics 2017, 33, 2594–2595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Legendre, P.; Gallagher, E.D. Ecologically meaningful transformations for ordination of species data. Oecologia 2001, 129, 271–280. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. vegan: Community Ecology Package. R package version 2.5-2. Cran R 2019, 2, 1–295. [Google Scholar]
- Weisbrod, B.; Wood, S.A.; Steiner, K.; Whyte-Wilding, R.; Puddick, J.; Laroche, O.; Dietrich, D.R. Is a Central Sediment Sample Sufficient? Exploring Spatial and Temporal Microbial Diversity in a Small Lake. Toxins 2020, 12, 580. [Google Scholar] [CrossRef] [PubMed]
- Leavitt, P.R.; Carpenter, S.R.; Kitchell, J.F. Whole-lake experiments: The annual record of fossil pigments and zooplankton. Limnol. Oceanogr. 1989, 34, 700–717. [Google Scholar] [CrossRef]
- Heiskanen, A.S.; Olli, K. Sedimentation and buoyancy of Aphanizomenon cf. flos-aquae (Nostocales, Cyanophyta) in a nutrient-replete and nutrient-depleted coastal area of the Baltic Sea. Phycologia 1996, 35, 94–101. [Google Scholar] [CrossRef]
- Halstvedt, C.B.; Rohrlack, T.; Andersen, T.; Skulberg, O.; Edvardsen, B. Seasonal dynamics and depth distribution of Planktothrix spp. in Lake Steinsfjorden (Norway) related to environmental factors. J. Plankton Res. 2007, 29, 471–482. [Google Scholar] [CrossRef] [Green Version]
- Gerphagnon, M.; Macarthur, D.J.; Latour, D.; Gachon, C.M.M.; Van Ogtrop, F.; Gleason, F.H.; Sime-Ngando, T. Microbial players involved in the decline of filamentous and colonial cyanobacterial blooms with a focus on fungal parasitism. Environ. Microbiol. 2015, 8, 2573–2587. [Google Scholar] [CrossRef]
- Nwosu, E.; Brauer, A.; Horn, F.; Wagner, D.; Brauer, A. Evaluating sedimentary DNA for tracing eutrophication-associated cyanobacteria species in subrecent sediments of lake Tiefer See, NE Germany. J. Paleolimnol. 2021. [Google Scholar] [CrossRef]
- Klemer, A.R.; Feuillade, J.; Feuillade, M. Cyanobacterial Blooms: Carbon and Nitrogen Limitation Have Opposite Effects on the Buoyancy of Oscillatoria. Science 1982, 215, 1629–1631. [Google Scholar] [CrossRef] [PubMed]
- Hense, I.; Beckmann, A. Towards a model of cyanobacteria life cycle-effects of growing and resting stages on bloom formation of N2-fixing species. Ecol. Modell. 2006, 195, 205–218. [Google Scholar] [CrossRef]
- Kromkamp, J.; Konopka, A.; Mur, L.R. Buoyancy regulation in a strain of Aphanizomenon flos-aquae (Cyanophycaea): The importance of carbohydrate accumulation and gas vesicle collapse. J. Gen. Microbiol. 1986, 132, 2113–2121. [Google Scholar] [CrossRef] [Green Version]
- Komárek, J.; Santanna, C.L.; Bohunická, M.; Mareš, J.; Hentschke, G.S.; Rigonato, J.; Fiore, M.F. Phenotype diversity and phylogeny of selected Scytonema-species (Cyanoprokaryota) from SE Brazil. Fottea 2013. [Google Scholar] [CrossRef] [Green Version]
- Tallberg, P.; Heiskanen, A.S. Species-specific phytoplankton sedimentation in relation to primary production along an inshore-offshore gradient in the Baltic sea. J. Plankton Res. 1998, 20, 2053–2070. [Google Scholar] [CrossRef]
- Hansson, L.A.; Rudstam, L.G.; Johnson, T.B.; Soranno, P.; Allen, Y. Patterns in algal recruitment from sediment to water in a dimictic, eutrophic lake. Can. J. Fish. Aquat. Sci. 1994, 51, 2825–2833. [Google Scholar] [CrossRef]
- Rajaniemi-Wacklin, P.; Rantala, A.; Mugnai, M.A.; Turicchia, S.; Ventura, S.; Komárková, J.; Lepistö, L.; Sivonen, K. Correspondence between phylogeny and morphology of Snowella spp. and Woronichinia naegeliana, cyanobacteria commonly occurring in lakes. J. Phycol. 2006, 42, 226–232. [Google Scholar] [CrossRef]
- Komárek, J.; Komárková-Legnerová, J. Variability of some planktic gomphosphaerioid cyanoprocaryotes in northern lakes. Nord. J. Bot. 1992, 12, 513–524. [Google Scholar] [CrossRef]
- Preston, T.; Stewart, W.D.P.; Reynolds, C.S. Bloom-forming cyanobacterium Microcystis aeruginosa overwinters on sediment surface. Nature 1980, 288, 365–367. [Google Scholar] [CrossRef]
- McDonald, K.E.; Lehman, J.T. Dynamics of aphanizomenon and microcystis (cyanobacteria) during experimental manipulation of an urban impoundment. Lake Reserv. Manag. 2013, 29, 103–115. [Google Scholar] [CrossRef]
- Kurmayer, R. Strategies for the co-existence of zooplankton with the toxic cyanobacterium Planktothrix rubescens in Lake Zurich. J. Plankton Res. 1999, 21, 659–683. [Google Scholar] [CrossRef]
- Wentzky, V.C.; Frassl, M.A.; Rinke, K.; Boehrer, B. Metalimnetic oxygen minimum and the presence of Planktothrix rubescens in a low-nutrient drinking water reservoir. Water Res. 2019, 148, 208–218. [Google Scholar] [CrossRef]
- Kyle, M.; Haande, S.; Sønstebø, J.; Rohrlack, T. Amplification of DNA in sediment cores to detect historic Planktothrix occurrence in three Norwegian lakes. J. Paleolimnol. 2014, 53, 61–72. [Google Scholar] [CrossRef]
- Rohrlack, T.; Edvardsen, B.; Skulberg, R.; Halstvedt, C.B.; Utkilen, H.C.; Ptacnik, R.; Skulberg, O.M. Oligopeptide chemotypes of the toxic freshwater cyanobacterium Planktothrix can form subpopulations with dissimilar ecological traits. Limnol. Oceanogr. 2008, 53, 1279–1293. [Google Scholar] [CrossRef]
- Callieri, C.; Stockner, J. Picocyanobacteria success in oligotrophic lakes: Fact or fiction? J. Limnol. 2000, 59, 72–76. [Google Scholar] [CrossRef]
- Jezberová, J.; Komárková, J. Morphological transformation in a freshwater Cyanobium sp. induced by grazers. Environ. Microbiol. 2007, 9, 1858–1862. [Google Scholar] [CrossRef] [PubMed]
- Callieri, C. Picophytoplankton in Freshwater Ecosystems: The Importance of Small-Sized Phototrophs. Freshw. Rev. 2008, 1, 1–28. [Google Scholar] [CrossRef]
- Callieri, C.; Cronberg, G.; Stockner, J.G. Freshwater picocyanobacteria: Single cells, microcolonies and colonial forms. In Ecology of cyanobacteria II: Their Diversity in Space and Time; Whitton, B., Ed.; Springer: Dordrecht, The Netherlands, 2012; pp. 229–269. [Google Scholar]
- Coolen, M.J.L.; Talbot, H.M.; Abbas, B.A.; Ward, C.; Schouten, S.; Volkman, J.K.; Sinninghe Damsté, J.S. Sources for sedimentary bacteriohopanepolyols as revealed by 16S rDNA stratigraphy. Environ. Microbiol. 2008, 10, 1783–1803. [Google Scholar] [CrossRef]
- Joint, I.; Owens, N.; Pomeroy, A. Seasonal production of photosynthetic picoplankton and nanoplankton in the Celtic Sea. Mar. Ecol. Prog. Ser. 1986, 28, 251–258. [Google Scholar] [CrossRef]
- Ballot, A.; Fastner, J.; Wiedner, C. Paralytic shellfish poisoning toxin-producing cyanobacterium Aphanizomenon gracile in Northeast Germany. Appl. Environ. Microbiol. 2010, 76, 1173–1180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.; Ficetola, G.F. Numerical methods for sedimentary-ancient-DNA-based study on past biodiversity and ecosystem functioning. Environ. DNA 2020, 2, 115–129. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PERMANOVA | Pairwise | Epi | Meta | Hypo | TrapM | TrapH | Sed | |
|---|---|---|---|---|---|---|---|---|
| Summary | Epi | 0.012 | 0.0775 | 0.0015 | 0.0015 | 0.786 | ||
| Permutation N: | 9999 | Meta | 3.962 | 0.027 | 0.0015 | 0.0015 | 0.8835 | |
| Total sum of squares: | 8.29 | Hypo | 3.244 | 4.66 | 0.0015 | 0.0015 | 1 | |
| Within-group sum of squares: | 5.438 | TrapM | 8.943 | 10.24 | 5.544 | 1 | 1 | |
| F: | 6.085 | TrapH | 11.15 | 12.78 | 6.638 | 0.4568 | 1 | |
| p (same): | 0.0001 | Sed | 4.397 | 5.151 | 4.244 | 0.7717 | 1.116 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nwosu, E.C.; Roeser, P.; Yang, S.; Ganzert, L.; Dellwig, O.; Pinkerneil, S.; Brauer, A.; Dittmann, E.; Wagner, D.; Liebner, S. From Water into Sediment—Tracing Freshwater Cyanobacteria via DNA Analyses. Microorganisms 2021, 9, 1778. https://doi.org/10.3390/microorganisms9081778
Nwosu EC, Roeser P, Yang S, Ganzert L, Dellwig O, Pinkerneil S, Brauer A, Dittmann E, Wagner D, Liebner S. From Water into Sediment—Tracing Freshwater Cyanobacteria via DNA Analyses. Microorganisms. 2021; 9(8):1778. https://doi.org/10.3390/microorganisms9081778
Chicago/Turabian StyleNwosu, Ebuka Canisius, Patricia Roeser, Sizhong Yang, Lars Ganzert, Olaf Dellwig, Sylvia Pinkerneil, Achim Brauer, Elke Dittmann, Dirk Wagner, and Susanne Liebner. 2021. "From Water into Sediment—Tracing Freshwater Cyanobacteria via DNA Analyses" Microorganisms 9, no. 8: 1778. https://doi.org/10.3390/microorganisms9081778