
Fungicide Resistance Evolving in Ramularia collo-cygni Population in Estonia


Abstract
:
1. Introduction
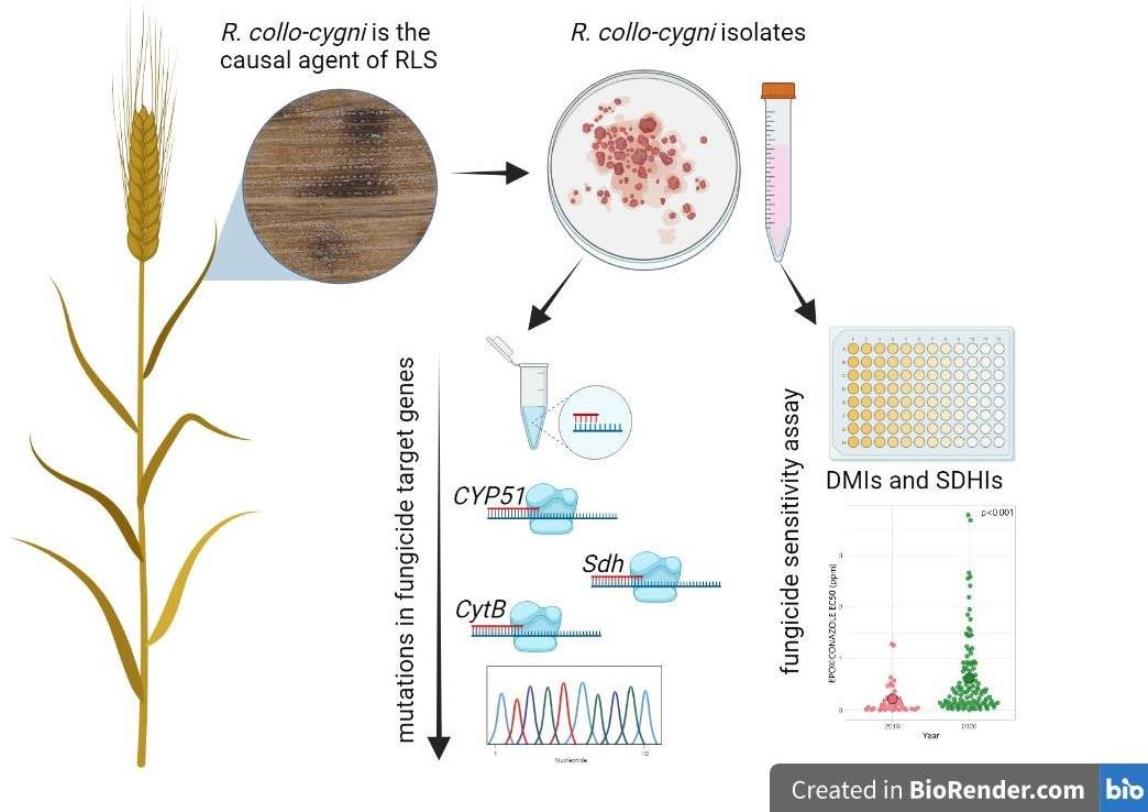
2. Materials and Methods
2.1. Isolate Collection
2.2. Determination of Fungicide Sensitivity
2.3. Identifying Target Site Mutations in the Genes of Fungicide Target Proteins
2.4. Statistical Analysis
3. Results
3.1. Status of DMI Fungicide Sensitivity
3.2. Mutations in CYP51 Gene
3.3. Status of SDHI Fungicide Sensitivity
3.4. Mutations in Sdh Protein Subunits
3.5. Mutation G143A Prevalence in CytB Gene
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Newton, A.C.; Flavell, A.J.; George, T.S.; Leat, P.; Mullholland, B.; Ramsay, L.; Revoredo-Giha, C.; Russell, J.; Steffenson, B.J.; Stuart Swanston, J.; et al. Crops that feed the world 4. Barley: A resilient crop? Strengths and weaknesses in the context of food security. Food Secur. 2011, 3, 141–178. [Google Scholar] [CrossRef]
- FAOSTAT. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 17 February 2021).
- Walters, D.R.; Havis, N.D.; Oxley, S.J.P. Ramularia collo-cygni: The biology of an emerging pathogen of barley. FEMS Microbiol. Lett. 2008, 279, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Havis, N.D.; Brown, J.K.M.; Clemente, G.; Frei, P.; Jedryczka, M.; Kaczmarek, J.; Kaczmarek, M.; Matusinsky, P.; McGrann, G.R.D.; Pereyra, S.; et al. Ramularia collo-cygni—An emerging pathogen of barley crops. Phytopathology 2015, 105, 895–904. [Google Scholar] [CrossRef] [Green Version]
- McGrann, G.R.; Havis, N.D. Ramularia leaf spot: A newly important threat to barley production. Outlooks Pest Manag. 2017, 28, 65–69. [Google Scholar] [CrossRef]
- Sooväli, P.; Tikhonova, M.; Matušinsky, P. First report of Ramularia leaf spot caused by Ramularia collo-cygni on leaves and seeds of barley in Estonia. Plant Dis. 2014, 98, 997. [Google Scholar] [CrossRef]
- McGrann, G.R.D.; Andongabo, A.; Sjõkvist, E.; Trivedi, U.; Dussart, F.; Kaczmarek, M.; Mackenzie, A.; Fountaine, J.M.; Taylor, J.M.G.; Paterson, L.J.; et al. The genome of the emerging barley pathogen Ramularia collo-cygni. BMC Genom. 2016, 17, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stabentheiner, E.; Minihofer, T.; Huss, H. Infection of barley by Ramularia collo-cygni: Scanning electron microscopic investigations. Mycopathologia 2009, 168, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Havis, N.D.; Nyman, M.; Oxley, S.J.P. Evidence for seed transmission and symptomless growth of Ramularia collo-cygni in barley (Hordeum vulgare). Plant Pathol. 2014, 63, 929–936. [Google Scholar] [CrossRef]
- Rodriguez, R.; Redman, R. More than 400 million years of evolution and some plants still can’t make it on their own: Plant stress tolerance via fungal symbiosis. J. Exp. Bot. 2008, 59, 1109–1114. [Google Scholar] [CrossRef] [PubMed]
- Schützendübel, A.; Stadler, M.; Wallner, D.; Von Tiedemann, A. A hypothesis on physiological alterations during plant ontogenesis governing susceptibility of winter barley to ramularia leaf spot. Plant Pathol. 2008, 57, 518–526. [Google Scholar] [CrossRef]
- Kaczmarek, M.; Piotrowska, M.J.; Fountaine, J.M.; Gorniak, K.; McGrann, G.R.D.; Armstrong, A.; Wright, K.M.; Newton, A.C.; Havis, N.D. Infection strategy of Ramularia collo-cygni and development of ramularia leaf spot on barley and alternative graminaceous hosts. Plant Pathol. 2017, 66, 45–55. [Google Scholar] [CrossRef]
- Harvey, I.C. Epidemiology and control of leaf and awn spot of barley caused by Ramularia collo-cygni. N. Z. Plant Prot. 2002, 55, 331–335. [Google Scholar] [CrossRef] [Green Version]
- Sjokvist, E.; Lemcke, R.; Kamble, M.; Turner, F.; Blaxter, M.; Havis, N.H.D.; Lyngkjær, M.F.; Radutoiu, S. Dissection of ramularia leaf spot disease by integrated analysis of barley and Ramularia collo-cygni transcriptome responses. Mol. Plant Microbe Interact. 2019, 32, 176–193. [Google Scholar] [CrossRef] [Green Version]
- Leisova-Svobodova, L.; Matusinsky, P.; Kucera, L. Variability of the Ramularia collo-cygni Population in Central Europe. J. Phytopathol. 2012, 160, 701–709. [Google Scholar] [CrossRef]
- Piotrowska, M.J.; Ennos, R.A.; Fountaine, J.M.; Burnett, F.J.; Kaczmarek, M.; Hoebe, P.N. Development and use of microsatellite markers to study diversity, reproduction and population genetic structure of the cereal pathogen Ramularia collo-cygni. Fungal Genet. Biol. 2016, 87, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Ficsor, A.; Tóth, B.; Varga, J.; Csősz, M.; Tomcsányi, A.; Mészáros, K.; Kótai, E.; Bakonyi, J. Variability of Pyrenophora teres f. teres in Hungary as revealed by mating type and RAPD analyses. J. Plant Pathol. 2014, 96, 515–523. [Google Scholar] [CrossRef]
- Poudel, B.; Vaghefi, N.; McLean, M.S.; Platz, G.J.; Sutherland, M.W.; Martin, A. Genetic structure of a Pyrenophora teres f. teres population over time in an Australian barley field as revealed by Diversity Arrays Technology markers. Plant Pathol. 2019, 68, 1331–1336. [Google Scholar] [CrossRef]
- McGrann, G.R.D.; Steed, A.; Burt, C.; Goddard, R.; Lachaux, C.; Bansal, A.; Corbitt, M.; Gorniak, K.; Nicholson, P.; Brown, J.K.M. Contribution of the drought tolerance-related Stress-responsive NAC1 transcription factor to resistance of barley to Ramularia leaf spot. Mol. Plant Pathol. 2015, 16, 201–209. [Google Scholar] [CrossRef] [Green Version]
- Wolter, M.; Hollricher, K.; Salamini, F.; Schulze-Lefert, P. The mlo resistance alleles to powdery mildew infection in barley trigger a developmentally controlled defence mimic phenotype. MGG Mol. Gen. Genet. 1993, 239, 122–128. [Google Scholar] [CrossRef]
- Piffanelli, P.; Zhou, F.; Casais, C.; Orme, J.; Jarosch, B.; Schaffrath, U.; Collins, N.C.; Panstruga, R.; Schulze-Lefert, P. The barley MLO modulator of defense and cell death is responsive to biotic and abiotic stress stimuli. Plant Physiol. 2002, 129, 1076–1085. [Google Scholar] [CrossRef] [Green Version]
- McGrann, G.R.D.; Stavrinides, A.; Russell, J.; Corbitt, M.M.; Booth, A.; Chartrain, L.; Thomas, W.T.B.; Brown, J.K.M. A trade off between mlo resistance to powdery mildew and increased susceptibility of barley to a newly important disease, Ramularia leaf spot. J. Exp. Bot. 2014, 65, 1025–1037. [Google Scholar] [CrossRef] [Green Version]
- West, J.S.; Townsend, J.A.; Stevens, M.; Fitt, B.D.L. Comparative biology of different plant pathogens to estimate effects of climate change on crop diseases in Europe. Eur. J. Plant Pathol. 2012, 133, 315–331. [Google Scholar] [CrossRef] [Green Version]
- Peraldi, A.; Griffe, L.L.; Burt, C.; Mcgrann, G.R.D.; Nicholson, P. Brachypodium distachyon exhibits compatible interactions with Oculimacula spp. and Ramularia collo-cygni, providing the first pathosystem model to study eyespot and ramularia leaf spot diseases. Plant Pathol. 2014, 63, 554–562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sooväli, P.; Koppel, M. Efficacy of fungicide tebuconazole in barley varieties with different resistance level. Agric. Food Sci. 2010, 19, 34–42. [Google Scholar] [CrossRef]
- Mäe, A.; Põllumaa, L.; Sooväli, P. Ramularia collo-cygni: A new pathogen spreading in barley fields in Estonia. Agric. Food Sci. 2018, 27, 138–145. [Google Scholar] [CrossRef]
- Stam, R.; Sghyer, H.; Tellier, A.; Hess, M.; Hückelhoven, R. The current epidemic of the barley pathogen Ramularia collo-cygni derives from a population expansion and shows global admixture. Phytopathology 2019, 109, 2161–2168. [Google Scholar] [CrossRef]
- Kuck, K.-H.; Russell, P.E. FRAC: Combined resistance risk assessment. Asp. Appl. Biol. 2006, 78, 3–10. [Google Scholar]
- Fountaine, J.M.; Fraaije, B.A. Development of QoI resistant alleles in populations of Ramularia collo-cygni. In Aspects of Applied Biology 92, Proceedings of the 2nd European Ramularia Workshop—A New Disease and Challenge in Barley Production, Edinburgh, UK, 7–8 April 2009; Association of Applied Biologists: Warwick, UK; The Warwick Enterprise Park Wellesbourne: Edinburgh, UK, 2009; pp. 123–126. [Google Scholar]
- Matusinsky, P.; Svobodova-Leisova, L.; Marik, P.; Tvaruzek, L.; Stemberkova, L.; Hanusova, M.; Minarikova, V.; Vysohlidova, M.; Spitzer, T. Frequency of a mutant allele of cytochrome b conferring resistance to QoI fungicides in the Czech population of Ramularia collo-cygni. J. Plant Dis. Prot. 2010, 117, 1861–3829. [Google Scholar] [CrossRef]
- Havis, N.D.; Gorniak, K.; Taylor, J.; Stanisz-Migal, M.; Burnett, F.J. Controlling Ramularia leaf spot in barley crops. In Proceedings of the Crop Production in Northern Britain 2018, Dundee, UK, 27–28 February 2018; The Association for Crop Protection in Northern Britain: Dundee, UK, 2018; pp. 91–96. [Google Scholar]
- Arena, M.; Auteri, D.; Barmaz, S.; Bellisai, G.; Brancato, A.; Brocca, D.; Bura, L.; Byers, H.; Chiusolo, A.; Court Marques, D.; et al. Peer review of the pesticide risk assessment of the active substance chlorothalonil. EFSA J. 2018, 16, 5126–5165. [Google Scholar] [CrossRef]
- Rehfus, A.; Matusinsky, P.; Strobel, D.; Bryson, R.; Stammler, G. Mutations in target genes of succinate dehydrogenase inhibitors and demethylation inhibitors in Ramularia collo-cygni in Europe. J. Plant Dis. Prot. 2019, 126, 447–459. [Google Scholar] [CrossRef]
- Piotrowska, M.J.; Fountaine, J.M.; Ennos, R.A.; Kaczmarek, M.; Burnett, F.J. Characterisation of Ramularia collo-cygni laboratory mutants resistant to succinate dehydrogenase inhibitors. Pest Manag. Sci. 2017, 73, 1187–1196. [Google Scholar] [CrossRef] [Green Version]
- FRAC—Fungicide Resistance Action Committee. Available online: https://www.frac.info/ (accessed on 10 March 2021).
- Rehfus, A.; Miessner, S.; Achenbach, J.; Strobel, D.; Bryson, R.; Stammler, G. Emergence of succinate dehydrogenase inhibitor resistance of Pyrenophora teres in Europe. Pest Manag. Sci. 2016, 72, 1977–1988. [Google Scholar] [CrossRef]
- Rehfus, A.; Strobel, D.; Bryson, R.; Stammler, G. Mutations in sdh genes in field isolates of Zymoseptoria tritici and impact on the sensitivity to various succinate dehydrogenase inhibitors. Plant Pathol. 2018, 67, 175–180. [Google Scholar] [CrossRef]
- Dooley, H.; Shaw, M.W.; Mehenni-Ciz, J.; Spink, J.; Kildea, S. Detection of Zymoseptoria tritici SDHI-insensitive field isolates carrying the SdhC-H152R and SdhD-R47W substitutions. Pest Manag. Sci. 2016, 72, 2203–2207. [Google Scholar] [CrossRef]
- Clark, W.S. Septoria tritici and azole performance. Asp. Appl. Biol. 2006, 78, 127–132. [Google Scholar]
- Lucas, J.A.; Hawkins, N.J.; Fraaije, B.A. The Evolution of Fungicide Resistance. In Advances in Applied Microbiology; Elsevier Ltd.: Amsterdam, The Netherlands, 2015; Volume 90, pp. 29–92. [Google Scholar]
- Huf, A.; Rehfus, A.; Lorenz, K.H.; Bryson, R.; Voegele, R.T.; Stammler, G. Proposal for a new nomenclature for CYP51 haplotypes in Zymoseptoria tritici and analysis of their distribution in Europe. Plant Pathol. 2018, 67, 1706–1712. [Google Scholar] [CrossRef]
- Mair, W.; Lopez-Ruiz, F.; Stammler, G.; Clark, W.; Burnett, F.; Hollomon, D.; Ishii, H.; Thind, T.S.; Brown, J.K.; Fraaije, B.; et al. Proposal for a unified nomenclature for target-site mutations associated with resistance to fungicides. Pest Manag. Sci. 2016, 72, 1449–1459. [Google Scholar] [CrossRef] [PubMed]
- Leistrumaitė, A.; Liatukas, Ž. Resistance of spring barley cultivars to the new disease Ramularia leaf spot, caused by Ramularia collo-cygni. Agron. Res. 2006, 4, 251–255. [Google Scholar]
- Strobel, D.; Bryson, R.; Stammler, G.; Prochnow, J. A European overview of the occurrence of Ramularia collo-cygni and its sensitivity to fluxapyroxad. In Proceedings of the 11th Conference of the European Foundation for Plant Pathology—Healthy People, Krakow, Poland, 8–13 September 2014. [Google Scholar]
- Zhang, Y.J.; Zhang, S.; Liu, X.Z.; Wen, H.A.; Wang, M. A simple method of genomic DNA extraction suitable for analysis of bulk fungal strains. Lett. Appl. Microbiol. 2010, 51, 114–118. [Google Scholar] [CrossRef] [PubMed]
- BLAST: Basic Local Alignment Search Tool. Available online: https://blast.ncbi.nlm.nih.gov/Blast.cgi (accessed on 17 February 2021).
- Goedhart, J. SuperPlotsOfData—A web app for the transparent display and quantitative comparison of continuous data from discrete conditions. bioRxiv 2020. [Google Scholar] [CrossRef]
- Dussart, F.; Creissen, H.E.; Havis, N.D. Ramularia collo-cygni—An Enemy in Waiting. eLS 2020, 1–8. [Google Scholar] [CrossRef]
- Assinger, T.; Torriani, S.F.F.; Accardo, S.; Fountaine, J.; Steinkellner, S. Fungicide strategies and resistance of Ramularia collo-cygni to demethylation and succinate dehydrogenase inhibitors in Austrian winter barley (Hordeum vulgare). J. Plant Dis. Prot. 2021, 128, 735–748. [Google Scholar] [CrossRef]
- Heick, T.M.; Matzen, N.; Jørgensen, L.N. Reduced field efficacy and sensitivity of demethylation inhibitors in the Danish and Swedish Zymoseptoria tritici populations. Eur. J. Plant Pathol. 2020, 157, 625–636. [Google Scholar] [CrossRef]
- Kiiker, R.; Juurik, M.; Heick, T.M.; Mäe, A. Changes in DMI, SDHI, and QoI fungicide sensitivity in the Estonian Zymoseptoria tritici population between 2019 and 2020. Microorganisms 2021, 9, 814. [Google Scholar] [CrossRef] [PubMed]
- Crous, P.W.; Aptroot, A.; Kang, J.-C.; Braun, U.; Wingfield, M.J. Mycosphaerella and its anamorphs. Stud. Mycol. 2000, 45, 107–121. [Google Scholar]
- Matsuzaki, Y.; Kiguchi, S.; Suemoto, H.; Iwahashi, F. Antifungal activity of metyltetraprole against the existing QoI-resistant isolates of various plant pathogenic fungi: Metyltetraprole against QoI-R isolates. Pest Manag. Sci. 2020, 76, 1743–1750. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| County * | Fields | Mutations | Fungicide Sensitivity Assay | |||
|---|---|---|---|---|---|---|
| 2019 | 2020 | 2019 | 2020 | 2019 | 2020 | |
| IV | NA ** | 5 | NA | 64 | NA | 7 |
| JÕ | 2 | 3 | 9 | 35 | 5 | 15 |
| JÄ | NA | 7 | NA | 36 | NA | 8 |
| LV | 3 | 1 | 17 | 11 | 8 | 7 |
| PÕ | NA | 3 | NA | 15 | NA | 15 |
| RA | NA | 1 | NA | 6 | NA | 6 |
| TA | 4 | 6 | 20 | 63 | 12 | 20 |
| VA | 1 | NA | 5 | NA | 5 | NA |
| VI | 1 | 5 | 5 | 48 | NA | 30 |
| VÕ | 3 | 1 | 12 | 7 | 8 | 7 |
| TOTAL | 14 | 32 | 68 | 285 | 38 | 115 |
| County * | CYP51 Mutation Frequency (%) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| I381T | I384T | S459C | S459Y | S459T | S459L | ||||
| 2019 | 2020 | 2019 | 2020 | 2019 | 2020 | 2020 | 2019 | 2019 | |
| IV | NA ** | 100 | NA | 100 | NA | 100 | 0 | NA | NA |
| JÕ | 97 | 78 | 97 | 78 | 89 | 46 | 54 | 0 | 0 |
| JÄ | NA | 100 | NA | 100 | NA | 37 | 63 | NA | NA |
| LV | 100 | 100 | 100 | 100 | 100 | 100 | 0 | 0 | 0 |
| PÕ | NA | 100 | NA | 100 | NA | 86 | 0 | NA | NA |
| RA | NA | 100 | NA | 100 | NA | 50 | 17 | NA | NA |
| TA | 100 | 70 | 100 | 70 | 89 | 100 | 0 | 11 | 0 |
| VA | 100 | NA | 100 | NA | 40 | NA | NA | 60 | 0 |
| VI | NA | 90 | NA | 90 | NA | 88 | 12 | NA | NA |
| VÕ | 83 | 100 | 83 | 100 | 83 | 100 | 0 | 0 | 17 |
| Average | 96 | 93 | 96 | 93 | 80 | 79 | 16 | 14 | 3 |
| County * | SdhC Mutation Frequency (%) | |||||||
|---|---|---|---|---|---|---|---|---|
| H146R | N164H | G167C | V184L | |||||
| 2019 | 2020 | 2019 | 2020 | 2019 | 2020 | 2019 | 2020 | |
| IV | NA ** | 88 | NA | 13 | NA | 13 | NA | 50 |
| JÕ | 43 | 50 | 0 | 10 | 0 | 10 | 11 | 20 |
| JÄ | NA | 42 | NA | 21 | NA | 21 | NA | 21 |
| LV | 88 | 0 | 0 | 100 | 0 | 100 | 0 | 0 |
| PÕ | NA | 100 | NA | 0 | 0 | 0 | NA | 0 |
| RA | NA | 67 | NA | 50 | NA | 50 | NA | 50 |
| TA | 85 | 36 | 0 | 7 | 0 | 7 | 0 | 0 |
| VA | 40 | NA | 0 | NA | 0 | NA | 0 | NA |
| VI | NA | 57 | NA | 50 | NA | 50 | NA | 50 |
| VÕ | 58 | 58 | 0 | 0 | 0 | 10 | 0 | 0 |
| Average | 63 | 55 | 0 | 28 | 0 | 29 | 2 | 21 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kiiker, R.; Juurik, M.; Mäe, A. Fungicide Resistance Evolving in Ramularia collo-cygni Population in Estonia. Microorganisms 2021, 9, 1514. https://doi.org/10.3390/microorganisms9071514
Kiiker R, Juurik M, Mäe A. Fungicide Resistance Evolving in Ramularia collo-cygni Population in Estonia. Microorganisms. 2021; 9(7):1514. https://doi.org/10.3390/microorganisms9071514
Chicago/Turabian StyleKiiker, Riinu, Marite Juurik, and Andres Mäe. 2021. "Fungicide Resistance Evolving in Ramularia collo-cygni Population in Estonia" Microorganisms 9, no. 7: 1514. https://doi.org/10.3390/microorganisms9071514