
No Metagenomic Evidence of Causative Viral Pathogens in Postencephalitic Parkinsonism Following Encephalitis Lethargica

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients and Available Materials
2.2. Immunohistochemistry from Archived Brain Tissues
2.3. Sequence-Independent Amplification and Next-Generation Sequencing of FFPE Tissue Samples
3. Results
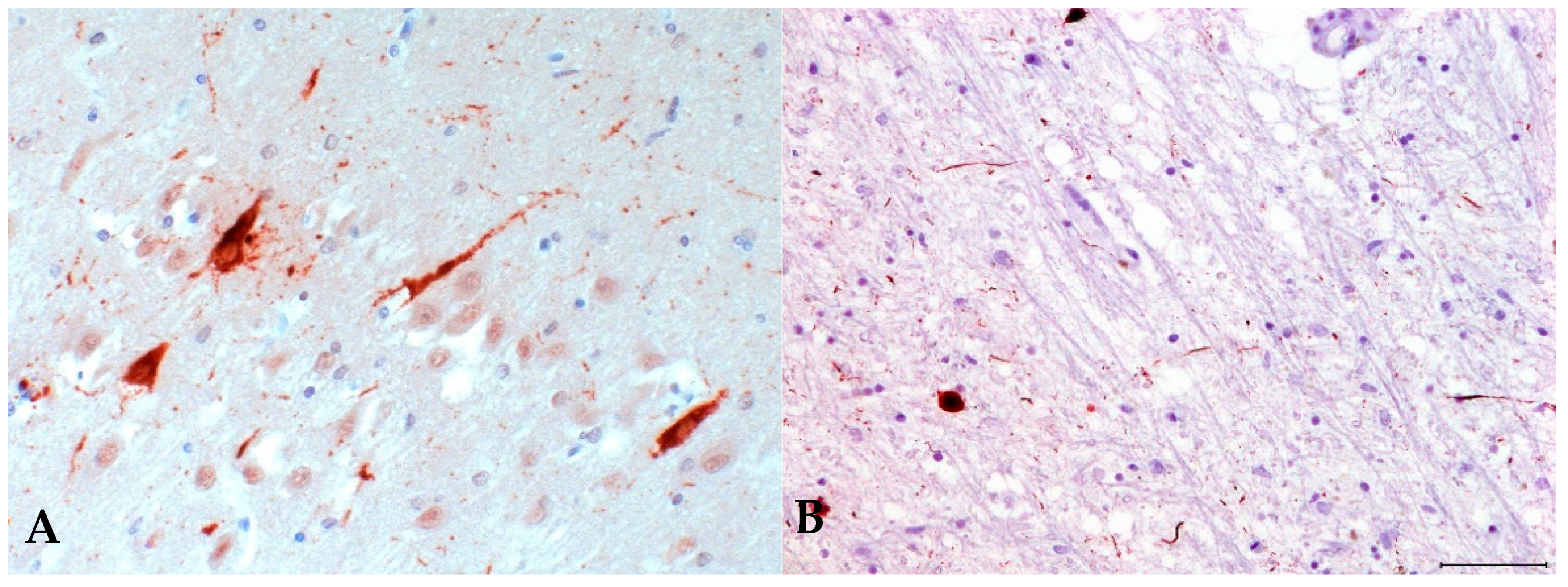
3.1. Immunohistochemistry for Postencephalitic Parkinsonism (PEP)
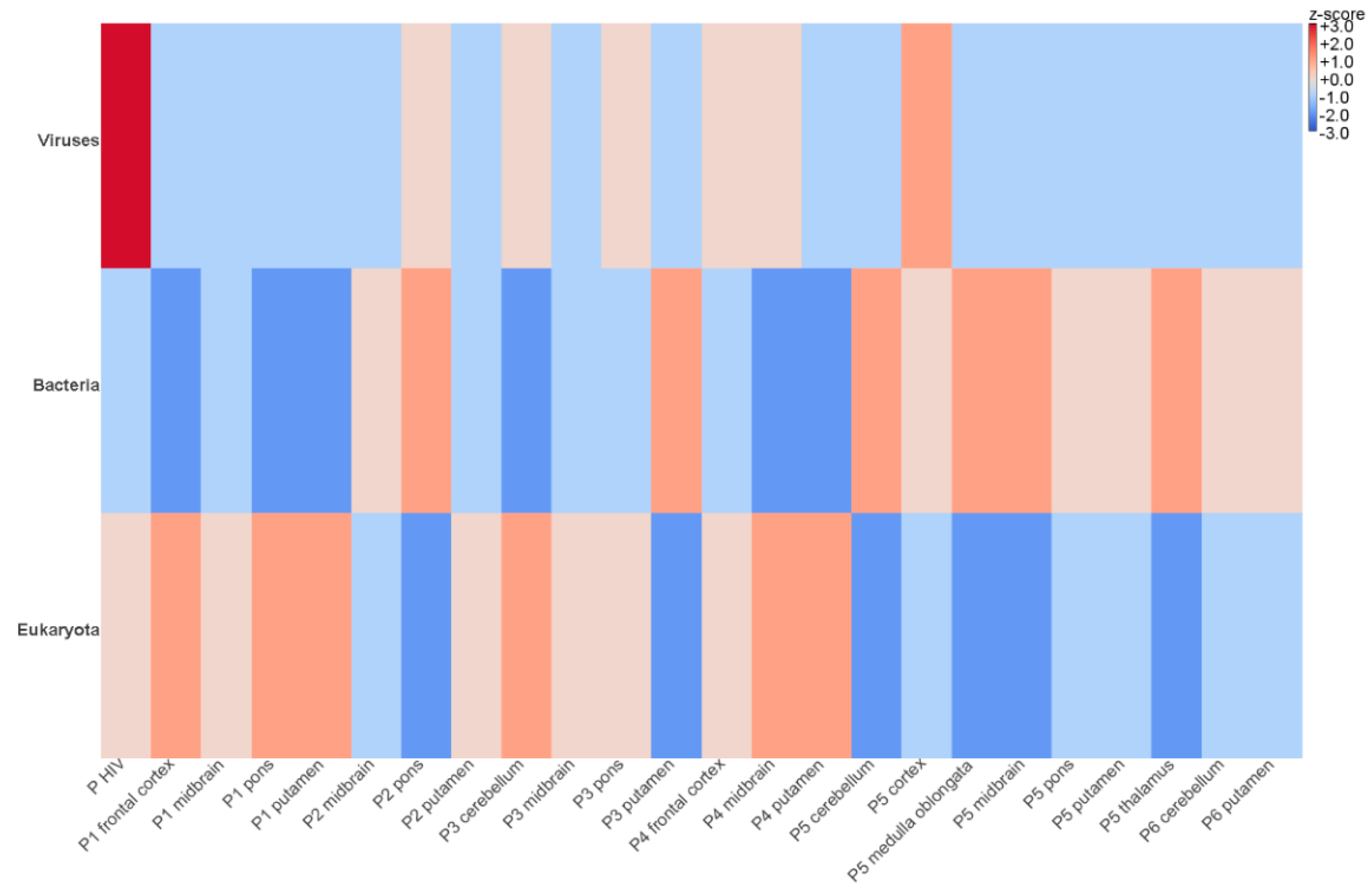
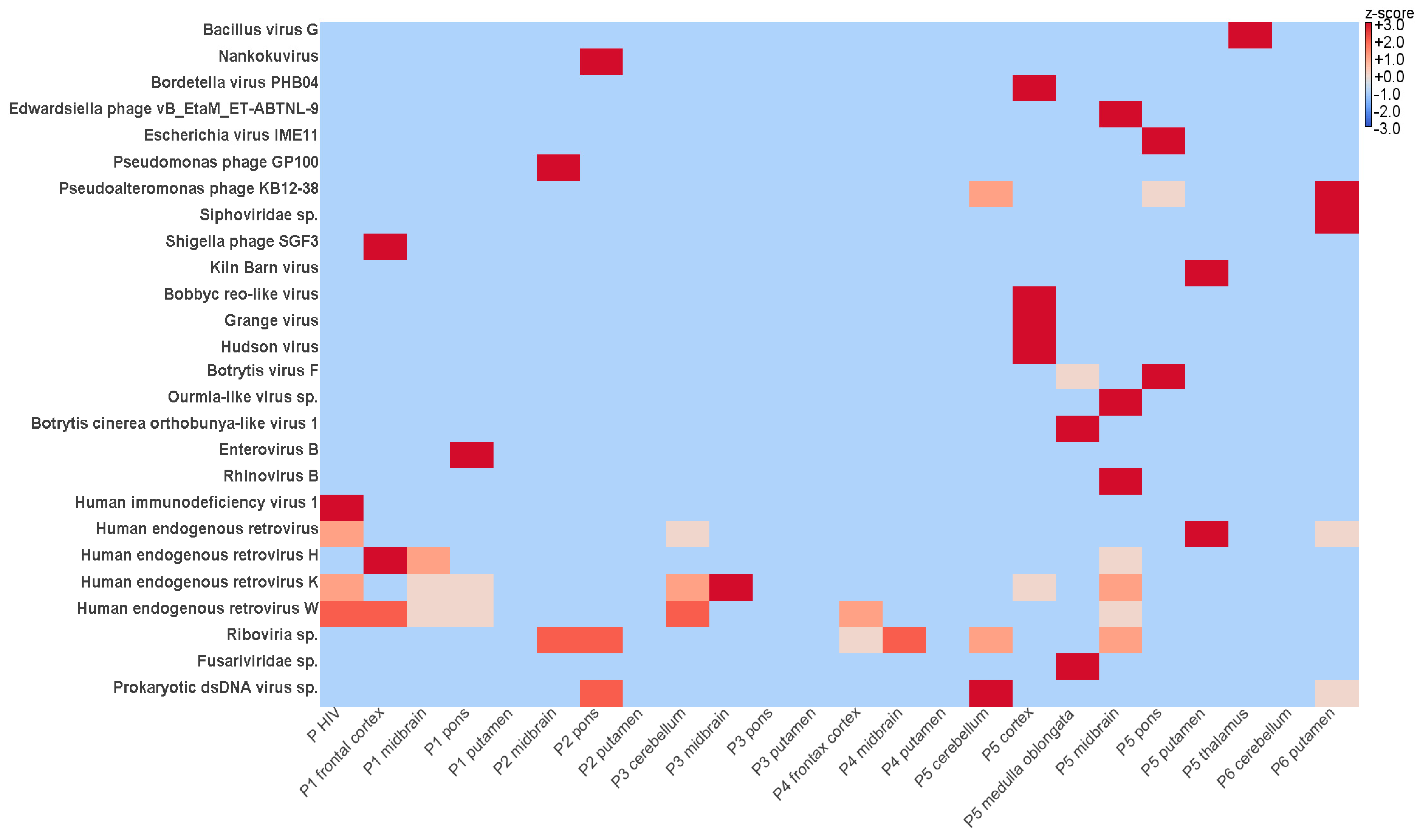
3.2. Identification of Viral Sequences in FFPE Samples Using NGS
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Tappe, D.; Alquezar-Planas, D.E. Medical and molecular perspectives into a forgotten epidemic: Encephalitis lethargica, viruses, and high-throughput sequencing. J. Clin. Virol. 2014, 61, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Vilensky, J.A.; Gilman, S.; McCall, S. A historical analysis of the relationship between encephalitis lethargica and postencephalitic parkinsonism: A complex rather than a direct relationship. Mov. Disord. 2010, 25, 1116–1123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vilensky, J.A.; Gilman, S.; McCall, S. Does the historical literature on encephalitis lethargica support a simple (direct) relationship with postencephalitic Parkinsonism? Mov. Disord. 2010, 25, 1124–1130. [Google Scholar] [CrossRef] [PubMed]
- Jellinger, K.A. Postencephalitic Parkinsonism. In Neurodegeneration: The Molecular Pathology of Dementia and Movement Disorders, 2nd ed.; Dickson, D.W., Weller, R.O., Eds.; Wiley: New York, NY, USA, 2011. [Google Scholar]
- Jellinger, K.A. Absence of α-synuclein pathology in postencephalitic parkinsonism. Acta Neuropathol. 2009, 118, 371–379. [Google Scholar] [CrossRef]
- Josephs, K.A.; Parisi, J.E.; Dickson, D.W. Alpha-synuclein studies are negative in postencephalic parkinsonism of von Economo. Neurology 2002, 59, 645. [Google Scholar] [CrossRef]
- Ludlow, M.; Kortekaas, J.; Herden, C.; Hoffmann, B.; Tappe, D.; Trebst, C.; Griffin, D.E.; Brindle, H.; Solomon, T.; Brown, A.S.; et al. Neurotropic virus infections as the cause of immediate and delayed neuropathology. Acta Neuropathol. 2016, 131, 159–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffmann, B.; Tappe, D.; Herden, C.; Boldt, A.; Niederstraßer, O.; Müller, T.; Van Der Grinten, E.; Lutter, C.; Abendroth, B.; Teifke, J.P.; et al. A Variegated Squirrel Bornavirus Associated with Fatal Human Encephalitis. N. Engl. J. Med. 2015, 373, 154–162. [Google Scholar] [CrossRef]
- Schlottau, K.; Forth, L.; Angstwurm, K.; Höper, D.; Zecher, D.; Liesche, F.; Hoffmann, B.; Kegel, V.; Seehofer, D.; Platen, S.; et al. Fatal Encephalitic Borna Disease Virus 1 in Solid-Organ Transplant Recipients. N. Engl. J. Med. 2018, 379, 1377–1379. [Google Scholar] [CrossRef]
- Casals, J.; Elizan, T.S.; Yahr, M.D. Postencephalitic parkinsonism—A review. J. Neural Transm. 1998, 105, 645–676. [Google Scholar] [CrossRef]
- Jang, H.; Boltz, D.A.; Webster, R.G.; Smeyne, R.J. Viral parkinsonism. Biochim. Biophys. Acta 2009, 1792, 714–721. [Google Scholar] [CrossRef] [Green Version]
- Bodewes, R.; van Run, P.R.; Schürch, A.C.; Koopmans, M.P.; Osterhaus, A.D.; Baumgärtner, W.; Kuiken, T.; Smits, S.L. Virus characterization and discovery in formalin-fixed paraffin-embedded tissues. J. Virol. Methods 2015, 214, 54–59. [Google Scholar] [CrossRef] [PubMed]
- Worobey, M.; Gemmel, M.; Teuwen, D.E.; Haselkorn, T.; Kunstman, K.; Bunce, M.; Muyembe, J.-J.; Kabongo, J.-M.M.; Kalengayi, R.M.; Van Marck, E.; et al. Direct evidence of extensive diversity of HIV-1 in Kinshasa by 1960. Nature 2008, 455, 661–664. [Google Scholar] [CrossRef] [Green Version]
- Taubenberger, J.K.; Reid, A.H.; Krafft, A.E.; Bijwaard, K.E.; Fanning, T.G. Initial genetic characterization of the 1918 “Spanish” influenza virus. Science 1997, 275, 1793–1796. [Google Scholar] [CrossRef] [Green Version]
- McCall, S.; Henry, J.M.; Reid, A.; Taubenberger, J.K. Influenza RNA not Detected in Archival Brain Tissues from Acute Encephalitis Lethargica Cases or in postencephalitic parkinson Cases. J. Neuropathol. Exp. Neurol. 2001, 60, 696–704. [Google Scholar] [CrossRef] [Green Version]
- Lo, K.C.; Geddes, J.F.; Daniels, R.S.; Oxford, J.S. Lack of detection of influenza genes in archived formalin-fixed, paraffin wax-embedded brain samples of encephalitis lethargica patients from 1916 to 1920. Virchows Arch. 2003, 442, 591–596. [Google Scholar] [CrossRef]
- Cadar, D.; Allendorf, V.; Schulze, V.; Ulrich, R.G.; Schlottau, K.; Ebinger, A.; Hoffmann, B.; Hoffmann, D.; Rubbenstroth, D.; Ismer, G.; et al. Introduction and spread of variegated squirrel bornavirus 1 (VSBV-1) between exotic squirrels and spill-over infections to humans in Germany. Emerg. Microbes Infect. 2021, 10, 602–611. [Google Scholar] [CrossRef]
- Xiao, Y.-L.; Kash, J.C.; Beres, S.; Sheng, Z.-M.; Musser, J.M.; Taubenberger, J.K. High-throughput RNA sequencing of a formalin-fixed, paraffin-embedded autopsy lung tissue sample from the 1918 influenza pandemic. J. Pathol. 2013, 229, 535–545. [Google Scholar] [CrossRef] [Green Version]
- Carrick, D.M.; Mehaffey, M.G.; Sachs, M.; Altekruse, S.; Camalier, C.; Chuaqui, R.; Cozen, W.; Das, B.; Hernandez, B.Y.; Lih, C.-J.; et al. Robustness of Next Generation Sequencing on Older Formalin-Fixed Paraffin-Embedded Tissue. PLoS ONE 2015, 10, e0127353. [Google Scholar] [CrossRef] [Green Version]
- Tomazatos, A.; Marschang, R.E.; Maranda, I.; Baum, H.; Bialonski, A.; Spînu, M.; Lühken, R.; Schmidt-Chanasit, J.; Cadar, D. Letea Virus: Comparative Genomics and Phylogenetic Analysis of a Novel Reassortant Orbivirus Discovered in Grass Snakes (Natrix natrix). Viruses 2020, 12, 243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorenzo, R.H.; Cadar, D.; Koundouno, F.; Juste, J.; Bialonski, A.; Baum, H.; García-Mudarra, J.; Hakamaki, H.; Bencsik, A.; Nelson, E.; et al. Metagenomic Snapshots of Viral Components in Guinean Bats. Microorganisms 2021, 9, 599. [Google Scholar] [CrossRef]
- Riederer, P.; ter Meulen, V. Correction to: Coronaviruses: A challenge of today and a call for extended human postmortem brain analyses. J. Neural Transm. 2021, 127, 1217–1228. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, K.; Liu, F.; Gong, C.-X.; Grundke-Iqbal, I. Tau in Alzheimer Disease and Related Tauopathies. Curr. Alzheimer Res. 2010, 7, 656–664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, J.; Dickson, D.W. Propagation of tau pathology: Hypotheses, discoveries, and yet unresolved questions from experimental and human brain studies. Acta Neuropathol. 2016, 131, 27–48. [Google Scholar] [CrossRef] [PubMed]
- Schafernak, K.; Bigio, E.H. West Nile Virus Encephalomyelitis with Polio-like Paralysis & Nigral Degeneration. Can. J. Neurol. Sci. 2006, 33, 407–410. [Google Scholar]
- Bancher, C.; Leitner, H.; Jellinger, K.; Eder, H.; Setinek, U.; Fischer, P.; Wegiel, J.; Wisniewski, H.M. On the relationship between measles virus and Alzheimer neurofibrillary tangles in subacute sclerosing panencephalitis. Neurobiol. Aging 1996, 17, 527–533. [Google Scholar] [CrossRef]
- Gelpi, E.; Höftberger, R.; Graus, F.; Ling, H.; Holton, J.L.; Dawson, T.; Popovic, M.; Pretnar-Oblak, J.; Högl, B.; Schmutzhard, E.; et al. Neuropathological criteria of anti-IgLON5-related tauopathy. Acta Neuropathol. 2016, 132, 531–543. [Google Scholar] [CrossRef] [Green Version]
- Armangue, T.; Spatola, M.; Vlagea, A.; Mattozzi, S.; Cárceles-Cordon, M.; Martinez-Heras, E.; Llufriu, S.; Muchart, J.; Erro, M.E.; Abraira, L.; et al. Frequency, symptoms, risk factors, and outcomes of autoimmune encephalitis after herpes simplex encephalitis: A prospective observational study and retrospective analysis. Lancet Neurol. 2018, 17, 760–772. [Google Scholar] [CrossRef] [Green Version]
- Ising, C.; Venegas, C.; Zhang, S.; Scheiblich, H.; Schmidt, S.V.; Vieira-Saecker, A.; Schwartz, S.; Albasset, S.; McManus, R.M.; Tejera, D.; et al. NLRP3 inflammasome activation drives tau pathology. Nature 2019, 575, 669–673. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patient | Sex | Age at Time of Death (Year) | Onset of EL, Onset of PEP; Interval | Storage Time Until Analysis | Clinical Symptoms |
|---|---|---|---|---|---|
| 1 | F | 66 (1979) | 1939, 1951; 12 years | 42 years | akinesia, rigidity, oculogyric crises, tremor, hypomimia, gait disorder |
| 2 | F | 77 (1982) | 1921, 1965; 44 years | 39 years | akinesia, rigidity, gait disturbances |
| 3 | M | 66 (1983) | 1932, 1960; 28 years | 38 years | akinesia, rigidity, oculogyric crises, sialorrhea, gait disorder |
| 4 | F | 65 (1977) | 1923, 1933; 10 years | 44 years | akinesia, bradykinesia, rigidity, oculogyric crises |
| 5 | F | 51 (1959) | 1926, 1945; 19 years | 62 years | akinesia, rigidity, oculogyric crises, hypomimia |
| 6 | M | 62 (1970) | 1923, 1950; 27 years | 51 years | akinesia, rigidity, gait disturbances, oculogyric crises |
| 7 * | M | unknown | N/A | 27 years | N/A |
| Patient | Brain Regions Available and Sequenced by NGS | Trimmed Reads | Host Reads | Viral Reads |
|---|---|---|---|---|
| 1 | frontal cortex | 9,517,032 | 3,639,675 | 302 |
| putamen | 8,920,649 | 3,229,635 | 736 | |
| midbrain | 9,718,416 | 3,978,842 | 503 | |
| pons | 8,543,882 | 3,252,964 | 725 | |
| 2 | putamen | 9,565,917 | 3,170,398 | 831 |
| midbrain | 8,840,396 | 2,086,639 | 1136 | |
| pons | 9,638,905 | 1,850,362 | 7613 | |
| 3 | cerebellum | 10,901,531 | 4,228,111 | 1145 |
| putamen | 8,132,530 | 2,063,999 | 1354 | |
| midbrain | 9,522,435 | 4,053,318 | 4406 | |
| pons | 7,959,775 | 3,268,825 | 4540 | |
| 4 | frontal cortex | 10,557,783 | 4,130,625 | 1295 |
| putamen | 10,221,537 | 4,338,290 | 3600 | |
| midbrain | 9,838,675 | 4,584,931 | 1256 | |
| 5 | cortex | 8,452,734 | 1,802,060 | 3984 |
| thalamus | 10,267,166 | 1,340,110 | 1688 | |
| cortex | 9,139,183 | 1,787,936 | 3453 | |
| putamen | 9,534,555 | 1,759,967 | 2344 | |
| midbrain | 12,018,105 | 2,011,537 | 2552 | |
| cerebellum | 11,802,862 | 1,490,561 | 2147 | |
| pons | 12,276,639 | 4,245,357 | 1198 | |
| medulla oblongata | 13,412,314 | 3,597,636 | 3849 | |
| 6 | putamen | 9,080,742 | 1,510,840 | 1235 |
| cerebellum | 11,276,435 | 1,037,770 | 2289 | |
| 7 * | midbrain | 11,345,665 | 1,227,560 | 8435 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cadar, D.; Jellinger, K.A.; Riederer, P.; Strobel, S.; Monoranu, C.-M.; Tappe, D. No Metagenomic Evidence of Causative Viral Pathogens in Postencephalitic Parkinsonism Following Encephalitis Lethargica. Microorganisms 2021, 9, 1716. https://doi.org/10.3390/microorganisms9081716
Cadar D, Jellinger KA, Riederer P, Strobel S, Monoranu C-M, Tappe D. No Metagenomic Evidence of Causative Viral Pathogens in Postencephalitic Parkinsonism Following Encephalitis Lethargica. Microorganisms. 2021; 9(8):1716. https://doi.org/10.3390/microorganisms9081716
Chicago/Turabian StyleCadar, Dániel, Kurt A. Jellinger, Peter Riederer, Sabrina Strobel, Camelia-Maria Monoranu, and Dennis Tappe. 2021. "No Metagenomic Evidence of Causative Viral Pathogens in Postencephalitic Parkinsonism Following Encephalitis Lethargica" Microorganisms 9, no. 8: 1716. https://doi.org/10.3390/microorganisms9081716