
Human and Animal Dirofilariasis in Southeast of France

 , , , and
, , , and 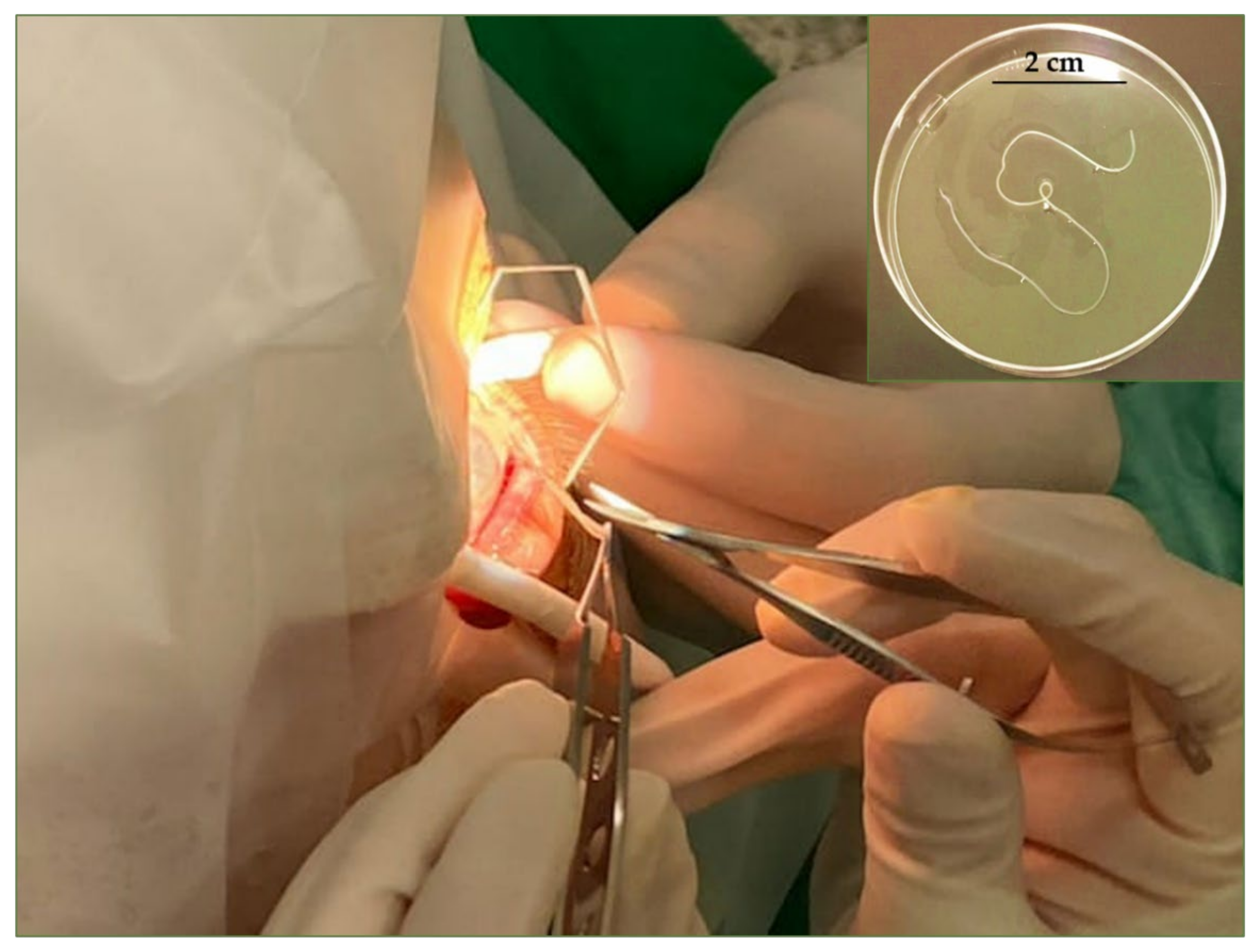{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Sampling and Study Area
2.2. Case Report
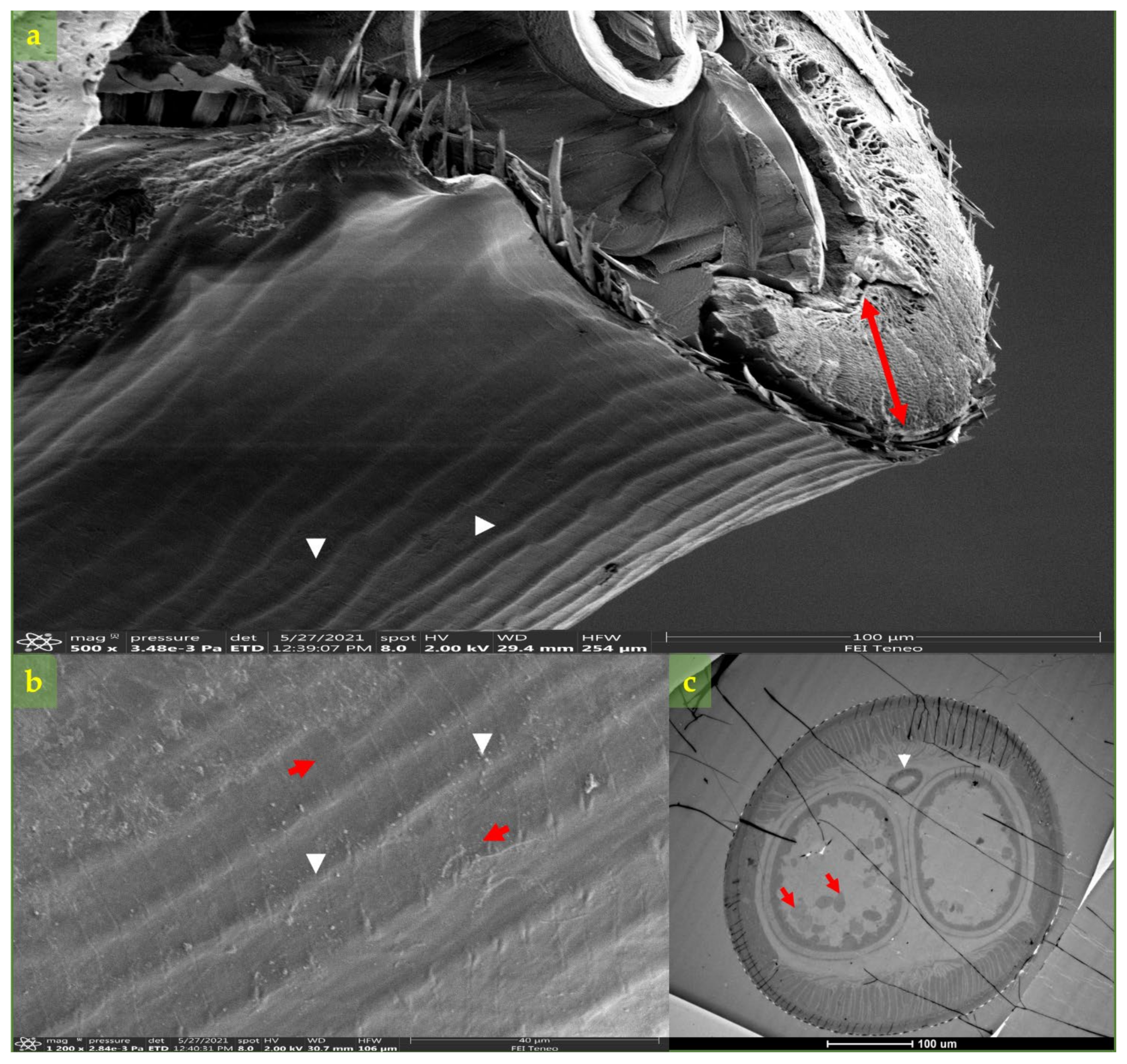
2.3. Morphological and Molecular Diagnosis of Dirofilaria spp.
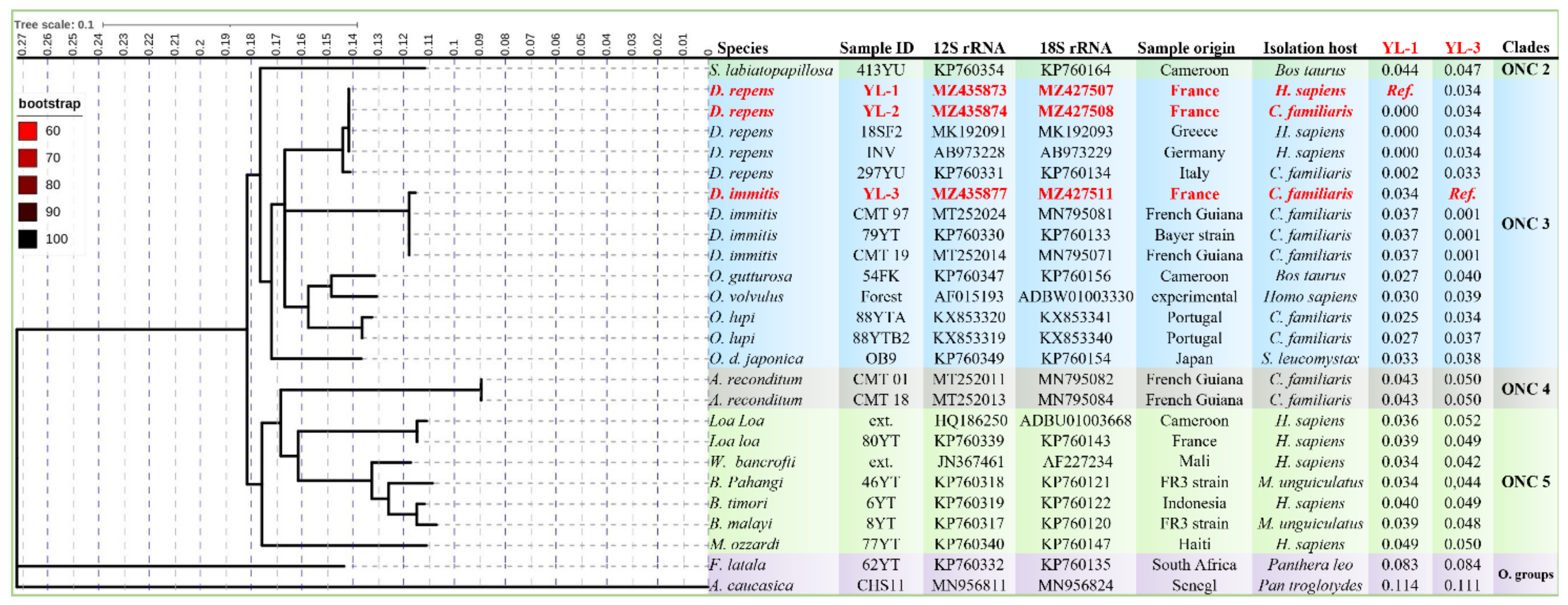
2.4. Phylogenetic Analysis
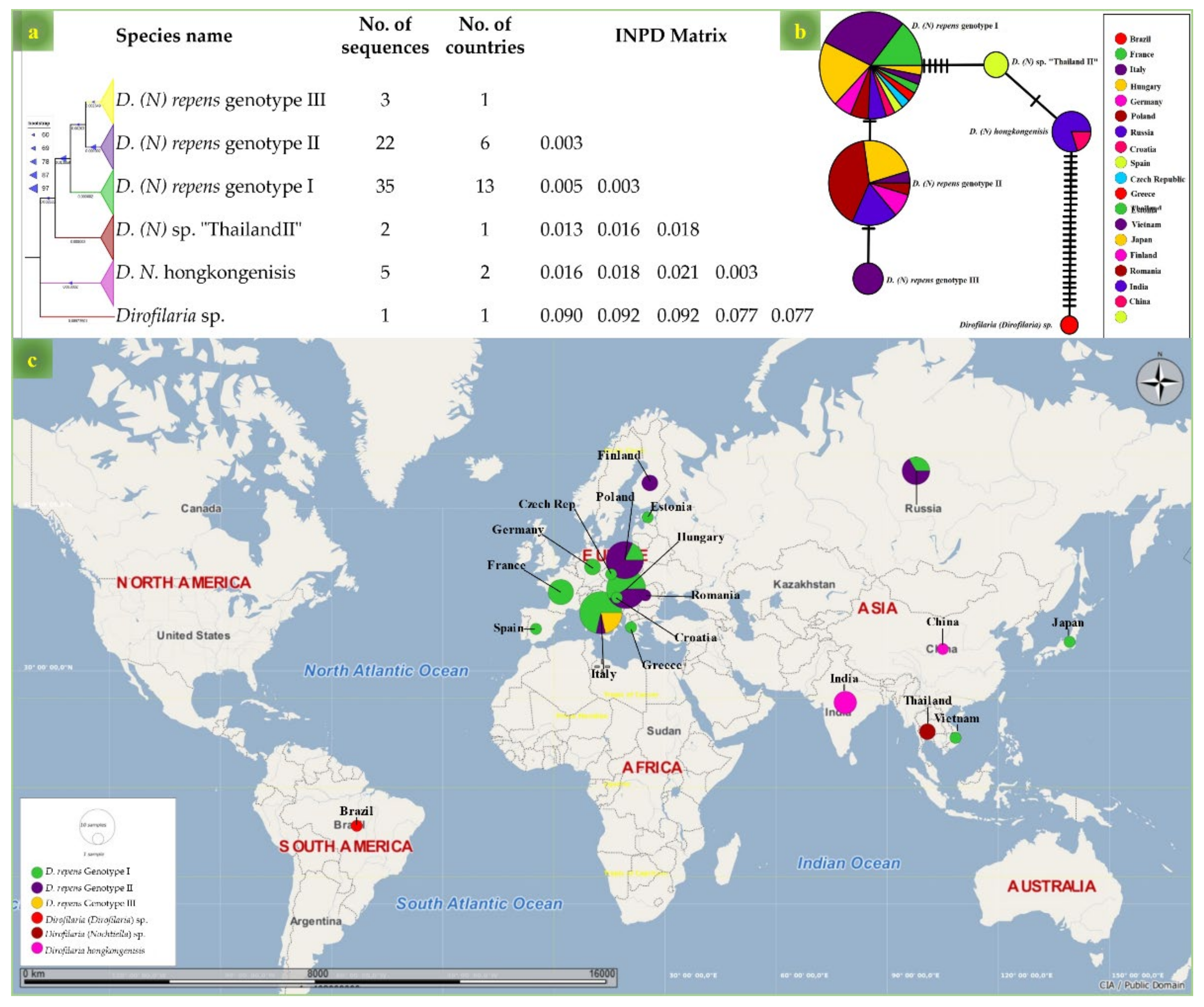
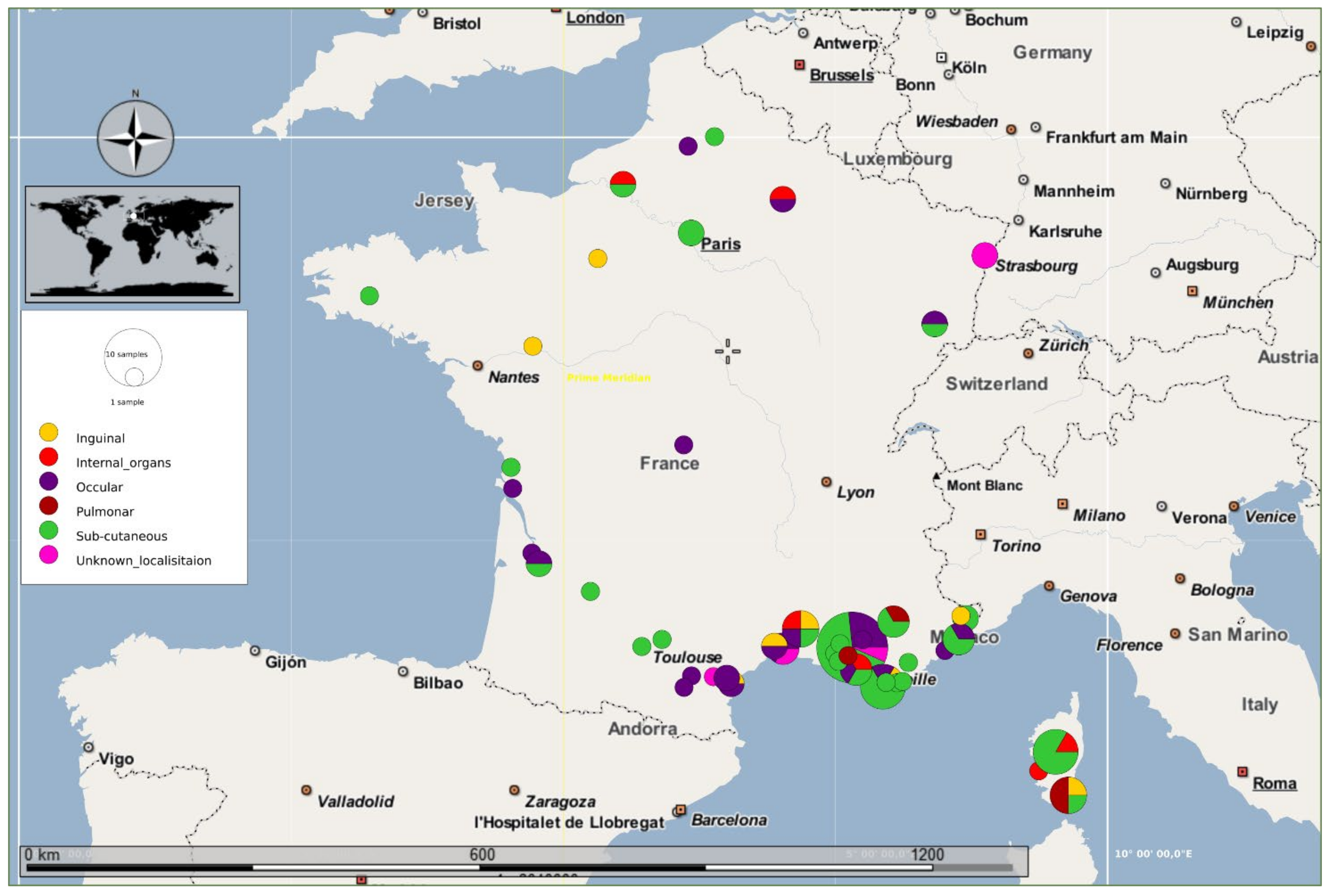
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Addario, C. Su di un nematode dell’occhio umano. Ann. Ottalmolog. 1885, 14, 134. [Google Scholar]
- Pampiglione, S. Dirofilariosi umana sottocongiuntivale: Su di un probabile caso osservato in Francia da Amato Lusitano nel XVI secolo. Parassitologia 1995, 37, 75. [Google Scholar] [PubMed]
- Aldrovandi, U.; Ferroni, C.; Ferroni, G.B.; Real Colegio de Cirugía de San Carlos (Madrid). De Animalibus Insectis Libri Septem cum Singulorum Iconibus ad Viuum Expressis; Denuo impress. Bonon.: Apud Clementem Ferronium: Lausanne, Switzerland, 1698; pp. 1–767. [Google Scholar]
- Simón, F.; Siles-Lucas, M.; Morchón, R.; González-Miguel, J.; Mellado, I.; Carretón, E.; Montoya-Alonso, J.A. Human and animal dirofilariasis: The emergence of a zoonotic mosaic. Clin. Microbiol. Rev. 2012, 25, 507–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otranto, D.; Dantas-Torres, F.; Brianti, E.; Traversa, D.; Petrić, D.; Genchi, C.; Capelli, G. Vector-borne helminths of dogs and humans in Europe. Parasit. Vectors 2013, 6, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dantas-Torres, F.; Otranto, D. Dirofilariosis in the Americas: A more virulent Dirofilaria immitis? Parasit. Vectors 2013, 6, 1. [Google Scholar] [CrossRef] [Green Version]
- Orihel, T.C.; Eberhard, M.L. Zoonotic filariasis. Clin. Microbiol. Rev. 1998, 11, 366–381. [Google Scholar] [CrossRef] [Green Version]
- Canestri Trotti, G.; Pampiglione, S.; Rivasi, F. The species of the genus Dirofilaria, Railliet & Henry, 1911. Parassitologia 1997, 39, 369–374. [Google Scholar]
- Simón, F.; González-Miguel, J.; Diosdado, A.; Gómez, P.J.; Morchón, R.; Kartashev, V. The complexity of zoonotic filariasis episystem and its consequences: A multidisciplinary view. Biomed. Res. Int. 2017, 2017, 6436130. [Google Scholar] [CrossRef] [Green Version]
- Aonuma, H.; Yoshimura, A.; Perera, N.; Shinzawa, N.; Bando, H.; Oshiro, S.; Nelson, B.; Fukumoto, S.; Kanuka, H. Loop-mediated isothermal amplification applied to filarial parasites detection in the mosquito vectors: Dirofilaria immitis as a study model. Parasit. Vectors 2009, 2, 15. [Google Scholar] [CrossRef] [Green Version]
- Ducos de Lahitte, J.; Davoust, B.; Dorchies, P. Filariose canine à Dirofilaria immitis. Enquête sur la fréquence et la répartition en zone méditerranéenne. Bull. Soc. Fr. Parasitol. 1984, 3, 105–108. [Google Scholar]
- Davoust, B.; Ducos de Lahitte, J. Development of dirofilariosis enzooty in the military kennels of south-east France. Rev. Med. Vet. 1989, 140, 15–19. (In French) [Google Scholar] [CrossRef]
- Doby, J.M.; Guiguen, C.; Lefeuvre, R. Présence de Dirofilaria immitis en Bretagne. Bull. Soc. Fr. Parasitol. 1986, 4, 51–54. [Google Scholar]
- Doby, J.M.; Couatarmanach, A.; Aznar, C. Filarioses canines par Dirofilaria immitis (Leidy, 1856) et D. repens (Railliet et Henry, 1911), dans l’Ouest de la France. Bull. Soc. Fr. Parasitol. 1986, 4, 229–233. [Google Scholar]
- Laidoudi, Y.; Ringot, D.; Watier-Grillot, S.; Davoust, B.; Mediannikov, O. A cardiac and subcutaneous canine dirofilariosis outbreak in a kennel in central France. Parasite 2019, 26, 72. [Google Scholar] [CrossRef]
- Tahir, D.; Petit, C. Filariose sous-cutanée à Dirofilaria repens. L’Essentiel 2017, 435, 20–24. [Google Scholar]
- Tahir, D.; Bittar, F.; Barré-Cardi, H.; Sow, D.; Dahmani, M.; Mediannikov, O.; Raoult, D.; Davoust, B.; Parola, P. Molecular survey of Dirofilaria immitis and Dirofilaria repens by new real-time TaqMan®PCR assay in dogs and mosquitoes (Diptera: Culicidae) in Corsica (France). Vet. Parasitol. 2017, 235, 1–7. [Google Scholar] [CrossRef]
- Benzaquen, M.; Marmottant, E.; Parola, P.; Berbis, P. Update on cutaneous dirofilariasis. Ann. Dermatol. Venereol. 2017, 144, 607–611. [Google Scholar] [CrossRef]
- Fournier, G.; Morquin, D.; Goulabchand, R.; Tingaud, C.; de Boutray, M. Dirofilariose autochtone intra-musculaire temporale. Med. Mal. Infect. 2018, 48, 424–426. [Google Scholar] [CrossRef]
- Raccurt, C.P. La dirofilariose, zoonose emergente et meconnue en France. Med. Trop. 1999, 59, 389–400. [Google Scholar]
- Marty, P.; Le Fichoux, Y.; Dellamonica, P.; Rosa, E.; Kempf, B. Filariose sous-cutanée à Dirofilaria repens. Une nouvelle observation française. Nouv. Presse Med. 1981, 10, 2114–2115. [Google Scholar]
- Pampiglione, S.; Peraldi, R.; Burelli, J.P. Dirofilariose humaine en Corse: Un nouveau cas autochtone. Révision des cas déja publiés. Bull. Soc. Pathol. Exot. 1999, 92, 305–308. [Google Scholar]
- Basset, D.; Lachaud, L.; Marmouset-Ottaviani, C.; Dedet, J.-P. Nouveau cas de dirofilariose en Corse. Presse Med. 2003, 32, 702. [Google Scholar] [PubMed]
- Basset, D.; Villain, M.; Bastien, P.; Taupin, P.; Jarry, D.M. La dirofilariose en France: Un cas languedocien chez un sujet sérologiquement positif pour le VIH. Presse Med. 1996, 25, 1372–1375. [Google Scholar]
- Foissac, M.; Million, M.; Mary, C.; Dales, J.P.; Souraud, J.B.; Piarroux, R.; Parola, P. Subcutaneous infection with Dirofilaria immitis nematode in human, France. Emerg. Infect. Dis. 2013, 19, 171–173. [Google Scholar] [CrossRef] [PubMed]
- Benzaquen, M.; Brajon, D.; Delord, M.; Yin, N.; Bittar, F.; Toga, I.; Berbis, P.; Parola, P. Cutaneous and pulmonary dirofilariasis due to Dirofilaria repens. Br. J. Dermatol. 2015, 173, 788–791. [Google Scholar] [CrossRef]
- Yilmaz, E.; Fritzenwanker, M.; Pantchev, N.; Lendner, M.; Wongkamchai, S.; Otranto, D.; Kroidl, I.; Dennebaum, M.; Le, T.H.; Anh Le, T.; et al. The mitochondrial genomes of the zoonotic canine filarial parasites Dirofilaria (Nochtiella) repens and Candidatus Dirofilaria (Nochtiella) honkongensis provide evidence for presence of cryptic species. PLoS Negl. Trop. Dis. 2016, 10, e0005028. [Google Scholar] [CrossRef] [PubMed]
- Dantas-Torres, F.; Otranto, D. On the validity of “Candidatus Dirofilaria hongkongensis” and on the use of the provisional status Candidatus in zoological nomenclature. Parasit. Vectors 2020, 13, 287. [Google Scholar] [CrossRef]
- Laidoudi, Y.; Davoust, B.; Varloud, M.; Niang, E.H.A.; Fenollar, F.; Mediannikov, O. Development of a multiplex qPCR-based approach for the diagnosis of Dirofilaria immitis, D. repens and Acanthocheilonema reconditum. Parasit. Vectors 2020, 13, 319. [Google Scholar] [CrossRef] [PubMed]
- Laidoudi, Y.; Medkour, H.; Latrofa, M.S.; Davoust, B.; Diatta, G.; Sokhna, C.; Barciela, A.; Hernandez-Aguilar, R.A.; Raoult, D.; Otranto, D.; et al. Zoonotic Abbreviata caucasica in wild chimpanzees (Pan troglodytes verus) from Senegal. Pathogens 2020, 9, 517. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Hall, T.; Biosciences, I.; Carlsbad, C. BioEdit: An important software for molecular biology. GERF Bull. Biosci. 2011, 2, 60–61. [Google Scholar] [CrossRef]
- Tamura, K. Estimation of the number of nucleotide substitutions when there are strong transition-transversion and G+C-content biases. Mol. Biol. Evol. 1992, 9, 678–687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Lefoulon, E.; Bain, O.; Bourret, J.; Junker, K.; Guerrero, R.; Cañizales, I.; Kuzmin, Y.; Satoto, T.B.T.; Cardenas-Callirgos, J.M.; de Souza Lima, S.; et al. Shaking the Tree: Multi-locus sequence typing usurps current onchocercid (Filarial Nematode) phylogeny. PLoS Negl. Trop. Dis. 2015, 9, 1–19. [Google Scholar] [CrossRef]
- Katoh, K.; Misawa, K.; Kuma, K.I.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [Green Version]
- Gouy, M.; Guindon, S.; Gascuel, O. Sea view version 4: A multiplatform graphical user interface for sequence alignment and phylogenetic tree building. Mol. Biol. Evol. 2010, 27, 221–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, L.T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Minh, B.Q.; Nguyen, M.A.T.; Von Haeseler, A. Ultrafast approximation for phylogenetic bootstrap. Mol. Biol. Evol. 2013, 30, 1188–1195. [Google Scholar] [CrossRef] [PubMed]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [Green Version]
- Afgan, E.; Baker, D.; Batut, B.; Beek, M.; Bouvier, D.; Cech, M.; Chilton, J.; Clements, D.; Coraor, N.; Gruning, B.A.; et al. The Galaxy platform for accessible, reproducible and collaborative biomedical analyses: 2018 update. Nucleic Acids Res. 2018, 46, 537–544. [Google Scholar] [CrossRef] [Green Version]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, 256–259. [Google Scholar] [CrossRef] [Green Version]
- Otranto, D.; Diniz, D.G.; Dantas-Torres, F.; Casiraghi, M.; de Almeida, I.N.F.; de Almeida, L.N.F.; Dos Santos, J.N.; Furtado, A.P.; de Almeida Sobrinho, E.F.; Bain, O. Human intraocular filariasis caused by Dirofilaria sp. nematode, Brazil. Emerg. Infect. Dis. 2011, 17, 863–866. [Google Scholar] [CrossRef]
- Leigh, J.W.; Bryant, D. PopART: Full-feature software for haplotype network construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- Wong, M.M.; Brummer, M.E.G. Cuticular morphology of five species of Dirofilaria: A scanning electron microscope study. J. Parasitol. 1978, 64, 108–114. [Google Scholar] [CrossRef] [PubMed]
- Ministère des Solidarités et de la Santé (France). Cartes de Présence du Moustique Tigre (Aedes albopictus) en France Métropolitaine. Available online: https://solidarites-sante.gouv.fr/sante-et-environnement/risques-microbiologiques-physiques-et-chimiques/especes-nuisibles-et-parasites/article/cartes-de-presence-du-moustique-tigre-aedes-albopictus-en-france-metropolitaine (accessed on 18 January 2021).
- Medkour, H.; Laidoudi, Y.; Marié, J.-L.; Fenollar, F.; Davoust, B.; Mediannikov, O. Molecular investigation of vector-borne pathogens in red foxes (Vulpes vulpes) from southern france. J. Wildl. Dis. 2020, 56, 837–850. [Google Scholar] [CrossRef] [PubMed]
- Dantas-Torres, F.; Otranto, D. Best practices for preventing vector-borne diseases in dogs and humans. Trends Parasitol. 2016, 32, 43–55. [Google Scholar] [CrossRef]
- Schorderet-Weber, S.; Noack, S.; Selzer, P.M.; Kaminsky, R. Blocking transmission of vector-borne diseases. Int. J. Parasitol. Drugs Drug Resist. 2017, 7, 90–109. [Google Scholar] [CrossRef]
- Schnieder, T. Bekämpfung von Ektoparasiten (Flöhe, Zecken, Läuse, Haarlinge, Sand- und Stechmücken) bei Hunden und Katzen. European Scientific Counsel Companion Animal Parasites (ESCCAP). J. Verbrauch. Lebensm. 2010, 5, 351–369. [Google Scholar] [CrossRef]
- Nelson, C.T.; McCall, J.W.; Jones, S.; Moorhead, A. Current canine guidelines for the prevention, diagnosis, and management of heartworm (Dirofilaria immitis) infection in dogs. Am. Hear. Soc. 2018, 2018, 1–35. [Google Scholar]
- Laidoudi, Y.; Medkour, H.; Tahir, D.; Dahmana, H.; Marié, J.-L.; Varloud, M.; Mediannikov, O.; Davoust, B. Field implementation of a monthly multi-modal prophylactic strategy (Dinotefuran-Permethrin-Pyriproxyfen and Milbemycine Oxime-Praziquantel) against canine vector-borne diseases in Corsica. Preprints 2020. [Google Scholar] [CrossRef]
- McCall, J.W.; Genchi, C.; Kramer, L.H.; Guerrero, J.; Venco, L. Heartworm disease in animals and humans. Adv. Parasitol. 2008, 66, 193–285. [Google Scholar] [CrossRef] [PubMed]
- Wysmołek, M.E.; Klockiewicz, M.; Sobczak-filipiak, M. Case studies of severe microfilaremia in four dogs naturally infected with case studies of severe microfilaremia in four dogs naturally infected with Dirofilaria repens as the primary disease or a disease complicating factor. Front. Vet. Sci. 2020, 7, 577466. [Google Scholar] [CrossRef] [PubMed]
- Basset, D.; Sauguet, P. Dirofilaria et dirofilarioses. Rev. Francoph. Lab. 2016, 483, 37–44. [Google Scholar] [CrossRef]
- Otranto, D.; Dantas-Torres, F.; Breitschwerdt, E.B. Managing canine vector-borne diseases of zoonotic concern: Part two. Trends Parasitol. 2009, 25, 228–235. [Google Scholar] [CrossRef]
- Panarese, R.; Iatta, R.; Mendoza-Roldan, J.A.; Szlosek, D.; Braff, J.; Liu, J.; Beugnet, F.; Dantas-Torres, F.; Beall, M.J.; Otranto, D. Comparison of diagnostic tools for the detection of Dirofilaria immitis infection in dogs. Pathogens 2020, 9, 499. [Google Scholar] [CrossRef]
- Yilmaz, E.; Wongkamchai, S.; Ramünke, S.; Koutsovoulos, G.D.; Blaxter, M.L.; Poppert, S.; Schaper, R.; von Samson-Himmelstjerna, G.; Krücken, J. High genetic diversity in the Dirofilaria repens species complex revealed by mitochondrial genomes of feline microfilaria samples from Narathiwat, Thailand. Transbound. Emerg. Dis. 2019, 66, 389–399. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, J.; Kobayashi, S.; Okata, U.; Matsuzaki, H.; Mori, M.; Chen, K.R.; Iwata, S. Molecular analysis of Dirofilaria repens removed from a subcutaneous nodule in a Japanese woman after a tour to Europe. Parasite 2015, 22, 2. [Google Scholar] [CrossRef] [Green Version]
- Ciuca, L.; Simòn, F.; Rinaldi, L.; Kramer, L.; Genchi, M.; Cringoli, G.; Acatrinei, D.; Miron, L.; Morchon, R. Seroepidemiological survey of human exposure to Dirofilaria spp. in Romania and Moldova. Acta Trop. 2018, 187, 169–174. [Google Scholar] [CrossRef] [PubMed]
- Otranto, D.; Brianti, E.; Gaglio, G.; Dantas-Torres, F.; Azzaro, S.; Giannetto, S. Human ocular infection with Dirofilaria repens (Railliet and Henry, 1911) in an area endemic for canine dirofilariasis. Am. J. Trop. Med. Hyg. 2011, 84, 1002–1004. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Laidoudi, Y.; Otranto, D.; Stolowy, N.; Amrane, S.; Santhakumari Manoj, R.R.; Polette, L.; Watier-Grillot, S.; Mediannikov, O.; Davoust, B.; L'Ollivier, C. Human and Animal Dirofilariasis in Southeast of France. Microorganisms 2021, 9, 1544. https://doi.org/10.3390/microorganisms9071544
Laidoudi Y, Otranto D, Stolowy N, Amrane S, Santhakumari Manoj RR, Polette L, Watier-Grillot S, Mediannikov O, Davoust B, L'Ollivier C. Human and Animal Dirofilariasis in Southeast of France. Microorganisms. 2021; 9(7):1544. https://doi.org/10.3390/microorganisms9071544
Chicago/Turabian StyleLaidoudi, Younes, Domenico Otranto, Natacha Stolowy, Sophie Amrane, Ranju Ravindran Santhakumari Manoj, Laurine Polette, Stéphanie Watier-Grillot, Oleg Mediannikov, Bernard Davoust, and Coralie L'Ollivier. 2021. "Human and Animal Dirofilariasis in Southeast of France" Microorganisms 9, no. 7: 1544. https://doi.org/10.3390/microorganisms9071544