
Hematological Changes in Dogs with Visceral Leishmaniasis Are Associated with Increased IFN-γ and TNF Gene Expression Levels in the Bone Marrow

 ,
,  , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals; Samples; and Clinical, Serological and Hematological Analyses
2.2. Histological Analysis
2.3. RNA and DNA Extraction for Molecular Biology
2.4. Detection of Leishmania infantum Burden in the Bone Marrow
2.5. Cytokine Gene Amplification by Quantitative (RTqPCR)
2.6. Gene Expression Levels and Analysis of the Results
2.7. Ethics Approval
3. Results
3.1. General Characteristics of the Dogs
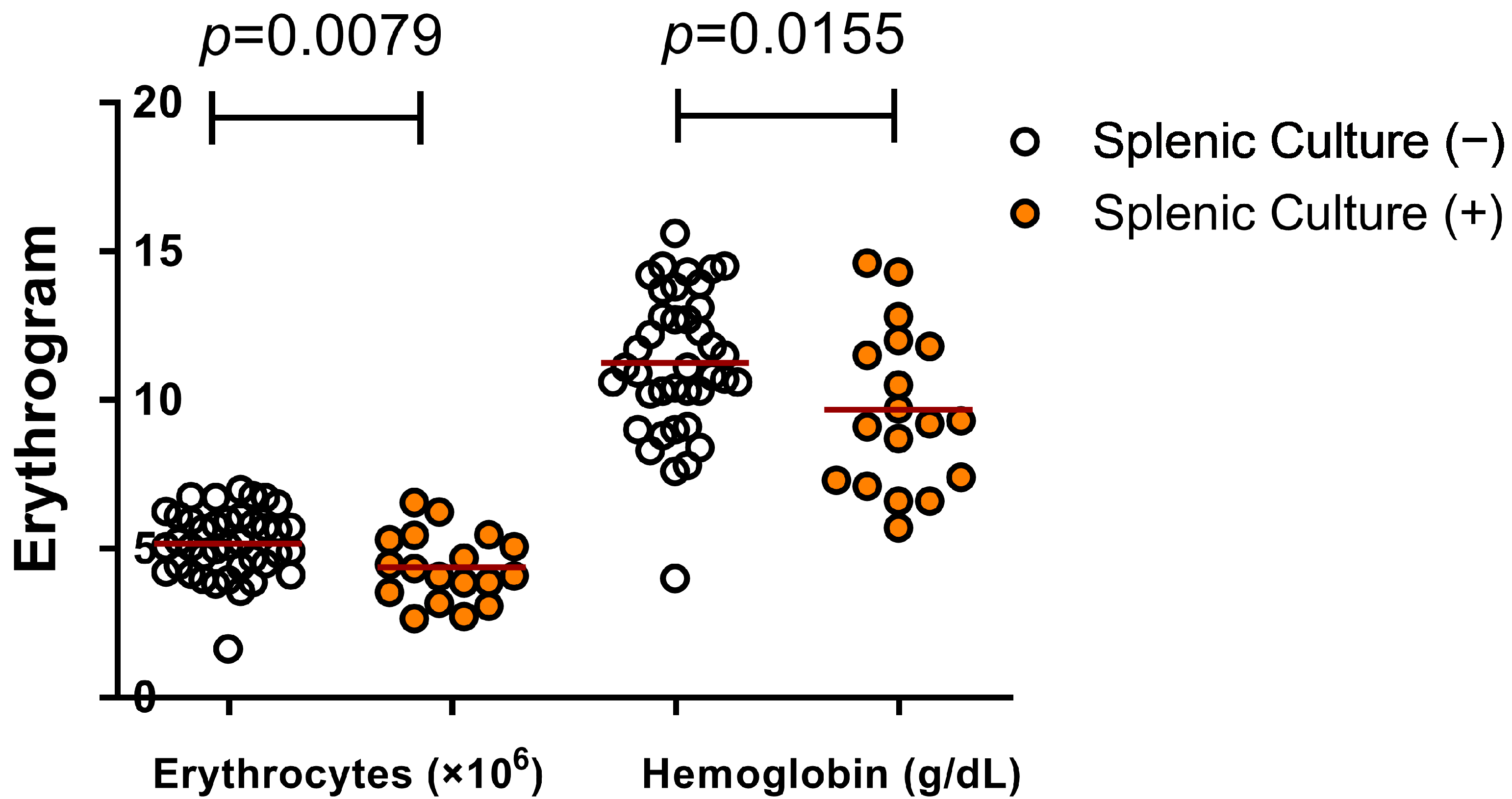
3.2. Active Infection by L. infantum and Changes in Hematological Parameters
3.3. Bone Marrow Changes in Dogs with Active L. infantum Infection
3.4. Hematological Changes in Dogs with Active Infection and Splenic White Pulp Disorganization
3.5. Cytokine Gene Expression Levels in Bone Marrow and Hematological Changes in Dogs Infected with L. infantum
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Desjeux, P. Leishmaniasis: Current situation and new perspectives. Comp. Immunol. Microbiol. Infect. Dis. 2004, 27, 305–318. [Google Scholar] [CrossRef]
- Murray, H.W.; Berman, J.D.; Davies, C.R.; Saravia, N.G. Advances in leishmaniasis. Lancet 2005, 366, 1561–1577. [Google Scholar] [CrossRef]
- WHO World Health Organization. Fact Sheet N°375. Available online: http://www.who.int/mediacentre/factsheets/fs375/en/ (accessed on 20 August 2016).
- Deane, L.M. Epidemiologia e profilaxia do Calazar Americano. Rev. Bras. Malariol. Doencas. Trop. 1956, 10, 431–449. [Google Scholar]
- Marzochi, M.C.A.; Coutinho, S.G.; De Souza, W.J.; De Toledo, L.M.; Grimaldi, J.G.; Momem, H.; Pacheco, R.S.; Sabroza, P.C.; De Souza, M.A.; Rangel, J.F.B. Canine visceral leishmaniasis in Rio de Janeiro, Brazil. Clinical, parasitological, thera-peutical and epidemiological findings (1977–1983). Mem. Inst. Oswaldo Cruz. 1985, 80, 17–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alvar, J.; Cañavate, C.; Molina, R.; Moreno, J.; Nieto, J. Canine Leishmaniasis. Adv. Parasitol. 2004, 57, 1–88. [Google Scholar] [CrossRef]
- Sanchez, M.A.; Diaz, N.L.; Zerpa, O.; Negron, E.; Convit, J.; Tapia, F.J. Organ-specific immunity in canine visceral leish-maniasis: Analysis of symptomatic and asymptomatic dogs naturally infected with Leishmania chagasi. Am. J. Trop. Med. Hyg. 2004, 70, 618–624. [Google Scholar] [CrossRef]
- Reis, A.B.; Martins-Filho, O.A.; Teixeira-Carvalho, A.; Carvalho, M.G.; Mayrink, W.; França-Silva, J.C.; Giunchetti, R.C.; Genaro, O.; Correa-Oliveira, R. Parasite density and impaired biochemical/hematological status are associated with severe clinical aspects of canine visceral leishmaniasis. Res. Veter Sci. 2006, 81, 68–75. [Google Scholar] [CrossRef] [Green Version]
- Baneth, G.; Koutinas, A.F.; Solano-Gallego, L.; Bourdeau, P.; Ferrer, L. Canine leishmaniosis new concepts and insights on an expanding zoonosis: Part one. Trends Parasitol. 2008, 24, 324–330. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, E.; Tumbar, T.; Guasch, G. Socializing with the neighbors: Stem cells and their niche. Cell 2004, 116, 769–778. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Xie, T. STEM CELL NICHE: Structure and Function. Annu. Rev. Cell Dev. Biol. 2005, 21, 605–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valli, V.E.O.; Gentry, P.A. Hematopoietic system. In Pathology of Domestic Animals, 6th ed.; Jubb, K.V.F., Kennedy, P.C., Palmer’s, N.C., Eds.; Elsevier: Amsterdam, The Netherlands, 2007; pp. 107–324. [Google Scholar]
- Robb, L. Cytokine receptors and hematopoietic differentiation. Oncogene 2007, 26, 6715–6723. [Google Scholar] [CrossRef] [Green Version]
- Manzillo, F.V.; Restucci, B.; Pagano, A.; Gradoni, L.; Oliva, G. Pathological changes in the bone marrow of dogs with leish-maniosis. Vet Rec. 2006, 158, 690–694. [Google Scholar] [CrossRef]
- Paranhos-Silva, M.; Oliveira, G.G.; Reis, E.A.; De Menezes, R.M.; Fernandes, O.; Sherlock, I.; Gomes, R.B.; Pontes-De-Carvalho, L.C.; Dos-Santos, W.L. A follow-up of Beagle dogs intradermally infected with Leishmania chagasi in the presence or absence of sand fly saliva. Veter. Parasitol. 2003, 114, 97–111. [Google Scholar] [CrossRef]
- Nicolato, R.D.C.; De Abreu, R.T.; Roatt, B.; Aguiar-Soares, R.D.D.O.; Reis, L.E.S.; Carvalho, M.D.G.; Carneiro, C.M.; Giunchetti, R.C.; Bouillet, L.E.M.; Lemos, D.S.; et al. Clinical Forms of Canine Visceral Leishmaniasis in Naturally Leishmania infantum–Infected Dogs and Related Myelogram and Hemogram Changes. PLoS ONE 2013, 8, e82947. [Google Scholar] [CrossRef] [Green Version]
- Reis, A.B.; Teixeira-Carvalho, A.; Giunchetti, R.C.; Roatt, B.; Coura-Vital, W.; Nicolato, R.D.C.; Silveira-Lemos, D.; Correa-Oliveira, R.; Martins-Filho, O.D.A. Cellular immunophenotypic profile in the splenic compartment during canine visceral leishmaniasis. Veter Immunol. Immunopathol. 2014, 157, 190–196. [Google Scholar] [CrossRef] [Green Version]
- Santana, C.C.; Vassallo, J.; De Freitas, L.A.R.; Oliveira, G.G.S.; Pontes-De-Carvalho, L.C.; Dos-Santos, W.L. Inflammation and structural changes of splenic lymphoid tissue in visceral leishmaniasis: A study on naturally infected dogs. Parasite Immunol. 2008, 30, 515–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lima, I.; Silva, J.S.; Almeida, V.A.; Junior, F.G.L.; Souza, P.A.; Larangeira, D.F.; Moura-Neto, J.P.; Fraga, D.B.; De Freitas, L.A.R.; Dos-Santos, W.L. Severe Clinical Presentation of Visceral Leishmaniasis in Naturally Infected Dogs with Disruption of the Splenic White Pulp. PLoS ONE 2014, 9, e87742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavalcanti, A.S.; Ribeiro-Alves, M.; Pereira, L.O.; Mestre, G.L.; Ferreira, A.B.; Morgado, F.N.; Boité, M.C.; Cupolillo, E.; Moraes, M.O.; Porrozzi, R. Parasite load induces progressive spleen architecture breakage and impairs cytokine mRNA ex-pression in Leishmania infantum-naturally infected dogs. PLoS ONE 2015, 10, e0123009. [Google Scholar] [CrossRef]
- De Abreu, R.T.; Carvalho, M.D.G.; Carneiro, C.M.; Giunchetti, R.C.; Teixeira-Carvalho, A.; Martins-Filho, O.A.; Coura-Vital, W.; Correa-Oliveira, R.; Reis, A.B. Influence of Clinical Status and Parasite Load on Erythropoiesis and Leucopoiesis in Dogs Naturally Infected with Leishmania (Leishmania) chagasi. PLoS ONE 2011, 6, e18873. [Google Scholar] [CrossRef] [Green Version]
- Dos-Santos, W.L.; Jesus, E.; Paranhos-Silva, M.; Pereira, A.; Santos, J.; Baleeiro, C.; Nascimento, E.; Moreira, E.; Oliveira, G.; Pontes-De-Carvalho, L. Associations among immunological, parasitological and clinical parameters in canine visceral leishmaniasis: Emaciation, spleen parasitism, specific antibodies and leishmanin skin test reaction. Veter. Immunol. Immunopathol. 2008, 123, 251–259. [Google Scholar] [CrossRef]
- Weiss, D.J. Histopathology of Canine Nonneoplastic Bone Marrow. Veter. Clin. Pathol. 1986, 15, 7–11. [Google Scholar] [CrossRef]
- Holloway, S.A.; Meyer, D.J.; Mannella, C. Prednisolone and danazol for treatment of immune-mediated anemia, thrombocytopenia, and ineffective erythroid regeneration in a dog. J. Am. Veter. Med. Assoc. 1990, 197, 1045–1048. [Google Scholar]
- Hermida, M.D.-R.; De Melo, C.V.B.; Lima, I.D.S.; Oliveira, G.G.D.S.; Dos-Santos, W.L.C. Histological Disorganization of Spleen Compartments and Severe Visceral Leishmaniasis. Front. Cell. Infect. Microbiol. 2018, 8, 394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maizels, R.M.; Robertson, B.D.; Blaxter, M.L.; Selkirk, M.E. Parasite antigens, parasite genes. In A Laboratory Manual for Molecular Parasitology; Cambridge University Press: Cambridge, UK, 1991. [Google Scholar]
- van Eys Guillaume, J.J.M.; Schoone, G.J.; Kroon, N.C.; Ebeling, S.B. Sequence analysis of small subunit ribosomal RNA genes and its use for detection and identification of Leishmania parasites. Mol. Biochem. Parasitol. 1992, 51, 133–142. [Google Scholar] [CrossRef]
- Carrillo, E.; Ahmed, S.; Goldsmith-Pestana, K.; Nieto, J.; Osorio, Y.; Travi, B.; Moreno, J.; Mc Mahon-Pratt, D. Immunogenici-ty of the P-8 amastigote antigen in the experimental model of canine visceral leishmaniasis. Vaccine 2007, 25, 1534–1543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biswas, T.; Chakraborty, M.; Naskar, K.; Ghosh, D.K.; Ghosal, J. Anemia in Experimental Visceral Leishmaniasis in Hamsters. J. Parasitol. 1992, 78, 140. [Google Scholar] [CrossRef] [PubMed]
- Herwaldt, B.L. Leishmaniasis. Lancet 1999, 354, 1191–1199. [Google Scholar] [CrossRef] [Green Version]
- Varma, N.; Naseem, S. Hematologic Changes in Visceral Leishmaniasis/Kala Azar. Indian J. Hematol. Blood Transfus. 2010, 26, 78–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tryphonas, L.; Zawidzka, Z.; Bernard, M.A.; Janzen, E.A. Visceral leishmaniasis in a dog: Clinical, hematological and pathological observations. Can. J. Comp. Med. Rev. Can. Med. Comp. 1977, 41, 1–12. [Google Scholar]
- Agu, W.E.; Farrell, J.P.; Soulsby, E.J.L. Pathogenesis of anaemia in hamsters infected withLeishmania donovani. Parasitol. Res. 1982, 68, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Lafuse, W.P.; Story, R.; Mahylis, J.; Gupta, G.; Varikuti, S.; Steinkamp, H.; Oghumu, S.; Satoskar, A.R. Leishmania donovani Infection Induces Anemia in Hamsters by Differentially Altering Erythropoiesis in Bone Marrow and Spleen. PLoS ONE 2013, 8, e59509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goto, Y.; Cheng, J.; Omachi, S.; Morimoto, A. Prevalence, severity, and pathogeneses of anemia in visceral leishmaniasis. Parasitol. Res. 2016, 116, 457–464. [Google Scholar] [CrossRef] [PubMed]
- Dos-Santos, W.L.; Pagliari, C.; Santos, L.G.; Almeida, V.A.; Silva, T.L.V.; Coutinho, J.J.; Souza, T.; Duarte, M.I.S.; de Freitas, L.A.; Costa, C.H. A case of conventional treatment failure in visceral leishmaniasis: Leukocyte distribution and cy-tokine expression in splenic compartments. BMC Infect. Dis. 2014, 14, 491. [Google Scholar] [CrossRef]
- Faleiro, R.J.; Kumar, R.; Hafner, L.M.; Engwerda, C.R. Immune Regulation during Chronic Visceral Leishmaniasis. PLoS Negl. Trop. Dis. 2014, 8, e2914. [Google Scholar] [CrossRef] [Green Version]
- Selleri, C.; Maciejewski, J.P.; Sato, T.; Young, N.S. Interferon-gamma constitutively expressed in the stromal microenviron-ment of human marrow cultures mediates potent hematopoietic inhibition. Blood 1996, 87, 4149–4157. [Google Scholar] [CrossRef] [Green Version]
- De Bruin, A.M.; Voermans, C.; Nolte, M.A. Impact of interferon-γ on hematopoiesis. Blood 2014, 124, 2479–2486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiss, G. Iron metabolism in the anemia of chronic disease. Biochim. Biophys. Acta Gen. Subj. 2009, 1790, 682–693. [Google Scholar] [CrossRef] [PubMed]
- Zoller, E.E.; Lykens, J.E.; Terrell, C.E.; Aliberti, J.; Filipovich, A.H.; Henson, P.M.; Jordan, M.B. Hemophagocytosis causes a consumptive anemia of inflammation. J. Exp. Med. 2011, 208, 1203–1214. [Google Scholar] [CrossRef] [PubMed]
- Rekhtman, N.; Radparvar, F.; Evans, T.; Skoultchi, A.I. Direct interaction of hematopoietic transcription factors PU.1 and GATA-1: Functional antagonism in erythroid cells. Genes Dev. 1999, 13, 1398–1411. [Google Scholar] [CrossRef] [Green Version]
- Pronk, C.J.; Veiby, O.P.; Bryder, D.; Jacobsen, S.E.W. Tumor necrosis factor restricts hematopoietic stem cell activity in mice: Involvement of two distinct receptors. J. Exp. Med. 2011, 208, 1563–1570. [Google Scholar] [CrossRef] [Green Version]
- Roodman, G.D.; Bird, A.; Hutzler, D.; Montgomery, W. Tumor necrosis factor-alpha and hematopoietic progenitors: Effects of tumor necrosis factor on the growth of erythroid progenitors CFU-E and BFU-E and the hematopoietic cell lines K562, HL60, and HEL cells. Exp. Hematol. 1987, 15, 928–935. [Google Scholar]
- Fard, R.M.N.; Heidarpour, M.; Shojaei, A.; Zaeemi, M.; Javdani, G.; Golshahi, H. A case report of typical leishmaniasis in dog. J. Parasit. Dis. 2013, 39, 339–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torrecilha, R.; Utsunomiya, Y.T.; Bosco, A.; Almeida, B.; Pereira, P.; Narciso, L.; Pereira, D.; Baptistiolli, L.; Calvo-Bado, L.; Courtenay, O.; et al. Correlations between peripheral parasite load and common clinical and laboratory alterations in dogs with visceral leishmaniasis. Prev. Veter. Med. 2016, 132, 83–87. [Google Scholar] [CrossRef]
- Barrouin-Melo, S.M.; Larangeira, D.F.; Santos, S.O.; Chagas-Júnior, A.D.; Paixão, M.; Aguiar, P.H.; dos-Santos, W.L.; Pontes-de-Carvalho, L. A standardized cytological and immunochemical method for the analysis of fine-needle spleen aspi-rates: Assessment of leukocyte population changes in canine visceral leishmaniosis. Vet. Immunol. Immunopathol. 2006, 111, 251–261. [Google Scholar] [CrossRef] [Green Version]
- Andrade, T.M.; Carvalho, E.M.; Rocha, H. Bacterial Infections in Patients with Visceral Leishmaniasis. J. Infect. Dis. 1990, 162, 1354–1359. [Google Scholar] [CrossRef] [PubMed]
- Maia, C.; Campino, L. Cytokine and Phenotypic Cell Profiles of Leishmania infantum Infection in the Dog. J. Trop. Med. 2012, 2012, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Frequency | (%) |
|---|---|---|
| N | 63 | 100 |
| Male | 34/63 | 54.0 |
| Female | 29/63 | 46.0 |
| Size: | ||
| Small | 19/63 | 30.2 |
| Middle | 38/63 | 60.3 |
| Large | 06/63 | 9.5 |
| Estimated age: | ||
| Six months to 2 years old | 17/63 | 27.0 |
| 2–6 years old | 38/63 | 60.3 |
| More than 6 years old | 08/63 | 12.7 |
| Detection of L. infantum infection: | ||
| Infection confirmed by any test (total) | 50/63 | 79.3 |
| Spleen culture | 18/57 | 31.6 |
| ELISA | 47/58 | 81.0 |
| Clinical category: | ||
| Oligosymptomatic (2 or 3 clinical signs) | 24/63 | 38.1 |
| Polysymptomatic (more than 3 clinical signs) | 39/63 | 61.9 |
| Normal Parameters | ELISA (n = 47) | Spleen Culture (n = 18) |
|---|---|---|
| Clinical score | 3.7 a—SD: 1.4 | 5.1 b—SD: 1.4 |
| Blood tests | ||
| Erythrogram (5.5–8.5 × 106/μL) | 5.3 c—SD: 1.1 | 4.4 d—SD: 1.1 |
| Hemoglobin (12–18 g/dL) | 11.3 e—SD: 2.4 | 9.7 f—SD: 2.7 |
| Leukocytes (6–17 × 103/μL) | 13050 g—SD: 4637 | 19478 h—SD: 11863 |
| Platelets (200,000–500,000/mm) | 132521—SD: 113142 | 148588—SD: 120859 |
| Urea (21.4–59.92 mg/dL) | 40.2—SD: 16.6 | 43.8—SD: 16.3 |
| Creatinine (0.5–1.5 mg/dL) | 0.82—SD: 0.25 | 0.71—SD: 0.27 |
| Total proteins (5.4–7.1 g/dL) | 10.9—SD: 1.5 | 11.0—SD: 2.6 |
| Albumin (2.6–3.3 g/dL) | 3.0—SD: 0.8 | 2.7—SD: 0.9 |
| Globulin (2.7-4.4 g/dL) | 7.9—SD: 1.4 | 8.0—SD: 1.7 |
| A/G ratio | 0.4—SD: 0.1 | 0.3—SD: 0.1 |
| AST (21–45 UI/L) | 37.0 k—SD: 24.7 | 63.0 l—SD: 55.0 |
| ALT (21–73 UI/L) | 37.7—SD: 17.1 | 46.8—SD: 24.3 |
| TGL (20–112 mg/dL) | 51.5 m—SD: 25.8 | 70.6 n—SD: 22.5 |
| Bone marrow histology | (n = 26) | (n = 13) |
| Cellularity | 90.0%—SD: 11.7 | 94.2%—SD: 4.9 |
| Erythroid hypoplasia | 35%—N = 09/26 | 23%—N = 03/13 |
| Granulocytic hyperplasia | 96%—N = 25/26 | 100%—N = 13/13 |
| Megak. hyperplasia | 73%—N = 19/26 | 54%—N = 07/13 |
| Histiocytosis | 47% i—N = 12/26 | 92% j—N = 11/13 |
| Hemorrhage | 35%—N = 09/26 | 15%—N = 02/13 |
| Congestion | 27%—N = 07/26 | 31%—N = 05/13 |
| Emperipolesis | 35%—N = 09/26 | 23%—N = 03/13 |
| Megakaryocyte dysplasia | 69%—N = 18/26 | 39%—N = 05/13 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Almeida, V.; Lima, I.; Fraga, D.; Carrillo, E.; Moreno, J.; dos-Santos, W.L.C. Hematological Changes in Dogs with Visceral Leishmaniasis Are Associated with Increased IFN-γ and TNF Gene Expression Levels in the Bone Marrow. Microorganisms 2021, 9, 1618. https://doi.org/10.3390/microorganisms9081618
Almeida V, Lima I, Fraga D, Carrillo E, Moreno J, dos-Santos WLC. Hematological Changes in Dogs with Visceral Leishmaniasis Are Associated with Increased IFN-γ and TNF Gene Expression Levels in the Bone Marrow. Microorganisms. 2021; 9(8):1618. https://doi.org/10.3390/microorganisms9081618
Chicago/Turabian StyleAlmeida, Valter, Isadora Lima, Deborah Fraga, Eugenia Carrillo, Javier Moreno, and Washington L. C. dos-Santos. 2021. "Hematological Changes in Dogs with Visceral Leishmaniasis Are Associated with Increased IFN-γ and TNF Gene Expression Levels in the Bone Marrow" Microorganisms 9, no. 8: 1618. https://doi.org/10.3390/microorganisms9081618