
Degradation of Bile Acids by Soil and Water Bacteria


Abstract
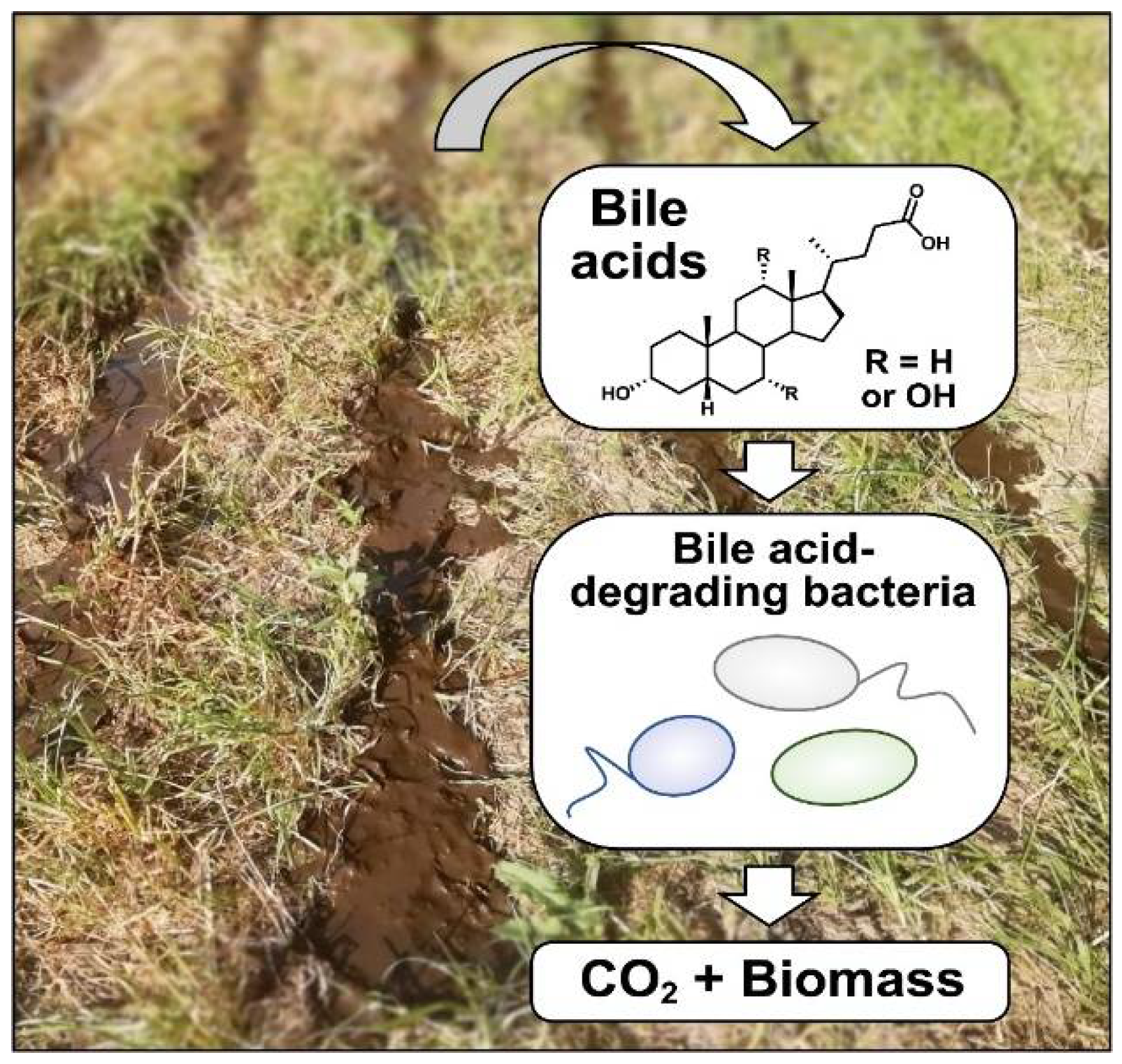
:1. Bile Acids and Their Occurrence in the Environment
2. Bacterial Bile Acid Degradation
2.1. Diversity of Bile Acid-Degrading Bacteria
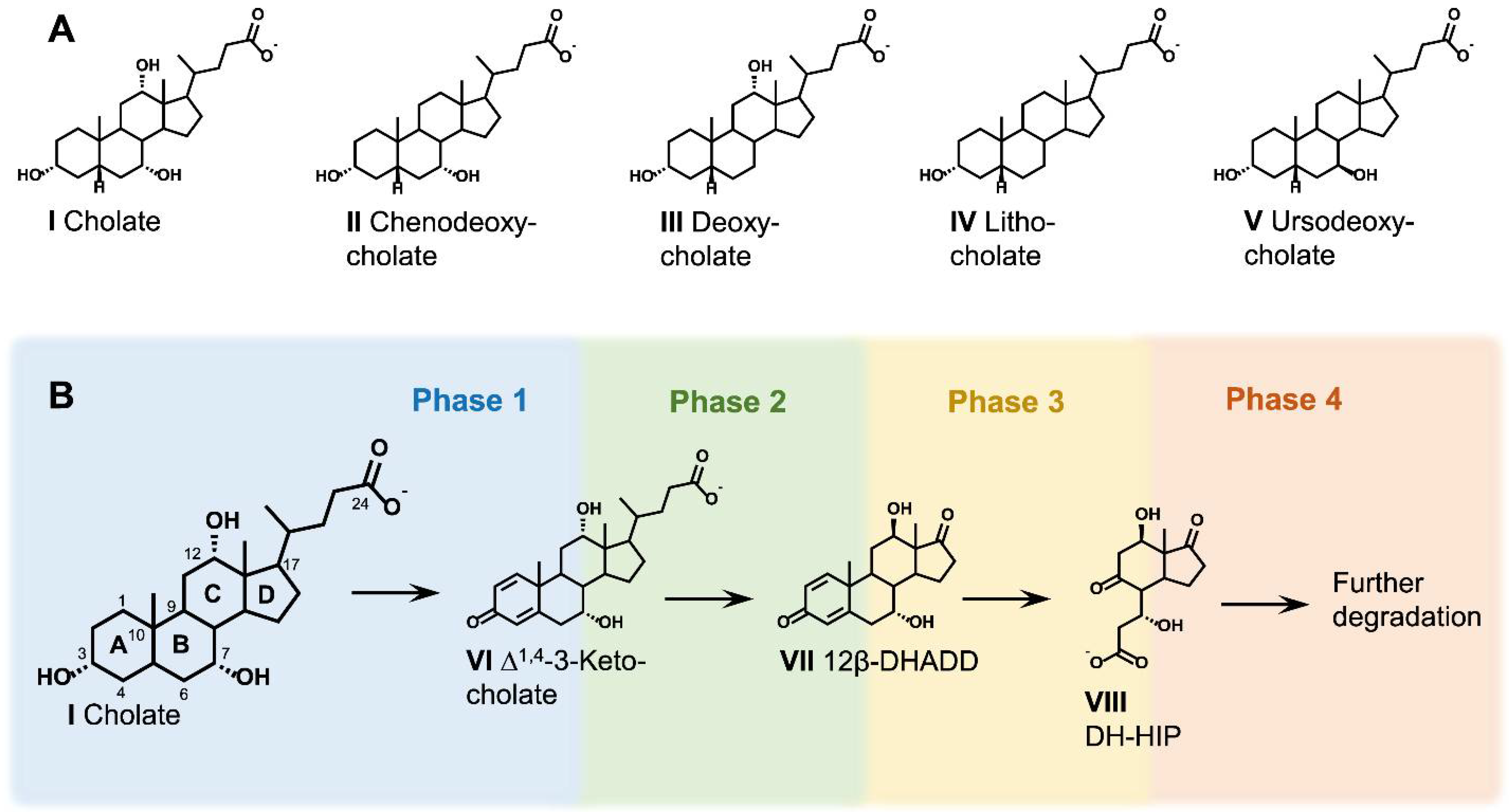
2.2. General Aspects of Metabolic Bile Acid Degradation Pathways

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bile Acid Degradation Pathway | Function | P. stutzeri Chol1 | C. testosteroni CNB-2 *** | R. jostii RHA1 | Sphingobium sp. Strain Chol11 |
|---|---|---|---|---|---|
| Cleavage of conjugated bile acids | Bile salt amidase | C211_RS11020 | CTCNB1_RS06560 (ORF26) CTCNB1_TS06555 (ORF25) | unknown (RHA1_RS22310) | Nov2c227 (Bsa) |
| C5 side-chain degradation | CoA-ligase | C211_RS11125 (StdA1 *) | CTCNB1_RS06840 | RHA1_RS28415 (CasG) | Nov2c230 (SclA) |
| ACADs | C211_RS11115 (Scd1A) | CTCNB1_RS06830 | RHA1_RS28395 (CasC) | Nov2c221 (Scd4A) | |
| C211_RS11120 (Scd1B) | CTCNB1_RS06835 | --- | Nov2c222 (Scd4B) | ||
| Enoyl-CoA hydratase | C211_RS11210 (Shy1) | CTCNB1_RS06880 | RHA1_RS28400 (CasD) | absent | |
| Steroid aldolase | C211_RS11205 (Sal1) | CTCNB1_RS06875 | absent | absent | |
| Steroid aldehyde dehydrogenase | C211_RS11010 (Sad) | CTCNB1_RS06680 | absent | absent | |
| 2-hydroxy-CoA dehydrogenase | absent | absent | RHA1_RS28410 | absent | |
| Steroid thiolase | absent | absent | RHA1_RS28390 (CasB) | absent | |
| C3 side-chain degradation | CoA-ligase | C211_RS11185 (StdA2 *) | CTCNB1_RS06820 | RHA1_RS28425 (CasI) | absent |
| ACADs | C211_RS11105 (Scd2A) | CTCNB1_RS06815 | RHA1_RS28440 (CasL) | absent | |
| C211_RS11090 (Scd2B) | CTCNB1_RS06800 | RHA1_RS28450 (CasN) | absent | ||
| Enoyl-CoA hydratase | C211_RS11085 (Shy2) | CTCNB1_RS06795 | RHA1_RS28445 (CasM) | absent | |
| C211_RS11095 | CTCNB1_RS06805 | RHA1_RS28455 (CasO) | absent | ||
| Steroid aldolase | C211_RS11100 (Sal2) | CTCNB1_RS06810 | RHA1_RS28460 (CasP) | absent | |
| A-ring oxidation | 3α-Hydroxysteroid dehydrogenase | C211_RS10975 | CTCNB1_RS06750 (3α-HSD) | unknown | Nov2c6 |
| ∆1-Ketosteroid dehydrogenase | C211_RS11030 (Δ1-KstD) | CTCNB1_RS06925 (TesH) | RHA1_RS28305 (KstD3) | Nov2c82 | |
| --- | --- | RHA1_RS28380 (KstD3b) | --- | ||
| Δ4-5α-Ketosteroid dehydrogenase | C211_RS11110 | CTCNB1_RS06930 (TesI) | RHA1_RS27810 | Nov2c17 | |
| Δ4-5β-Ketosteroid dehydrogenase | Unknown | CTCNB1_RS06740 | RHA1_RS28420 (CasH) | Nov2c19 (Δ4-5β-KSTD1) | |
| 9,10-seco pathway | 3-Ketosteroid-9α hydroxylase, oxygenase | C211_RS11300 | CTCNB1_RS06665 | RHA1_RS28370 (KshA3) | Nov2c407 |
| --- | --- | --- | Nov2c430 | ||
| --- | --- | --- | Nov2c440 | ||
| 3-Ketosteroid-9α hydroxylase, reductase | C211_RS11040 | CTCNB1_RS06935 (ORF17) | RHA1_RS28480 (KshB3) | absent | |
| 9,10-seco-steroid hydroxylase, oxygenase | C211_RS11025 | CTCNB1_RS06920 (TesA2) | RHA1_RS28325 (HsaA3) | Nov2c349 | |
| 9,10-seco-steroid hydroxylase, reductase | C211_RS11005 | CTCNB1_RS06915 (TesA1) | RHA1_RS28295 (HsaB3) | absent (Nov2c347) | |
| 9,10-seco-steroid dioxygenase | C211_RS11215 | CTCNB1_RS06510 (TesB) | RHA1_RS28330 (HsaC3) | Nov2c350 | |
| 4,5,9,10-diseco-steroid hydroxylase | C211_RS11155 | CTCNB1_RS06910 (TesD) | RHA1_RS28300 (HsaD3) | Nov2c348 | |
| C/D-ring (HIP) side-chain degradation | CoA-ligase | C211_RS11045 (StdA3 *) | CTCNB1_RS06940 (ScdA) | RHA1_RS22410 ** (FadD3) | Nov2c359 |
| ACAD | C211_RS11065 (Scd3A) | CTCNB1_RS06570 (ScdC1) | RHA1_RS22390 ** | Nov2c367 | |
| C211_RS11070 (Scd3B) | CTCNB1_RS06575 (ScdC2) | RHA1_RS22415 ** | Nov2c361 | ||
| Enoyl-CoA hydratase | C211_RS11075 | CTCNB1_RS06585 (ScdD) | unknown | Nov2c364 | |
| C211_RS11270 | --- | --- | --- | ||
| 2-hydroxy-acyl-CoA dehydrogenase | C211_RS11260 | CTCNB1_RS06565 (ScdE) | RHA1_RS22710 ** | Absent (Nov2c360, Nov2c362) | |
| HIP thiolase | C211_RS11275 | CTCNB1_RS06590 (ORF33) | RHA1_RS28520 | Nov2c358 | |
| C/D-ring (HIC) degradation | 5-OH HIC-CoA reductase | C211_RS11235 | CTCNB1_RS06530 (ScdK) | RHA1_RS22685 ** (IpdC) | Nov2c354 |
| 5-Oxo HIC-CoA oxidase | C211_RS11265 | CTCNB1_RS06580 (ScdG) | RHA1_RS22420 ** (IpdF) | Nov2c363 | |
| HIEC-CoA hydrolase | unknown | CTCNB1_RS06535 (ScdY) | RHA1_RS27700 ** (EchA20) | Nov2c355 | |
| COCHEA-CoA hydrolase, α-subunit | C211_RS11220 | CTCNB1_RS06515 (ScdL1) | RHA1_RS22695 ** (IpdA) | Nov2c351 | |
| COCHEA-CoA hydrolase, β-subunit | C211_RS11225 | CTCNB1_RS06520 (ScdL2) | RHA1_RS22690 ** (IpdB) | Nov2c352 | |
| β-Keto CoA thiolase | C211_RS11255 | CTCNB1_RS06550 (ScdF) | RHA1_RS22430 ** (FadA6) | Nov2c366 | |
| ACAD | C211_RS11245 | CTCNB1_RS06540 (ORF21) | RHA1_RS22400 ** (FadE31) | Nov2c356 | |
| ACAD | C211_RS11250 | CTCNB1_RS06545 (ORF22) | RHA1_RS22395 ** (FadE32) | Nov2c357 | |
| Hydratase | C211_RS11230 | CTCNB1_RS06525 (ScdN) | RHA1_RS22405 | Nov2c353 | |
| 2-hydroxy-hexa-2,4-dienoate degradation | 2-Hydroxypenta-2,4-dienoate hydratase | C211_RS10995 | CTCNB1_RS06905 (TesE) | RHA1_RS28310 (HsaE3) | Nov2c346 |
| Acetaldehyde dehydrogenase | C211_RS10985 | CTCNB1_RS06895 (TesG) | RHA1_RS28315 (HsaG3) | Nov2c344 | |
| 4-Hydroxy-2-ketovalerate aldolase | C211_RS10990 | CTCNB1_RS06900 (TesF) | RHA1_RS28320 (HsaF3) | Nov2c345 | |
| Transformation of 12OH | 12α-Hydroxy-steroid dehydrogenase | C211_RS11165 | CTCNB1_RS06490 (SteA) | unknown | Nov2c15 |
| 12-Keto-steroid hydrogenase | C211_RS11170 | CTCNB1_RS06495 (SteB) | unknown | Nov2c16 | |
| 12β-hydroxy steroid dehydratase | C211_RS11175 (Hsh1) | CTCNB1_RS06500 (ORF7) | unknown | Nov2c12 | |
| Steroid oxidoreducatse | C211_RS11180 (Sor1) | CTCNB1_RS06505 (ORF6) | unknown | Nov2c13 | |
| Δ4,6 variant | 7α-Hydroxysteroid dehydratase | absent | absent | absent | Nov2c400 (Hsh2) |
2.3. Uptake of Bile Acids
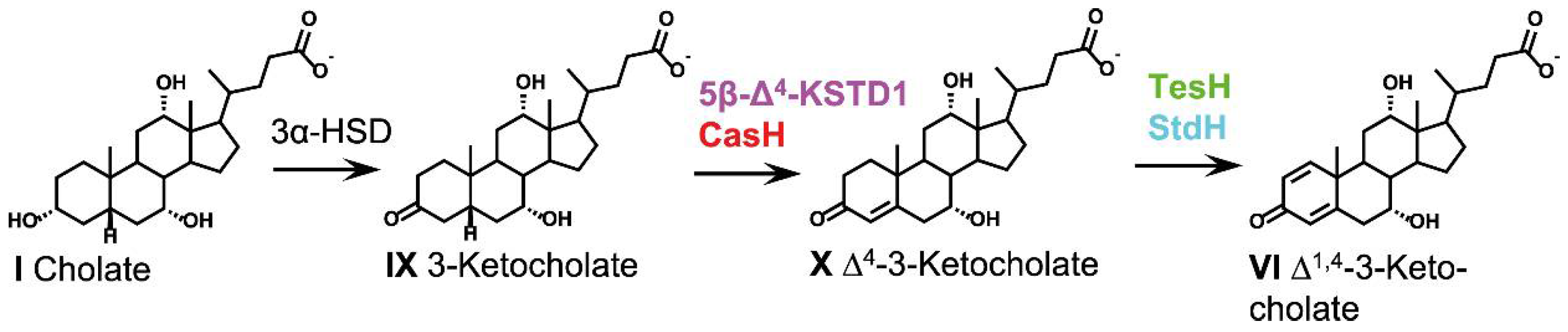
2.4. Oxidative Reactions at the A-Ring (Phase 1)

2.4.1. 3α-Hydroxysteroid Dehydrogenases
2.4.2. 3-Ketosteroid Dehydrogenases
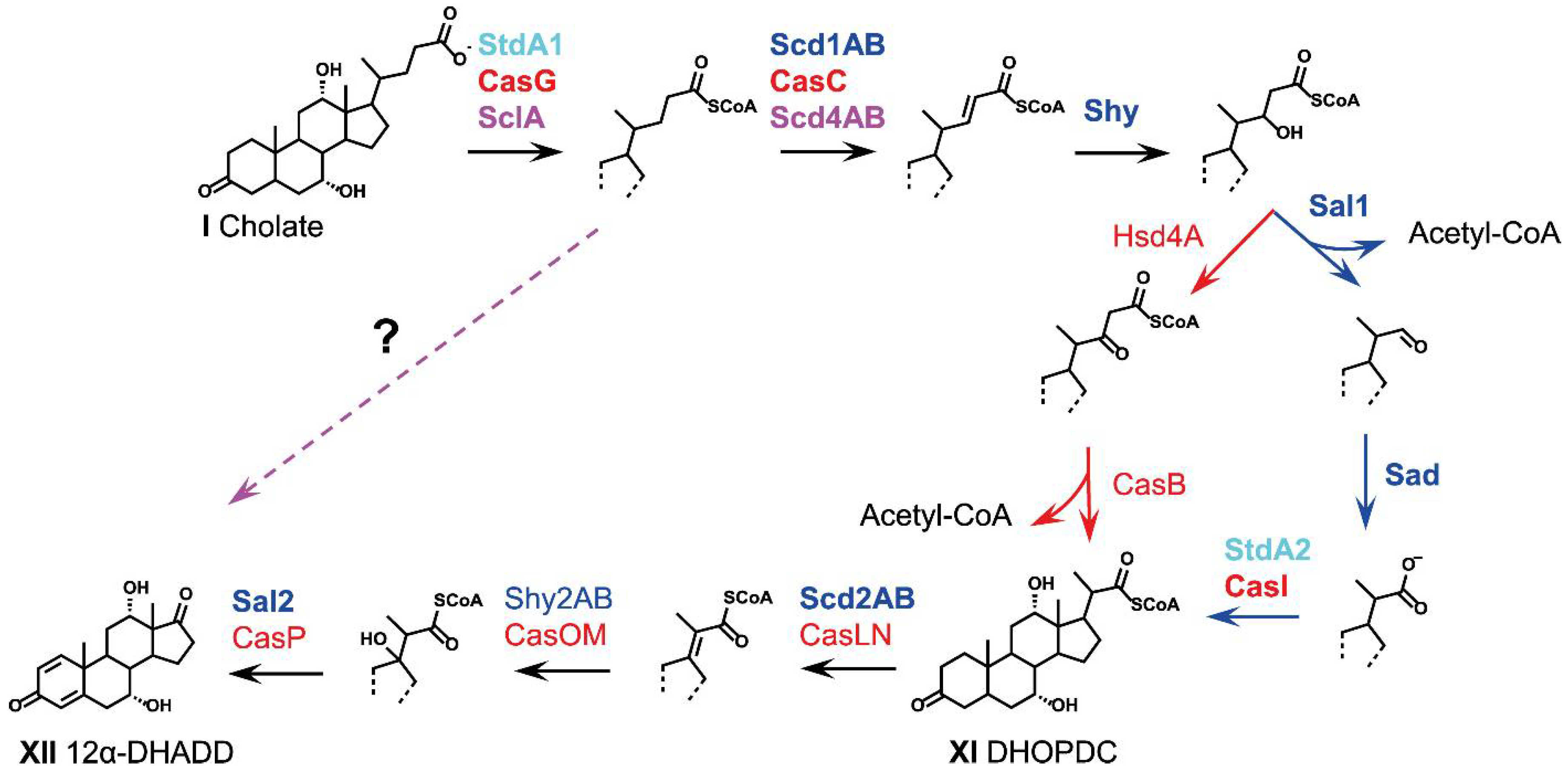
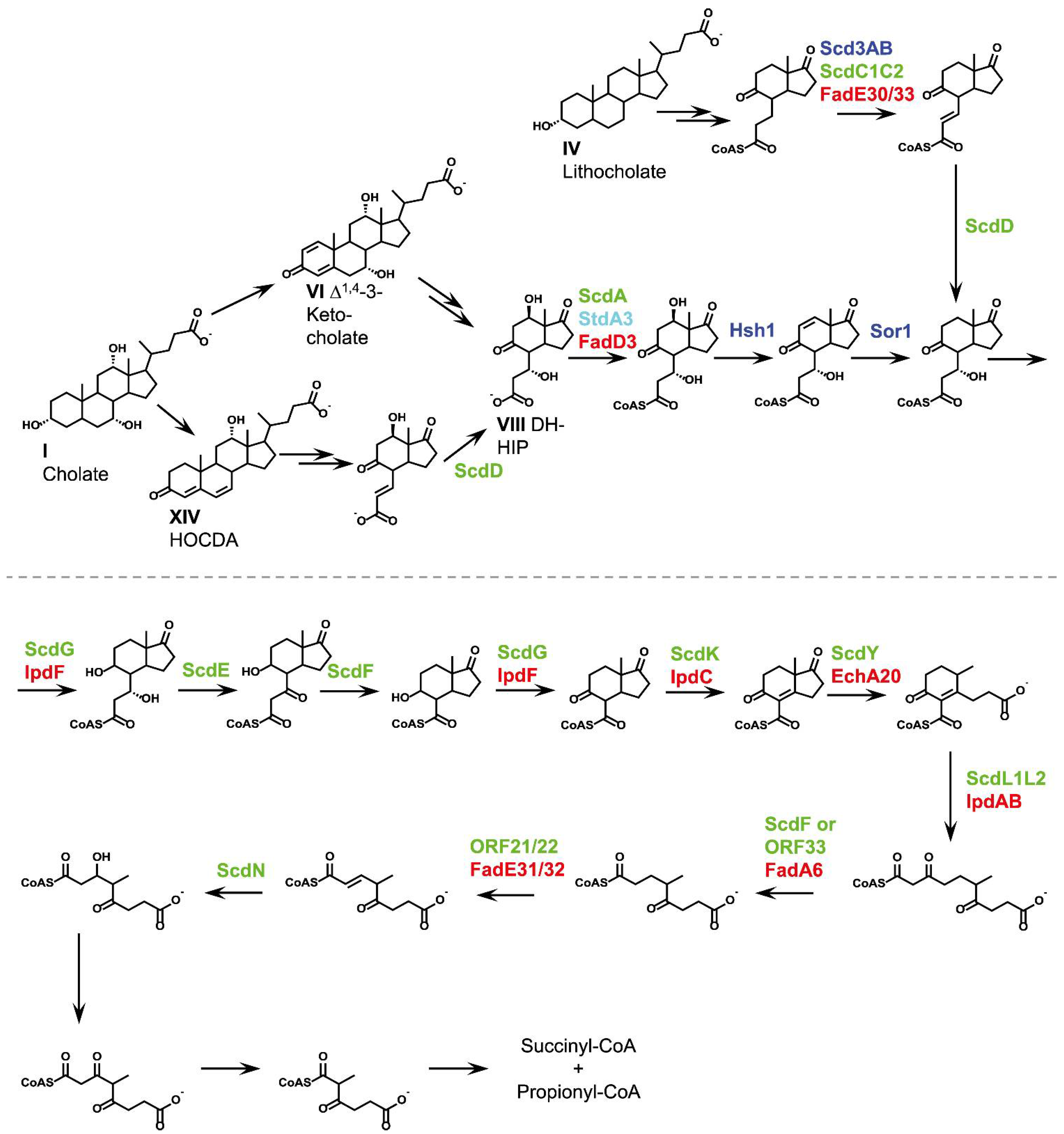
2.5. Side-Chain Degradation (Phase 2)

2.5.1. Cleavage of Conjugated Bile Acids
2.5.2. Coenzyme-A Activation
2.5.3. α,β-Dehydrogenation
2.5.4. Hydroxylation of the Double Bond
2.5.5. Release of Acetyl-CoA and Formation of C3-Carboxylic Side Chains
2.5.6. Release of Propionyl-CoA and Formation of Androstadienediones (ADDs)
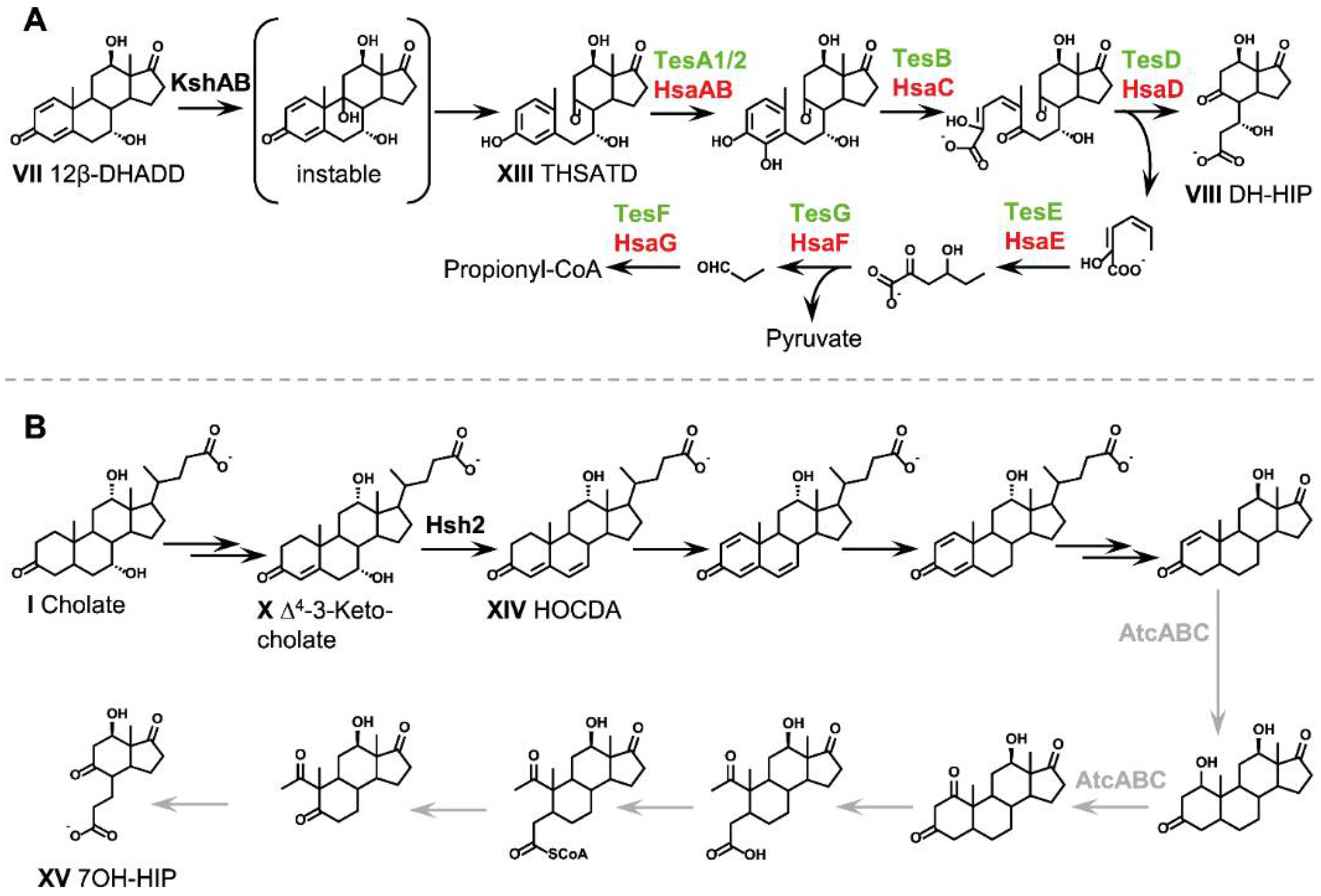
2.6. Aerobic Cleavage of the Steroid Skeleton via the 9,10-Seco Pathway (Phase 3a)
2.6.1. 9α-Hydroxylation: Opening of the B-Ring

2.6.2. Opening and Hydrolysis of the A-Ring
2.7. Anaerobic Cleavage of the Steroid Skeleton via the 2,3-Seco Pathway (Phase 3b)
2.8. C/D-Ring Degradation (Phase 4)

2.9. Fate of Hydroxy Groups on the Steroid Nucleus
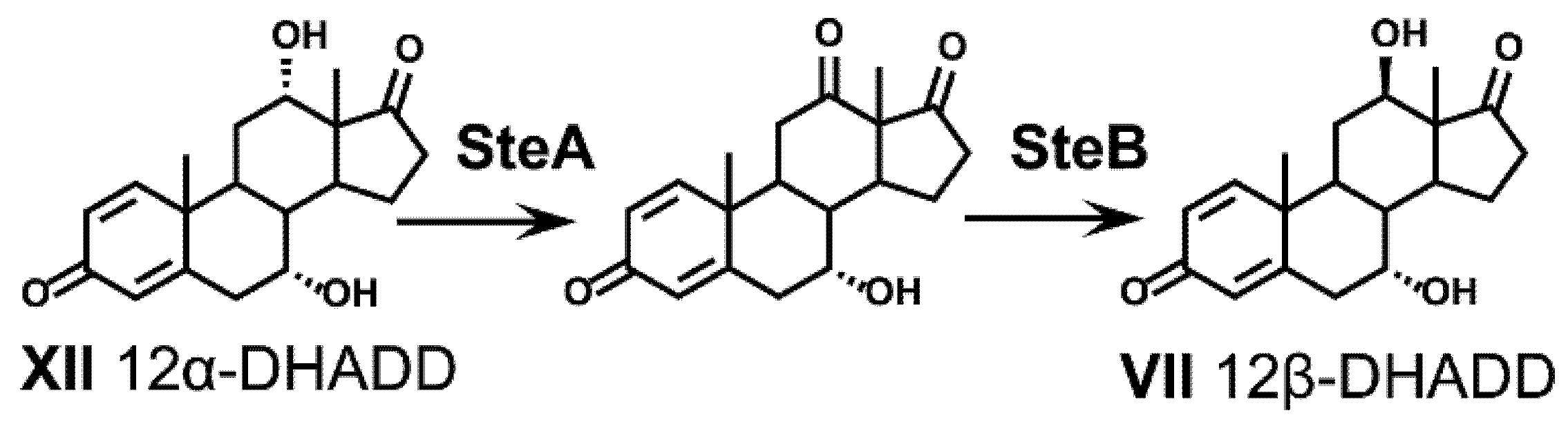
2.9.1. Fate of the 12α-Hydroxy Group
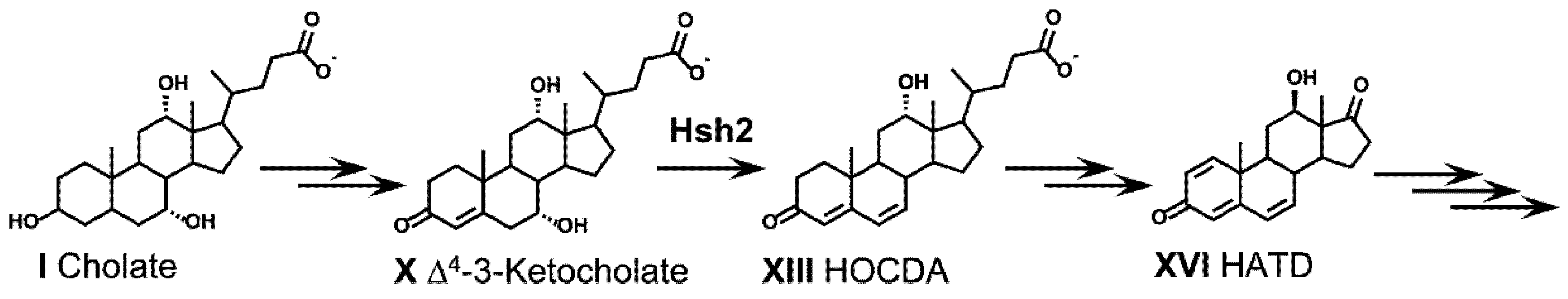
2.9.2. Fate of the 7α-Hydroxy Group
2.9.3. Dehydratation of 7-Hydroxy Groups during Bile Acid Degradation via Δ4,6-3-Keto Intermediates
3. Resistance Mechanisms against the Toxic Effects of Bile Acids
4. Regulation of Bile-Acid Degradation
5. Potential Ecological Effects of Bacterial Bile-Acid Degradation
6. Applied Aspects of Bacterial Bile-Acid Degradation
7. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Hofmann, A.F.; Hagey, L.R.; Krasowski, M.D. Bile salts of vertebrates: Structural variation and possible evolutionary significance. J. Lipid Res. 2010, 51, 226–246. [Google Scholar] [CrossRef] [Green Version]
- Vítek, L.; Haluzík, M. The role of bile acids in metabolic regulation. J. Endocrinol. 2016, 228, R85–R96. [Google Scholar] [CrossRef]
- Hylemon, P.B.; Zhou, H.; Pandak, W.M.; Ren, S.; Gil, G.; Dent, P. Bile acids as regulatory molecules. J. Lipid Res. 2009, 50, 1509–1520. [Google Scholar] [CrossRef] [Green Version]
- Ridlon, J.M.; Kang, D.-J.J.; Hylemon, P.B. Bile salt biotransformations by human intestinal bacteria. J. Lipid Res. 2006, 47, 241–259. [Google Scholar] [CrossRef] [Green Version]
- Funabashi, M.; Grove, T.L.; Wang, M.; Varma, Y.; McFadden, M.E.; Brown, L.C.; Guo, C.; Higginbottom, S.; Almo, S.C.; Fischbach, M.A. A metabolic pathway for bile acid dehydroxylation by the gut microbiome. Nature 2020, 582, 566–570. [Google Scholar] [CrossRef]
- Ridlon, J.M. Conceptualizing the vertebrate sterolbiome. Appl. Environ. Microbiol. 2020, 86, 1–22. [Google Scholar] [CrossRef]
- Doden, H.L.; Ridlon, J.M. Microbial hydroxysteroid dehydrogenases: From alpha to omega. Microorganisms 2021, 9, 469. [Google Scholar] [CrossRef]
- Winston, J.A.; Theriot, C.M. Diversification of host bile acids by members of the gut microbiota. Gut Microbes 2020, 11, 158–171. [Google Scholar] [CrossRef]
- Ridlon, J.; Kang, D.; Hylemon, P.; Bajaj, J. Bile acids and the gut microbiome. Curr. Opin. Gastroenterol. 2014, 30, 332–338. [Google Scholar] [CrossRef] [Green Version]
- Kriaa, A.; Bourgin, M.; Potiron, A.; Mkaouar, H.; Jablaoui, A.; Gérard, P.; Maguin, E.; Rhimi, M. Microbial impact on cholesterol and bile acid metabolism: Current status and future prospects. J. Lipid Res. 2019, 60, 323–332. [Google Scholar] [CrossRef] [Green Version]
- Tyagi, P.; Edwards, D.R.; Coyne, M.S. Use of sterol and bile acid biomarkers to identify domesticated animal sources of fecal pollution. Water. Air. Soil Pollut. 2008, 187, 263–274. [Google Scholar] [CrossRef]
- Mendelski, M.N.; Dölling, R.; Feller, F.M.; Hoffmann, D.; Ramos Fangmeier, L.; Ludwig, K.C.; Yücel, O.; Mährlein, A.; Paul, R.J.; Philipp, B. Steroids originating from bacterial bile acid degradation affect Caenorhabditis elegans and indicate potential risks for the fauna of manured soils. Sci. Rep. 2019, 9, 11120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tyagi, P.; Edwards, D.R.; Coyne, M.S. Fecal sterol and bile acid biomarkers: Runoff concentrations in animal waste-amended pastures. Water. Air. Soil Pollut. 2009, 198, 45. [Google Scholar] [CrossRef]
- Elhmmali, M.M.; Roberts, D.J.; Evershed, R.P. Combined analysis of bile acids and sterols/stanols from riverine particulates to assess sewage discharges and other fecal sources. Environ. Sci. Technol. 2000, 34, 39–46. [Google Scholar] [CrossRef]
- Buchinger, T.J.; Li, W.; Johnson, N.S. Bile salts as semiochemicals in fish. Chem. Senses 2014, 39, 647–654. [Google Scholar] [CrossRef]
- Meredith, T.L.; Caprio, J.; Kajiura, S.M. Sensitivity and specificity of the olfactory epithelia of two elasmobranch species to bile salts. J. Exp. Biol. 2012, 215, 2660–2667. [Google Scholar] [CrossRef] [Green Version]
- Merino, E.; Barrientos, A.; Rodríguez, J.; Naharro, G.; Luengo, J.M.; Olivera, E.R. Isolation of cholesterol- and deoxycholate-degrading bacteria from soil samples: Evidence of a common pathway. Appl. Microbiol. Biotechnol. 2013, 97, 891–904. [Google Scholar] [CrossRef]
- Holert, J.; Yücel, O.; Suvekbala, V.; Kulić, Ž.; Möller, H.; Philipp, B. Evidence of distinct pathways for bacterial degradation of the steroid compound cholate suggests the potential for metabolic interactions by interspecies cross-feeding. Environ. Microbiol. 2014, 16, 1424–1440. [Google Scholar] [CrossRef] [Green Version]
- Philipp, B.; Erdbrink, H.; Suter, M.J.F.; Schink, B. Degradation of and sensitivity to cholate in Pseudomonas sp. strain Chol1. Arch. Microbiol. 2006, 185, 192–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holert, J.; Cardenas, E.; Bergstrand, L.H.; Zaikova, E.; Hahn, A.S.; Hallam, S.J.; Mohn, W.W. Metagenomes reveal global distribution of bacterial steroid catabolism in natural, engineered, and host environments. mBio 2018, 9, e02345-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lengeler, J.; Drews, G.; Schlegel, H. Biology of the Prokaryotes; Lengeler, J.W., Drews, G., Schlegel, H.G., Eds.; Thieme: Stuttgart, Germany, 2009; ISBN 9781444313307. [Google Scholar]
- Bull, I.D.; Lockheart, M.J.; Elhmmali, M.M.; Roberts, D.J.; Evershed, R.P. The origin of faeces by means of biomarker detection. Environ. Int. 2002, 27, 647–654. [Google Scholar] [CrossRef]
- Feller, F.M.; Wöhlbrand, L.; Holert, J.; Schnaars, V.; Mohn, W.W.; Rabus, R.; Philipp, B. Proteome, bioinformatic and functional analyses reveal a distinct and conserved metabolic pathway for bile salt degradation in the Sphingomonadaceae. Appl. Environ. Microbiol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Horinouchi, M.; Hayashi, T.; Kudo, T. Steroid degradation in Comamonas testosteroni. J. Steroid Biochem. Mol. Biol. 2012, 129, 4–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yücel, O.; Borgert, S.R.; Poehlein, A.; Niermann, K.; Philipp, B. The 7α-hydroxysteroid dehydratase Hsh2 is essential for anaerobic degradation of the steroid skeleton of 7α-hydroxyl bile salts in the novel denitrifying bacterium Azoarcus sp. strain Aa7. Environ. Microbiol. 2019, 21, 800–813. [Google Scholar] [CrossRef]
- Birkenmaier, A.; Holert, J.; Erdbrink, H.; Möller, H.M.; Friemel, A.; Schönenberger, R.; Suter, M.J.F.; Klebensberger, J.; Philipp, B. Biochemical and genetic investigation of initial reactions in aerobic degradation of the bile acid cholate in Pseudomonas sp. strain Chol1. J. Bacteriol. 2007, 189, 7165–7173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergstrand, L.H.; Cardenas, E.; Holert, J.; van Hamme, J.D.; Mohn, W.W. Delineation of steroid-degrading microorganisms through comparative genomic analysis. mBio 2016, 7, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Mohn, W.W.; Wilbrink, M.H.; Casabon, I.; Stewart, G.R.; Liu, J.; van der Geize, R.; Eltis, L.D. Gene cluster encoding cholate catabolism in Rhodococcus spp. J. Bacteriol. 2012, 194, 6712–6719. [Google Scholar] [CrossRef] [Green Version]
- Guevara, G.; de las Heras, L.F.; Perera, J.; Llorens, J.M.N. Functional characterization of 3-ketosteroid 9α-hydroxylases in Rhodococcus ruber strain chol-4. J. Steroid Biochem. Mol. Biol. 2017, 172, 176–187. [Google Scholar] [CrossRef]
- Shtratnikova, V.Y.; Schelkunov, M.I.; Fokina, V.V.; Bragin, E.Y.; Lobastova, T.G.; Shutov, A.A.; Kazantsev, A.V.; Donova, M. V Genome-wide transcriptome profiling provides insight on cholesterol and lithocholate degradation mechanisms in Nocardioides simplex VKM Ac-2033D. Genes 2020, 11, 1229. [Google Scholar] [CrossRef]
- Philipp, B. Bacterial degradation of bile salts. Appl. Microbiol. Biotechnol. 2011, 89, 903–915. [Google Scholar] [CrossRef] [Green Version]
- Olivera, E.R.; Luengo, J.M. Steroids as environmental compounds recalcitrant to degradation: Genetic mechanisms of bacterial biodegradation pathways. Genes 2019, 10, 512. [Google Scholar] [CrossRef] [Green Version]
- Holert, J.; Kulić, Ž.; Yücel, O.; Suvekbala, V.; Suter, M.J.F.; Möller, H.M.; Philipp, B. Degradation of the acyl side chain of the steroid compound cholate in Pseudomonas sp. strain Chol1 proceeds via an aldehyde intermediate. J. Bacteriol. 2013, 195, 585–595. [Google Scholar] [CrossRef] [Green Version]
- Holert, J.; Jagmann, N.; Philipp, B. The essential function of genes for a hydratase and an aldehyde dehydrogenase for growth of Pseudomonas sp. strain Chol1 with the steroid compound cholate indicates an aldolytic reaction step for deacetylation of the side chain. J. Bacteriol. 2013, 195, 3371–3380. [Google Scholar] [CrossRef] [Green Version]
- Barrientos, Á.; Merino, E.; Casabon, I.; Rodríguez, J.; Crowe, A.M.; Holert, J.; Philipp, B.; Eltis, L.D.; Olivera, E.R.; Luengo, J.M. Functional analyses of three acyl-CoA synthetases involved in bile acid degradation in Pseudomonas putida DOC21. Environ. Microbiol. 2015, 17, 47–63. [Google Scholar] [CrossRef] [PubMed]
- Horinouchi, M.; Hayashi, T.; Koshino, H.; Malon, M.; Hirota, H.; Kudo, T. Identification of 9α-hydroxy-17-oxo-1,2,3,4,10,19-hexanorandrost-6- en-5-oic acid and β-oxidation products of the C-17 side chain in cholic acid degradation by Comamonas testosteroni TA441. J. Steroid Biochem. Mol. Biol. 2014, 143, 306–322. [Google Scholar] [CrossRef] [PubMed]
- Horinouchi, M.; Hayashi, T.; Koshino, H.; Malon, M.; Yamamoto, T.; Kudo, T. Identification of genes involved in inversion of stereochemistry of a C-12 hydroxyl group in the catabolism of cholic acid by Comamonas testosteroni TA441. J. Bacteriol. 2008, 190, 5545–5554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horinouchi, M.; Hayashi, T.; Koshino, H.; Kudo, T. ORF18-disrupted mutant of Comamonas testosteroni TA441 accumulates significant amounts of 9,17-dioxo-1,2,3,4,10,19-hexanorandrostan-5-oic acid and its derivatives after incubation with steroids. J. Steroid Biochem. Mol. Biol. 2006, 101, 78–84. [Google Scholar] [CrossRef] [PubMed]
- Horinouchi, M.; Koshino, H.; Malon, M.; Hirota, H.; Hayashi, T. Steroid degradation in Comamonas testosteroni TA441: Identification of the entire β-oxidation cycle of the cleaved B ring. Appl. Environ. Microbiol. 2019, 85, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Yücel, O.; Drees, S.; Jagmann, N.; Patschkowski, T.; Philipp, B. An unexplored pathway for degradation of cholate requires a 7α-hydroxysteroid dehydratase and contributes to a broad metabolic repertoire for the utilization of bile salts in Novosphingobium sp. strain Chol11. Environ. Microbiol. 2016, 18, 5187–5203. [Google Scholar] [CrossRef] [PubMed]
- Yücel, O.; Holert, J.; Ludwig, K.C.; Thierbach, S.; Philipp, B. A novel steroidcoenzyme A ligase from Novosphingobium sp. strain Chol11 is essential for an alternative degradation pathway for bile salts. Appl. Environ. Microbiol. 2018, 84, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Feller, F.M.; Marke, G.; Drees, S.L.; Wöhlbrand, L.; Rabus, R.; Philipp, B. Substrate inhibition of 5β-Δ4-3-ketosteroid dehydrogenase in Sphingobium sp. strain Chol11 acts as circuit breaker during growth with toxic bile salts. Front. Microbiol. 2021, 1, 655312. [Google Scholar] [CrossRef]
- Swain, K.; Casabon, I.; Eltis, L.D.; Mohn, W.W. Two transporters essential for reassimilation of novel cholate metabolites by Rhodococcus jostii RHA1. J. Bacteriol. 2012, 194, 6720–6727. [Google Scholar] [CrossRef] [Green Version]
- Wipperman, M.F.; Sampson, N.S.; Thomas, S.T.; Thomas, S.T. Pathogen roid rage: Cholesterol utilization by Mycobacterium tuberculosis. Crit. Rev. Biochem. Mol. Biol. 2014, 49, 269–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galán, B.; García-Fernández, J.; Felpeto-Santero, C.; Fernández-Cabezón, L.; García, J.L. Bacterial metabolism of steroids. In Aerobic Utilization of Hydrocarbons, Oils and Lipids; Rojo, F., Ed.; Springer International Publishing: Cham, Switzerland, 2017; pp. 1–22. ISBN 978-3-319-39782-5. [Google Scholar]
- Petrusma, M.; Van Der Geize, R.; Dijkhuizen, L. 3-Ketosteroid 9α-hydroxylase enzymes: Rieske non-heme monooxygenases essential for bacterial steroid degradation. Antonie Leeuwenhoek 2014, 106, 157–172. [Google Scholar] [CrossRef] [Green Version]
- Dresen, C.; Lin, L.Y.C.; D’Angelo, I.; Tocheva, E.I.; Strynadka, N.; Eltis, L.D. A flavin-dependent monooxygenase from Mycobacterium tuberculosis involved in cholesterol catabolism. J. Biol. Chem. 2010, 285, 22264–22275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayakawa, S. Microbial transformation of bile acids. A unified scheme for bile acid degradation, and hydroxylation of bile acids. Z. Allg. Mikrobiol. 1982, 22, 309–326. [Google Scholar] [CrossRef] [PubMed]
- Warnke, M.; Jacoby, C.; Jung, T.; Agne, M.; Mergelsberg, M.; Starke, R.; Jehmlich, N.; von Bergen, M.; Richnow, H.-H.; Brüls, T.; et al. A patchwork pathway for oxygenase-independent degradation of side chain containing steroids. Environ. Microbiol. 2017, 19, 4684–4699. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.H.; Lee, T.H.; Ismail, W.; Tsai, C.Y.; Lin, C.W.; Tsai, Y.W.; Chiang, Y.R. An oxygenase-independent cholesterol catabolic pathway operates under oxic conditions. PLoS ONE 2013, 8, e66675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holert, J.; Alam, I.; Larsen, M.; Antunes, A.; Bajic, V.B.; Stingl, U.; Philipp, B. Genome sequence of Pseudomonas sp. strain Chol1, a model organism for the degradation of bile salts and other steroid compounds. Genome Announc. 2013, 1, e00014-12. [Google Scholar] [CrossRef] [Green Version]
- Horinouchi, M.; Kurita, T.; Yamamoto, T.; Hatori, E.; Hayashi, T.; Kudo, T.; Hirosawa, W.-S. Steroid degradation gene cluster of Comamonas testosteroni consisting of 18 putative genes from meta-cleavage enzyme gene tesB to regulator gene tesR. Biochem. Biophys. Res. Commun. 2004, 324, 597–604. [Google Scholar] [CrossRef]
- Cabral, D.J.; Small, D.M.; Lilly, H.S.; Hamilton, J.A. Transbilayer movement of bile acids in model membranes. Biochemistry 1987, 26, 1801–1804. [Google Scholar] [CrossRef]
- Thanassi, D.G.; Cheng, L.W.; Nikaido, H. Active efflux of bile salts by Escherichia coli. J. Bacteriol. 1997, 179, 2512–2518. [Google Scholar] [CrossRef] [Green Version]
- Mallonee, D.H.; Hylemon, P.B. Sequencing and expression of a gene encoding a bile acid transporter from Eubacterium sp. strain VPI 12708. J. Bacteriol. 1996, 178, 7053–7058. [Google Scholar] [CrossRef] [Green Version]
- Ibero, J.; Galán, B.; Rivero-Buceta, V.; García, J.L. Unraveling the 17β-estradiol degradation pathway in Novosphingobium tardaugens NBRC 16725. Front. Microbiol. 2020, 11, 588300. [Google Scholar] [CrossRef] [PubMed]
- Somalinga, V.; Mohn, W.W. Rhodococcus jostii porin A (RjpA) functions in cholate uptake. Appl. Environ. Microbiol. 2013, 79, 6191–6193. [Google Scholar] [CrossRef] [PubMed]
- Mohn, W.W.; Van Der Geize, R.; Stewart, G.R.; Okamoto, S.; Liu, J.; Dijkhuizen, L.; Eltis, L.D. The actinobacterial mce4 locus encodes a steroid transporter. J. Biol. Chem. 2008, 283, 35368–35374. [Google Scholar] [CrossRef] [Green Version]
- Pandey, A.K.; Sassetti, C.M. Mycobacterial persistence requires the utilization of host cholesterol. Proc. Natl. Acad. Sci. USA 2008, 105, 4376–4380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oppermann, U.C.T.; Maser, E. Characterization of a 3α-hydroxysteroid dehydrogenase/carbonyl reductase from the Gram-negative bacterium Comamonas testosteroni. Eur. J. Biochem. 1996, 241, 744–749. [Google Scholar] [CrossRef] [PubMed]
- Maser, E.; Xiong, G.; Grimm, C.; Ficner, R.; Reuter, K. 3α-hydroxysteroid dehydrogenase/carbonyl reductase from Comamonas testosteroni: Biological significance, three-dimensional structure and gene regulation. Chem. Biol. Interact. 2001, 130–132, 707–722. [Google Scholar] [CrossRef]
- Kisiela, M.; Skarka, A.; Ebert, B.; Maser, E. Hydroxysteroid dehydrogenases (HSDs) in bacteria—A bioinformatic perspective. J. Steroid Biochem. Mol. Biol. 2012, 129, 31–46. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Gao, X.; Hong, L.; Ma, L.; Li, Y. Expression, purification and functional characterization of a novel 3α-hydroxysteroid dehydrogenase from Pseudomonas aeruginosa. Protein Expr. Purif. 2015, 115, 102–108. [Google Scholar] [CrossRef]
- Davidson, S.J.; Talalay, P. Purification and mechanism of action of a steroid Δ4-5-β-dehydrogenase. J. Biol. Chem. 1966, 241, 906–915. [Google Scholar] [CrossRef]
- Bar-Even, A.; Noor, E.; Savir, Y.; Liebermeister, W.; Davidi, D.; Tawfik, D.S.; Milo, R. The moderately efficient enzyme: Evolutionary and physicochemical trends shaping enzyme parameters. Biochemistry 2011, 50, 4402–4410. [Google Scholar] [CrossRef] [PubMed]
- Horinouchi, M.; Hayashi, T.; Yamamoto, T.; Kudo, T. A new bacterial steroid degradation gene cluster in Comamonas testosteroni TA441 which consists of aromatic-compound degradation genes for seco-steroids and 3-ketosteroid dehydrogenase genes. Appl. Environ. Microbiol. 2003, 69, 4421–4430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Oosterwijk, N.; Knol, J.; Dijkhuizen, L.; Van Der Geize, R.; Dijkstra, B.W. Structure and catalytic mechanism of 3-ketosteroid-Δ4-(5α)-dehydrogenase from Rhodococcus jostii RHA1 genome. J. Biol. Chem. 2012, 287, 30975–30983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dijkstra, B.W.; Van Oosterwijk, N.; Rohman, A. Structure and catalytic mechanism of 3-ketosteroid dehydrogenases. Procedia Chem. 2016, 18, 3–11. [Google Scholar] [CrossRef] [Green Version]
- Rohman, A.; Dijkstra, B.W. Application of microbial 3-ketosteroid Δ1-dehydrogenases in biotechnology. Biotechnol. Adv. 2021, 49, 107751. [Google Scholar] [CrossRef]
- Olivera, E.R.; de la Torre, M.; Barrientos, Á.; Luengo, J.M. Steroid catabolism in bacteria: Genetic and functional analyses of stdH and stdJ in Pseudomonas putida DOC21. Can. J. Biotechnol. 2018, 2, 88–99. [Google Scholar] [CrossRef] [Green Version]
- Florin, C.; Kohler, T.; Grandguillot, M.; Plesiat, P. Comamonas testosteroni 3-ketosteroid-Δ4-5α-dehydrogenase: Gene and protein characterization. J. Bacteriol 1996, 178, 3322–3330. [Google Scholar] [CrossRef] [Green Version]
- Knol, J.; Bodewits, K.; Hessels, G.I.; Dijkhuizen, L.; van der Geize, R. 3-Keto-5α-steroid Δ1-dehydrogenase from Rhodococcus erythropolis SQ1 and its orthologue in Mycobacterium tuberculosis H37Rv are highly specific enzymes that function in cholesterol catabolism. Biochem. J. 2008, 410, 339–346. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Ren, Y.; He, J.; Cheng, S.; Yuan, J.; Ge, F.; Li, W.; Zhang, Y.; Xie, G. Multiplicity of 3-ketosteroid Δ1-dehydrogenase enzymes in Gordonia neofelifaecis NRRL B-59395 with preferences for different steroids. Ann. Microbiol. 2015, 65, 1961–1971. [Google Scholar] [CrossRef]
- Rohman, A.; Dijkstra, B.W. The role and mechanism of microbial 3-ketosteroid Δ1-dehydrogenases in steroid breakdown. J. Steroid Biochem. Mol. Biol. 2019, 191, 105366. [Google Scholar] [CrossRef]
- Rohman, A.; Van Oosterwijk, N.; Thunnissen, A.M.W.H.; Dijkstra, B.W. Crystal structure and site-directed mutagenesis of 3-ketosteroid Δ1-dehydrogenase from Rhodococcus erythropolis SQ1 explain its catalytic mechanism. J. Biol. Chem. 2013, 288, 35559–35568. [Google Scholar] [CrossRef] [Green Version]
- Itagaki, E.; Matushita, H.; Hatta, T. Steroid transhydrogenase activity of 3-ketosteroid-Δ1-dehydrogenase from Nocardia corallina. J. Biochem. 1990, 108, 122–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jerussif, R.; Ringold, H.J. The mechanism of the bacterial C-1,2 dehydrogenation of steroids. III. Kinetics and isotope effects. Biochemistry 1965, 4, 2113–2126. [Google Scholar] [CrossRef]
- Feller, F.M.; Richtsmeier, P.; Wege, M.; Philipp, B. Comparative analysis of bile-salt degradation in Sphingobium sp. strain Chol11 and P. stutzeri Chol1 reveals functional diversity of β-proteobacterial steroid degradation enzymes and suggests a novel reaction sequence for side-chain degradation involving a hydroxylation step. bioRxiv 2021, 2021. [Google Scholar] [CrossRef]
- Jones, B.V.; Begley, M.; Hill, C.; Gahan, C.G.M.; Marchesi, J.R. Functional and comparative metagenomic analysis of bile salt hydrolase activity in the human gut microbiome. Proc. Natl. Acad. Sci. USA 2008, 105, 13580–13585. [Google Scholar] [CrossRef] [Green Version]
- Rösch, V.; Denger, K.; Schleheck, D.; Smits, T.H.M.; Cook, A.M. Different bacterial strategies to degrade taurocholate. Arch. Microbiol 2008, 190, 11–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, Z.; Lee, B.H. Bile salt hydrolases: Structure and function, substrate preference, and inhibitor development. Protein Sci. 2018, 27, 1742–1754. [Google Scholar] [CrossRef]
- Casabon, I.; Crowe, A.M.; Liu, J.; Eltis, L.D. FadD3 is an acyl-CoA synthetase that initiates catabolism of cholesterol rings C and D in Actinobacteria. Mol. Microbiol. 2013, 87, 269–283. [Google Scholar] [CrossRef]
- Warnke, M.; Jung, T.; Jacoby, C.; Agne, M.; Feller, F.M.; Philipp, B.; Seiche, W.; Breit, B.; Boll, M. Functional characterization of three specific acyl-coenzyme A synthetases involved in anaerobic cholesterol degradation in Sterolibacterium denitrificans Chol1S. Appl. Environ. Microbiol. 2018, 84, e02721-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holert, J.; Yücel, O.; Jagmann, N.; Prestel, A.; Möller, H.M.; Philipp, B. Identification of bypass reactions leading to the formation of one central steroid degradation intermediate in metabolism of different bile salts in Pseudomonas sp. strain Chol1. Environ. Microbiol. 2016, 18, 3373–3389. [Google Scholar] [CrossRef] [PubMed]
- Wipperman, M.F.; Yang, M.; Thomas, S.T.; Sampson, N.S. Shrinking the fadE proteome of Mycobacterium tuberculosis: Insights into cholesterol metabolism through identification of an α2β2 heterotetrameric acyl coenzyme A dehydrogenase family. J. Bacteriol. 2013, 195, 4331–4341. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.; Lu, R.; Guja, K.E.; Wipperman, M.F.; St. Clair, J.R.; Bonds, A.C.; Garcia-Diaz, M.; Sampson, N.S. Unraveling cholesterol catabolism in Mycobacterium tuberculosis: ChsE4-ChsE5 α2β2 acyl-CoA dehydrogenase initiates β-oxidation of 3-oxo-cholest-4-en-26-oyl CoA. ACS Infect. Dis. 2016, 1, 110–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stirling, A.J.; Gilbert, S.E.; Conner, M.; Mallette, E.; Kimber, M.S.; Seah, S.Y.K. A key glycine in bacterial steroid-degrading acyl-CoA dehydrogenases allows flavin-ring repositioning and modulates substrate side chain specificity. Biochemistry 2020, 197, 1360–1367. [Google Scholar] [CrossRef] [PubMed]
- Ruprecht, A.; Maddox, J.; Stirling, A.J.; Visaggio, N.; Seah, S.Y.K. Characterization of novel acyl coenzyme A dehydrogenases involved in bacterial steroid degradation. J. Bacteriol. 2015, 197, 1360–1367. [Google Scholar] [CrossRef] [Green Version]
- Pidugu, L.S.; Maity, K.; Ramaswamy, K. Analysis of proteins with the “Hot dog” fold: Prediction of function and identification of catalytic residues of hypothetical proteins. BMC Struct. Biol. 2009, 9, 37. [Google Scholar] [CrossRef] [Green Version]
- Haußmann, U.; Wolters, D.A.; Fränzel, B.; Eltis, L.D.; Poetsch, A. Physiological adaptation of the Rhodococcus jostii RHA1 membrane proteome to steroids as growth substrates. J. Proteome Res. 2013, 12, 1188–1198. [Google Scholar] [CrossRef]
- Haapalainen, A.M.; Meriläinen, G.; Wierenga, R.K. The thiolase superfamily: Condensing enzymes with diverse reaction specificities. Trends Biochem. Sci. 2006, 31, 64–71. [Google Scholar] [CrossRef]
- Schaefer, C.M.; Lu, R.; Nesbitt, N.M.; Schiebel, J.; Sampson, N.S.; Kisker, C. FadA5 a thiolase from Mycobacterium tuberculosis: A steroid-binding pocket reveals the potential for drug development against tuberculosis. Structure 2015, 23, 21–33. [Google Scholar] [CrossRef] [Green Version]
- Lu, R.; Schaefer, C.M.; Nesbitt, N.M.; Kuper, J.; Kisker, C.; Sampson, N.S. Catabolism of the cholesterol side chain in Mycobacterium tuberculosis is controlled by a redox-sensitive thiol switch. ACS Infect. Dis. 2017, 3, 666–675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casabon, I.; Swain, K.; Crowe, A.M.; Eltis, L.D.; Mohn, W.W. Actinobacterial acyl coenzyme A synthetases involved in steroid side-chain catabolism. J. Bacteriol. 2014, 196, 579–587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilbert, S.; Hood, L.; Seah, S.Y.K. Characterization of an aldolase involved in cholesterol side chain degradation in Mycobacterium tuberculosis. J. Bacteriol. 2018, 200, e00512-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, S.T.; Sampson, N.S. Mycobacterium tuberculosis utilizes a unique heterotetrameric structure for dehydrogenation of the cholesterol side chain. Biochemistry 2013, 52, 2895–2904. [Google Scholar] [CrossRef] [Green Version]
- Szentirmai, A. Microbial physiology of sidechain degradation of sterols. J. Ind. Microbiol. 1990, 6, 101–115. [Google Scholar] [CrossRef]
- Aggett, R.; Mallette, E.; Gilbert, S.E.; Vachon, M.A.; Schroeter, K.L.; Kimber, M.S.; Seah, S.Y.K. The steroid side-chain–cleaving aldolase Ltp2–ChsH2DUF35 is a thiolase superfamily member with a radically repurposed active site. J. Biol. Chem. 2019, 294, 11934–11943. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Guja, K.E.; Thomas, S.T.; Garcia-Diaz, M.; Sampson, N.S. A distinct MaoC-like enoyl-CoA hydratase architecture mediates cholesterol catabolism in Mycobacterium tuberculosis. ACS Chem. Biol. 2014, 9, 2632–2645. [Google Scholar] [CrossRef] [Green Version]
- Yuan, T.; Yang, M.; Gehring, K.; Sampson, N.S. Mycobacterium tuberculosis exploits a heterohexameric enoyl-CoA hydratase retro-aldolase complex for cholesterol catabolism. Biochemistry 2019, 58, 4224–4235. [Google Scholar] [CrossRef]
- Park, R.J.; Dunn, N.W.; Ide, J.A.; Park, J.; Dunn, N.W.; Ideb, J.A.; Park, R.J.; Dunn, N.W.; Ide, J.A. A catecholic 9,10-seco steroid as a product of aerobic catabolism of cholic acid by a Pseudomonas sp. Steroids 1986, 48, 439–450. [Google Scholar] [CrossRef]
- Van Der Geize, R.; Hessels, G.I.; Van Gerwen, R.; Van Der Meijden, P.; Dijkhuizen, L. Molecular and functional characterization of kshA and kshB, encoding two components of 3-ketosteroid 9α-hydroxylase, a class IA monooxygenase, in Rhodococcus erythropolis strain SQ1. Mol. Microbiol. 2002, 45, 1007–1018. [Google Scholar] [CrossRef]
- Petrusma, M.; Dijkhuizen, L.; Van Der Geize, R. Rhodococcus rhodochrous DSM 43269 3-ketosteroid 9α-hydroxylase, a two-component iron-sulfur-containing monooxygenase with subtle steroid substrate specificity. Appl. Environ. Microbiol. 2009, 75, 5300–5307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrusma, M.; Hessels, G.; Dijkhuizen, L.; van der Geize, R. Multiplicity of 3-ketosteroid-9α-hydroxylase enzymes in Rhodococcus rhodochrous DSM43269 for specific degradation of different classes of steroids. J. Bacteriol. 2011, 193, 3931–3940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Penfield, J.S.; Worrall, L.J.; Strynadka, N.C.; Eltis, L.D. Substrate specificities and conformational flexibility of 3-ketosteroid 9α-hydroxylases. J. Biol. Chem. 2014, 289, 25523–25536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibero, J.; Galán, B.; Díaz, E.; García, J.L. Testosterone degradative pathway of Novosphingobium tardaugens. Genes 2019, 10, 871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dagley, S. Catabolism of aromatic compounds by micro-organisms. Adv. Microbial. Physiol. 1971, 6, 1–46. [Google Scholar]
- Horinouchi, M.; Hayashi, T.; Kudo, T. The genes encoding the hydroxylase of 3-hydroxy-9,10-secoandrosta-1,3,5(10) -triene-9,17-dione in steroid degradation in Comamonas testosteroni TA441. J. Steroid Biochem. Mol. Biol. 2004, 92, 143–154. [Google Scholar] [CrossRef] [PubMed]
- Horinouchi, M.; Yamamoto, T.; Taguchi, K.; Arai, H.; Kudo, T. Meta-cleavage enzyme gene tesB is necessary for testosterone degradation in Comamonas testosteroni TA441. Microbiology 2001, 147, 3367–3375. [Google Scholar] [CrossRef] [Green Version]
- Yam, K.C.; D’Angelo, I.; Kalscheuer, R.; Zhu, H.; Wang, J.X.; Snieckus, V.; Ly, L.H.; Converse, P.J.; Jacobs, W.R.; Strynadka, N.; et al. Studies of a ring-cleaving dioxygenase illuminate the role of cholesterol metabolism in the pathogenesis of Mycobacterium tuberculosis. PLoS Pathog. 2009, 5, e1000344. [Google Scholar] [CrossRef]
- Van der Geize, R.; Yam, K.; Heuser, T.; Wilbrink, M.H.; Hara, H.; Anderton, M.C.; Sim, E.; Dijkhuizen, L.; Davies, J.E.; Mohn, W.W.; et al. A gene cluster encoding cholesterol catabolism in a soil Actinomycete provides insight into Mycobacterium tuberculosis survival in macrophages. Proc. Natl. Acad. Sci. USA 2007, 104, 1947–1952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horinouchi, M.; Hayashi, T.; Koshino, H.; Yamamoto, T.; Kudo, T. Gene encoding the hydrolase for the product of the meta-cleavage reaction in testosterone degradation by Comamonas testosteroni. Appl. Environ. Microbiol. 2003, 69, 2139–2152. [Google Scholar] [CrossRef] [Green Version]
- Seah, S.Y.K.; Ke, J.; Denis, G.; Horsman, G.P.; Fortin, P.D.; Whiting, C.J.; Eltis, L.D. Characterization of a C-C bond hydrolase from Sphingomonas wittichii RW1 with novel specificities towards polychlorinated biphenyl metabolites. J. Bacteriol. 2007, 189, 4038–4045. [Google Scholar] [CrossRef] [Green Version]
- Lack, N.; Lowe, E.D.; Liu, J.; Eltis, L.D.; Noble, M.E.M.; Sim, E.; Westwood, I.M. Structure of HsaD, a steroid-degrading hydrolase, from Mycobacterium tuberculosis. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2007, 64, 2–7. [Google Scholar] [CrossRef] [Green Version]
- Lack, N.A.; Yam, K.C.; Lowe, E.E.; Horsman, G.P.; Owen, R.L.; Sim, E.; Eltis, L.D. Characterization of a carbon-carbon hydrolase from Mycobacterium tuberculosis involved in cholesterol metabolism. J. Biol. Chem. 2010, 285, 434–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horinouchi, M.; Hayashi, T.; Koshino, H.; Kurita, T.; Kudo, T. Identification of 9,17-dioxo-1,2,3,4,10,19-hexanorandrostan-5-oic acid, 4-hydroxy-2-oxohexanoic acid, and 2-hydroxyhexa-2,4-dienoic acid and related enzymes involved in testosterone degradation in Comamonas testosteroni TA441. Appl. Environ. Microbiol. 2005, 71, 5275–5281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carere, J.; McKenna, S.E.; Kimber, M.S.; Seah, S.Y.K. Characterization of an aldolase-dehydrogenase complex from the cholesterol degradation pathway of Mycobacterium tuberculosis. Biochemistry 2013, 52, 3502–3511. [Google Scholar] [CrossRef]
- Fahrbach, M.; Krauss, M.; Preiss, A.; Kohler, H.P.E.; Hollender, J. Anaerobic testosterone degradation in Steroidobacter denitrificans-identification of transformation products. Environ. Pollut. 2010, 158, 2572–2581. [Google Scholar] [CrossRef] [PubMed]
- Fahrbach, M.; Kuever, J.; Remesch, M.; Huber, B.E.; Kämpfer, P.; Dott, W.; Hollender, J. Steroidobacter denitrificans gen. nov., sp. nov., a steroidal hormone-degrading γ-Proteobacterium. Int. J. Syst. Evol. Microbiol. 2008, 58, 2215–2223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tarlera, S.; Denner, E.B.M. Sterolibacterium denitrificans gen. nov., sp. nov., a novel cholesterol-oxidizing, denitrifying member of the β-Proteobacteria. Int. J. Syst. Evol. Microbiol. 2003, 53, 1085–1091. [Google Scholar] [CrossRef] [PubMed]
- Shih, C.-J.; Chen, Y.-L.; Wang, C.-H.; Wei, S.T.-S.; Lin, I.-T.; Ismail, W.A.; Chiang, Y.-R. Biochemical mechanisms and microorganisms involved in anaerobic testosterone metabolism in estuarine sediments. Front. Microbiol. 2017, 8, 1520. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.H.; Yu, C.P.; Lee, T.H.; Lin, C.W.; Ismail, W.; Wey, S.P.; Kuo, A.T.; Chiang, Y.R. Anoxic androgen degradation by the denitrifying bacterium Sterolibacterium denitrificans via the 2,3-seco pathway. Appl. Environ. Microbiol. 2014, 80, 3442–3452. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.-H.; Leu, Y.-L.; Ismail, W.; Tang, S.-L.; Tsai, C.-Y.; Chen, H.-J.; Kao, A.-T.; Chiang, Y.-R. Anaerobic and aerobic cleavage of the steroid core ring structure by Steroidobacter denitrificans. J. Lipid Res. 2013, 54, 1493–1504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, F.-C.; Chen, Y.-L.; Tang, S.-L.; Yu, C.-P.; Wang, P.-H.; Ismail, W.; Wang, C.-H.; Ding, J.-Y.; Yang, C.-Y.C.-Y.; Yang, C.-Y.C.-Y.; et al. Integrated multi-omics analyses reveal the biochemical mechanisms and phylogenetic relevance of anaerobic androgen biodegradation in the environment. ISME J. 2016, 10, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crowe, A.M.; Casabon, I.I.; Brown, K.L.; Liu, J.; Lian, J.; Rogalski, J.C.; Hurst, T.E.; Snieckus, V.; Foster, L.J.; Eltis, L.D. Catabolism of the last two steroid rings in Mycobacterium tuberculosis and other bacteria. mBio 2017, 8, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Gadbery, J.; Round, J.; Yuan, T.; Wipperman, M.F.; Story, K.T.; Crowe, A.; Casabon, I.; Liu, J.; Yang, X.; Eltis, L.D.; et al. IpdE1-IpdE2 is a heterotetrameric acyl coenzyme A dehydrogenase that is widely distributed in steroid-degrading bacteria. Biochemistry 2020, 59, 1113–1123. [Google Scholar] [CrossRef]
- Horinouchi, M.; Hayashi, T.; Koshino, H.; Malon, M.; Hirota, H.; Kudo, T. Identification of 9α-hydroxy-17-oxo-1,2,3,4,10,19-hexanorandrostan- 5-oic acid in steroid degradation by Comamonas testosteroni TA441 and its conversion to the corresponding 6-en-5-oyl coenzyme A (CoA) involving open reading frame 28 (ORF28)- and ORF30-encoded acyl-CoA dehydrogenases. J. Bacteriol. 2014, 196, 3598–3608. [Google Scholar] [PubMed] [Green Version]
- Horinouchi, M.; Koshino, H.; Malon, M.; Hirota, H.; Hayashi, T. Steroid degradation in Comamonas testosteroni TA441: Identification of metabolites and the genes involved in the reactions necessary before D-ring cleavage. Appl. Environ. Microbiol. 2018, 84, e01324-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horinouchi, M.; Malon, M.; Hirota, H.; Hayashi, T. Identification of 4-methyl-5-oxo-octane-1,8-dioic acid and the derivatives as metabolites of steroidal C,D-ring degradation in Comamonas testosteroni TA441. J. Steroid Biochem. Mol. Biol. 2019, 185, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Horinouchi, M.; Koshino, H.; Malon, M.; Hirota, H.; Hayashi, T. Identification of 9-oxo-1,2,3,4,5,6,10,19-octanor-13,17-secoandrost-8(14)-ene-7,17-dioic acid as a metabolite of steroid degradation in Comamonas testosteroni TA441 and the genes involved in the conversion. J. Steroid Biochem. Mol. Biol. 2019, 185, 268–276. [Google Scholar] [CrossRef] [PubMed]
- Helenius, A.; Simons, K. Solubilization of membranes by detergents. BBA Rev. Biomembr. 1975, 415, 29–79. [Google Scholar] [CrossRef]
- Begley, M.; Gahan, C.G.M.; Hill, C. The interaction between bacteria and bile. FEMS Microbiol. Rev. 2005, 29, 625–651. [Google Scholar] [CrossRef] [Green Version]
- Bernstein, C.; Bernstein, H.; Payne, C.M.; Beard, S.E.; Schneider, J. Bile salt activation of stress response promoters in Escherichia coli. Curr. Microbiol. 1999, 39, 68–72. [Google Scholar] [CrossRef] [PubMed]
- Cremers, C.M.; Knoefler, D.; Vitvitsky, V.; Banerjee, R.; Jakob, U. Bile salts act as effective protein-unfolding agents and instigators of disulfide stress in vivo. Proc. Natl. Acad. Sci. USA 2014, 111, E1610–E1619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gunn, J.S. Mechanisms of bacterial resistance and response to bile. Microbes Infect. 2000, 2, 907–913. [Google Scholar] [CrossRef]
- Nikaido, H. Molecular basis of bacterial outer membrane permeability revisited. Microbiol. Mol. Biol. Rev. 2003, 67, 593–656. [Google Scholar] [CrossRef] [Green Version]
- Hancock, R.E.W. The bacterial outer membrane as a drug barrier. Trends Microbiol. 1997, 5, 37–42. [Google Scholar] [CrossRef]
- Paul, S.; Alegre, K.O.; Holdsworth, S.R.; Rice, M.; Brown, J.A.; Mcveigh, P.; Kelly, S.M.; Law, C.J. A single-component multidrug transporter of the major facilitator superfamily is part of a network that protects Escherichia coli from bile salt stress. Mol. Microbiol. 2014, 92, 872–884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, C.Y.; Liu, W.H. Production of androsta-1,4-diene-3,17-dione from cholesterol using immobilized growing cells of Mycobacterium sp. NRRL B-3683 adsorbed on solid carriers. Appl. Microbiol. Biotechnol. 1992, 36, 598–603. [Google Scholar] [CrossRef]
- Perez, C.; Falero, A.; Llanes, N.; Hung, B.R.; Hervé, M.E.; Palmero, A.; Martí, E. Resistance to androstanes as an approach for androstandienedione yield enhancement in industrial mycobacteria. J. Ind. Microbiol. Biotechnol. 2003, 30, 623–626. [Google Scholar] [CrossRef]
- Göhler, A.; Xiong, G.; Paulsen, S.; Trentmann, G.; Maser, E. Testosterone-inducible regulator is a kinase that drives steroid sensing and metabolism in Comamonas testosteroni. J. Biol. Chem. 2008, 283, 17380–17390. [Google Scholar] [CrossRef] [Green Version]
- Gong, W.; Xiong, G.; Maser, E. Oligomerization and negative autoregulation of the LysR-type transcriptional regulator HsdR from Comamonas testosteroni. J. Steroid Biochem. Mol. Biol. 2012, 132, 203–211. [Google Scholar] [CrossRef]
- Kendall, S.L.; Burgess, P.; Balhana, R.; Withers, M.; Ten Bokum, A.; Lott, J.S.; Gao, C.; Uhia-Castro, I.; Stoker, N.G. Cholesterol utilization in mycobacteria is controlled by two TetR-type transcriptional regulators: kstR and kstR2. Microbiology 2010, 156, 1362–1371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Fernández, E.; Medrano, F.J.; Galán, B.; García, J.L. Deciphering the transcriptional regulation of cholesterol catabolic pathway in mycobacteria: Identification of the inducer of KstR repressor. J. Biol. Chem. 2014, 289, 17576–17588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casabon, I.; Zhu, S.H.; Otani, H.; Liu, J.; Mohn, W.W.; Eltis, L.D. Regulation of the KstR2 regulon of Mycobacterium tuberculosis by a cholesterol catabolite. Mol. Microbiol. 2013, 89, 1201–1212. [Google Scholar] [CrossRef]
- Taoka, H.; Yokoyama, Y.; Morimoto, K.; Kitamura, N.; Tanigaki, T.; Takashina, Y.; Tsubota, K.; Watanabe, M. Role of bile acids in the regulation of the metabolic pathways. World J. Diabetes July World J. Diabetes 2016, 10, 260–270. [Google Scholar] [CrossRef]
- Chiang, J.Y.L. Bile acid metabolism and signaling. Compr. Physiol. 2013, 3, 1191–1212. [Google Scholar]
- Li, W. Bile acid secreted by male sea lamprey that acts as a sex pheromone. Science 2002, 296, 138–141. [Google Scholar] [CrossRef] [Green Version]
- Li, K.; Scott, A.M.; Brant, C.O.; Fissette, S.D.; Riedy, J.J.; Hoye, T.R.; Li, W. Bile salt-like dienones having a novel skeleton or a rare substitution pattern function as chemical cues in adult sea lamprey. Org. Lett. 2017, 19, 4444–4447. [Google Scholar] [CrossRef] [PubMed]
- Brant, C.O.; Chung-Davidson, Y.-W.; Li, K.; Scott, A.M.; Li, W. Biosynthesis and release of pheromonal bile salts in mature male sea lamprey. BMC Biochem. 2013, 14, 30. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Zhang, Q.; Dexheimer, T.S.; Ren, J.; Neubig, R.R.; Li, W. Two highly related odorant receptors specifically detect a-bile acid pheromones in sea lamprey (Petromyzon marinus). J. Biol. Chem. 2020, 295, 12153–12166. [Google Scholar] [CrossRef] [PubMed]
- Hahn, M.A.; Effertz, C.; Bigler, L.; von Elert, E. 5α-cyprinol sulfate, a bile salt from fish, induces diel vertical migration in Daphnia. Elife 2019, 8, e44791. [Google Scholar] [CrossRef]
- Doyle, W.I.; Dinser, J.A.; Cansler, H.L.; Zhang, X.; Dinh, D.D.; Browder, N.S.; Riddington, I.M.; Meeks, J.P. Faecal bile acids are natural ligands of the mouse accessory olfactory system. Nat. Commun. 2016, 7, 11936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, K.; Brant, C.O.; Siefkes, M.J.; Kruckman, H.G.; Li, W. Characterization of a novel bile alcohol sulfate released by sexually mature male sea lamprey (Petromyzon marinus). PLoS ONE 2013, 8, e68157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulenburg, H.; Félix, M.A. The natural biotic environment of Caenorhabditis elegans. Genetics 2017, 206, 55–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frézal, L.; Félix, M.A. C. elegans outside the Petri dish. Elife 2015, 30, 4. [Google Scholar] [CrossRef] [PubMed]
- Maglich, J.M.; Sluder, A.; Guan, X.; Shi, Y.; McKee, D.D.; Carrick, K.; Kamdar, K.; Willson, T.M.; Moore, J.T. Comparison of complete nuclear receptor sets from the human, Caenorhabditis elegans and Drosophila genomes. Genome Biol. 2001, 2, RESEARCH0029. [Google Scholar] [CrossRef]
- Motola, D.L.; Cummins, C.L.; Rottiers, V.; Sharma, K.K.; Li, T.; Li, Y.; Suino-Powell, K.; Xu, H.E.; Auchus, R.J.; Antebi, A.; et al. Identification of ligands for DAF-12 that govern dauer formation and reproduction in C. elegans. Cell 2006, 124, 1209–1223. [Google Scholar] [CrossRef] [Green Version]
- Aguilaniu, H.; Fabrizio, P.; Witting, M. The role of dafachronic acid signaling in development and longevity in Caenorhabditis elegans: Digging deeper using cutting-edge analytical chemistry. Front. Endocrinol. Lausanne 2016, 7, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Mansell, D.S.; Bryson, R.J.; Harter, T.; Webster, J.P.; Kolodziej, E.P.; Sedlak, D.L. Fate of endogenous steroid hormones in steer feedlots under simulated rainfall-induced runoff. Environ. Sci. Technol 2011, 45, 8811–8818. [Google Scholar] [CrossRef]
- Gravert, T.K.O.; Fauser, P.; Olsen, P.; Hansen, M. In situ formation of environmental endocrine disruptors from phytosterol degradation: A temporal model for agricultural soils. Environ. Sci. Process. Impacts 2021, 23, 855–866. [Google Scholar] [CrossRef]
- Matsumoto, A.M.; Marck, B.T. DEA Agreement No. DEA-04- P0007 Final Report [Analysis of the Androgenic and Anabolic Activities of 1,4-Androstadien-3,17-dione and 19-nor-4,9(10)- Androstadienedione in Male Sprague Dawley Rats]. DEA Document ID DEA-2008-0007-0003. 2006. Available online: https://www.regulations.gov/document/DEA-2008-0007-0003 (accessed on 18 August 2017).
- Bhasin, S.; Jasuja, R. Pharmacological Analysis of Boldione and 19-nor-4,9(10)-Androstadienedione for Androgenic Activity Using C3H10T1/2 Stem Cells. DEA Document ID DEA-2008-0007-0002. 2005. Available online: https://www.regulations.gov/document/DEA-2008-0007-0002 (accessed on 18 August 2017).
- Barceloux, D.G.; Palmer, R.B. Anabolic—Androgenic steroids. Disease-a-Month 2013, 59, 226–248. [Google Scholar] [CrossRef]
- Kicman, A. Pharmacology of anabolic steroids. Br. J. Pharmacol. 2008, 154, 502–521. [Google Scholar] [CrossRef]
- Hou, L.; Xie, Y.; Ying, G.; Fang, Z. Developmental and reproductive characteristics of western mosquitofish (Gambusia affinis) exposed to paper mill effluent in the Dengcun River, Sihui, South China. Aquat. Toxicol. 2011, 103, 140–149. [Google Scholar] [CrossRef] [PubMed]
- Brockmeier, E.K.; Jayasinghe, B.S.; Pine, W.E.; Wilkinson, K.A.; Denslow, N.D. Exposure to paper mill effluent at a site in north central Florida elicits molecular-level changes in gene expression indicative of progesterone and androgen exposure. PLoS ONE 2014, 9, e106644. [Google Scholar] [CrossRef] [PubMed]
- Parks, L.G.; Lambright, C.S.; Orlando, E.F.; Guillette, L.J.; Ankley, G.T.; Gray, L.E. Masculinization of female mosquitofish in kraft mill effluent-contaminated fenholloway river water is associated with androgen receptor agonist activity. Toxicol. Sci. 2001, 62, 257–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carson, J.; Jenkins, R.; Wilson, E.; Howell, W.; Moore, R. Naturally occurring progesterone in Loblolly pine: A major steroid precursor of environmental androgens. Environ. Toxicol. Chem. 2007, 27, 1273–1278. [Google Scholar] [CrossRef]
- Dlugovitzky, D.G.; Fontela, M.S.; Martinel Lamas, D.J.; Valdez, R.A.; Romano, M.C. Mycobacterium smegmatis synthesizes in vitro androgens and estrogens from different steroid precursors. Can. J. Microbiol. 2015, 61, 451–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jenkins, R.L.; Wilson, E.M.; Angus, R.A.; Howell, W.M.; Kirk, M.; Moore, R.; Nance, M.; Brown, A. Production of androgens by microbial transformation of progesterone in vitro: A model for androgen production in rivers receiving paper mill effluent. Environ. Health Perspect. 2004, 112, 1508–1511. [Google Scholar] [CrossRef] [Green Version]
- Enmark, E.; Gustafsson, J.Å. Nematode genome sequence dramatically extends the nuclear receptor superfamily. Trends Pharmacol. Sci. 2000, 21, 85–87. [Google Scholar] [CrossRef]
- Sluder, A.E.; Mathews, S.W.; Hough, D.; Yin, V.P.; Maina, C.V. The nuclear receptor superfamily has undergone extensive proliferation and diversification in nematodes. Genome Res. 1999, 9, 103–120. [Google Scholar]
- Taubert, S.; Ward, J.D.; Yamamoto, K.R. Nuclear hormone receptors in nematodes: Evolution and function. Mol. Cell. Endocrinol. 2011, 334, 49–55. [Google Scholar] [CrossRef] [Green Version]
- Donova, M.V.; Egorova, O.V. Microbial steroid transformations: Current state and prospects. Appl. Microbiol. Biotechnol. 2012, 94, 1423–1447. [Google Scholar] [CrossRef] [PubMed]
- Bortolini, O.; Medici, A.; Poli, S. Biotransformations on steroid nucleus of bile acids. Steroids 1997, 62, 564–577. [Google Scholar] [CrossRef]
- Bouchier, I.A.D. The medical treatment of gallstones. Ann. Rev. Med. 1980, 31, 59–77. [Google Scholar] [CrossRef] [PubMed]
- Beuers, U.; Spengler, U.; Kruis, W.; Aydemir, Ü.; Wiebecke, B.; Heldwein, W.; Weinzierl, M.; Pape, G.R.; Sauerbruch, T.; Paumgartner, G. Ursodeoxycholic acid for treatment of primary sclerosing cholangitis: A placebo-controlled trial. Hepatology 1992, 16, 707–714. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.; Cho, J.H.; Kim, E.; Kim, Y.J. Ursodeoxycholic acid inhibits the proliferation of colon cancer cells by regulating oxidative stress and cancer stem-like cell growth. PLoS ONE 2017, 12, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Braun, M.; Sun, B.; Anselment, B.; Weuster-Botz, D. Novel whole-cell biocatalysts with recombinant hydroxysteroid dehydrogenases for the asymmetric reduction of dehydrocholic acid. Appl. Microbiol. Biotechnol. 2012, 95, 1457–1468. [Google Scholar] [CrossRef]
- Bordoni, A.; Amaretti, A.; Leonardi, A.; Boschetti, E.; Danesi, F.; Matteuzzi, D.; Roncaglia, L.; Raimondi, S.; Rossi, M. Cholesterol-lowering probiotics: In vitro selection and in vivo testing of bifidobacteria. Appl. Microbiol. Biotechnol. 2013, 97, 8273–8281. [Google Scholar] [CrossRef]
- Ooi, L.G.; Liong, M.T. Cholesterol-lowering effects of probiotics and prebiotics: A review of in vivo and in vitro findings. Int. J. Mol. Sci. 2010, 11, 2499–2522. [Google Scholar] [CrossRef]
- Kumar, R.; Grover, S.; Batish, V.K. Hypocholesterolaemic effect of dietary inclusion of two putative probiotic bile salt hydrolase-producing Lactobacillus plantarum strains in Sprague–Dawley rats. Br. J. Nutr. 2011, 105, 561–573. [Google Scholar] [CrossRef] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feller, F.M.; Holert, J.; Yücel, O.; Philipp, B. Degradation of Bile Acids by Soil and Water Bacteria. Microorganisms 2021, 9, 1759. https://doi.org/10.3390/microorganisms9081759
Feller FM, Holert J, Yücel O, Philipp B. Degradation of Bile Acids by Soil and Water Bacteria. Microorganisms. 2021; 9(8):1759. https://doi.org/10.3390/microorganisms9081759
Chicago/Turabian StyleFeller, Franziska Maria, Johannes Holert, Onur Yücel, and Bodo Philipp. 2021. "Degradation of Bile Acids by Soil and Water Bacteria" Microorganisms 9, no. 8: 1759. https://doi.org/10.3390/microorganisms9081759