
Candida albicans Biofilm Inhibition by Two Vaccinium macrocarpon (Cranberry) Urinary Metabolites: 5-(3′,4′-DihydroxyPhenyl)-γ-Valerolactone and 4-Hydroxybenzoic Acid

 , , , , , , and
, , , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Culture Conditions and Standardization
2.2. Crystal Violet Assay for Biofilm Quantification
2.3. XTT Reduction Assay
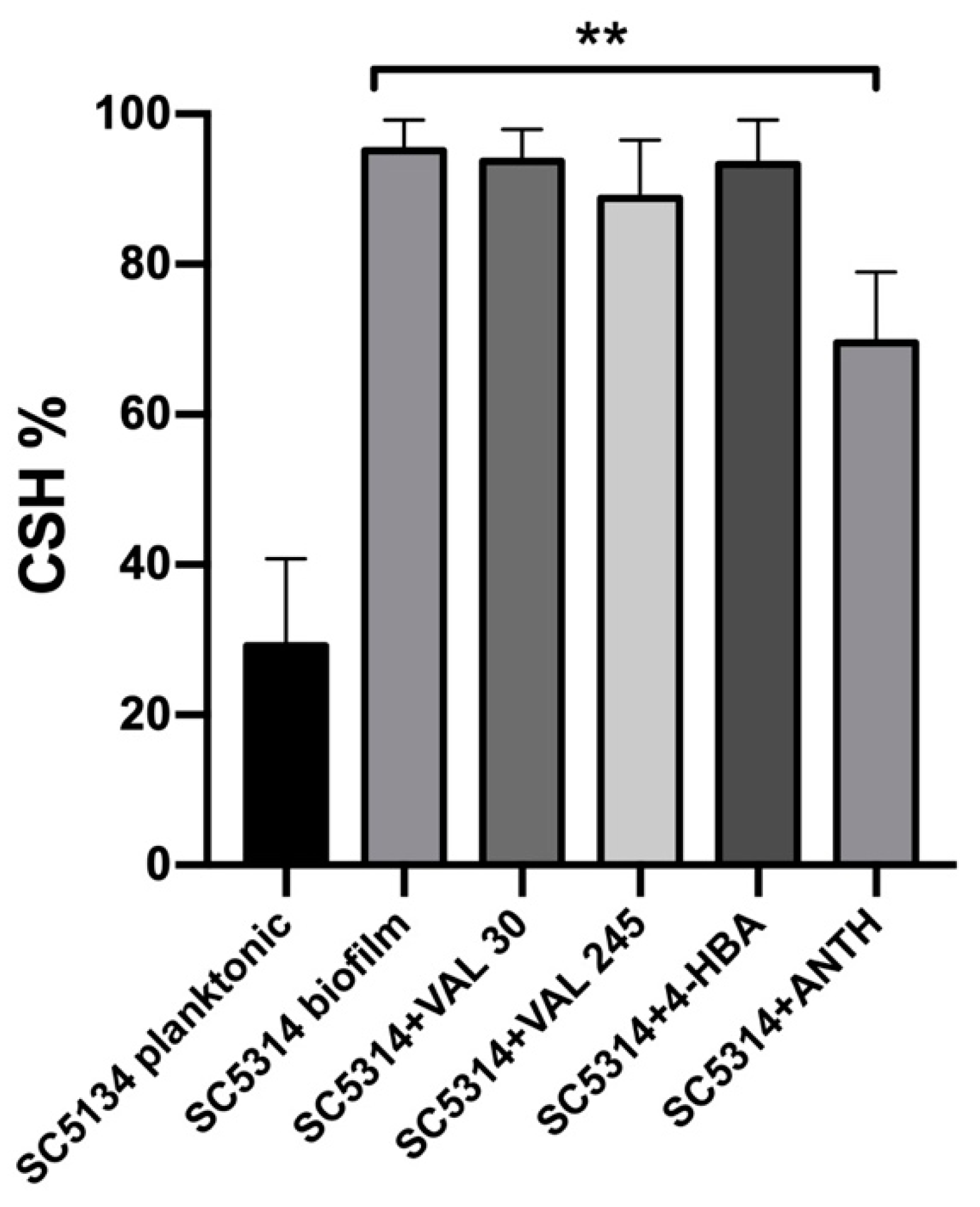
2.4. Cell Surface Hydrophobicity Assay
2.5. Confocal Laser Scanning Microscopy (CLSM)
2.6. CaCo-2 Culture and Cytotoxicity Assay
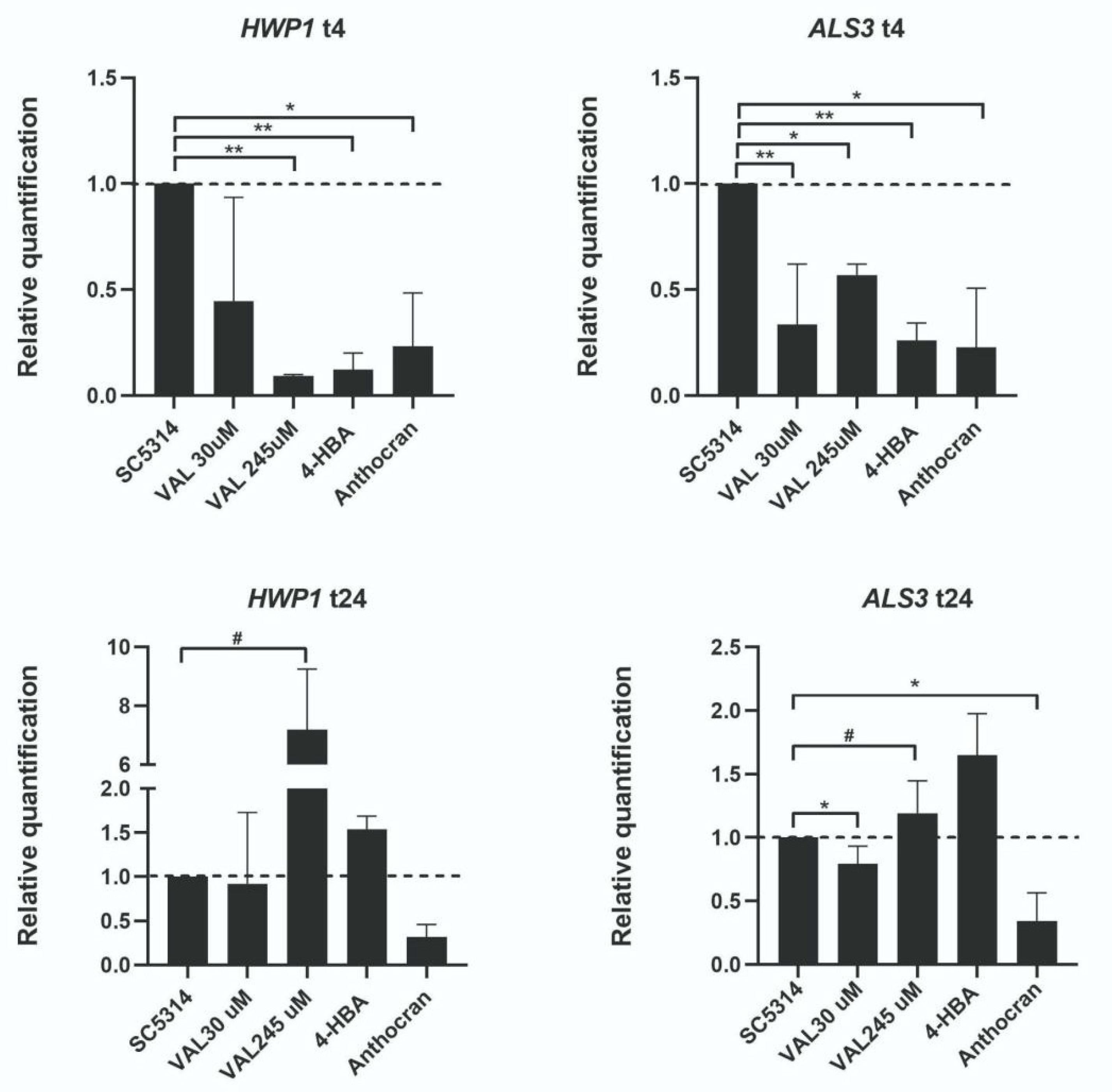
2.7. Gene Expression Analysis
2.8. Statistical Analysis
3. Results
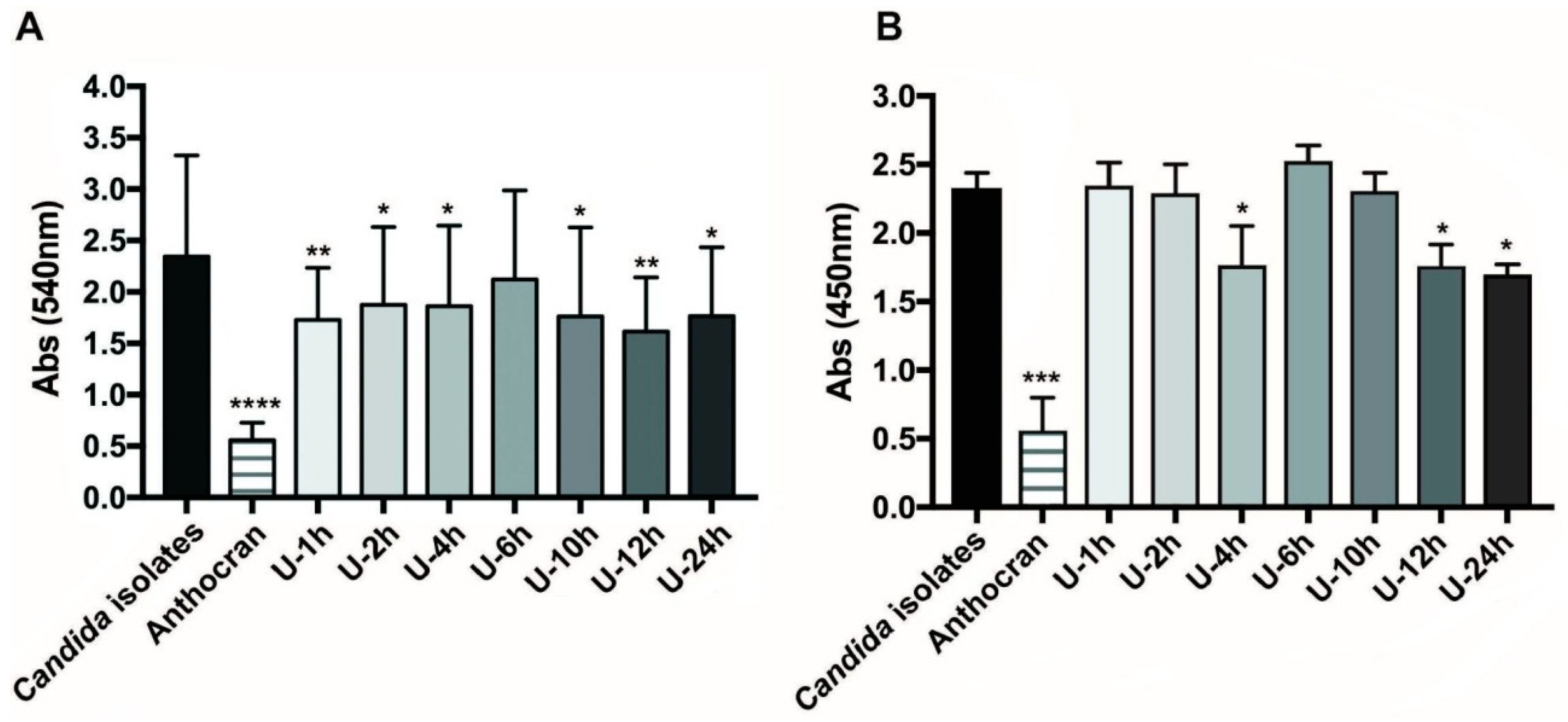
3.1. Inhibition of Biofilm-Formation of C. albicans Clinical Strain by Urine Fractions
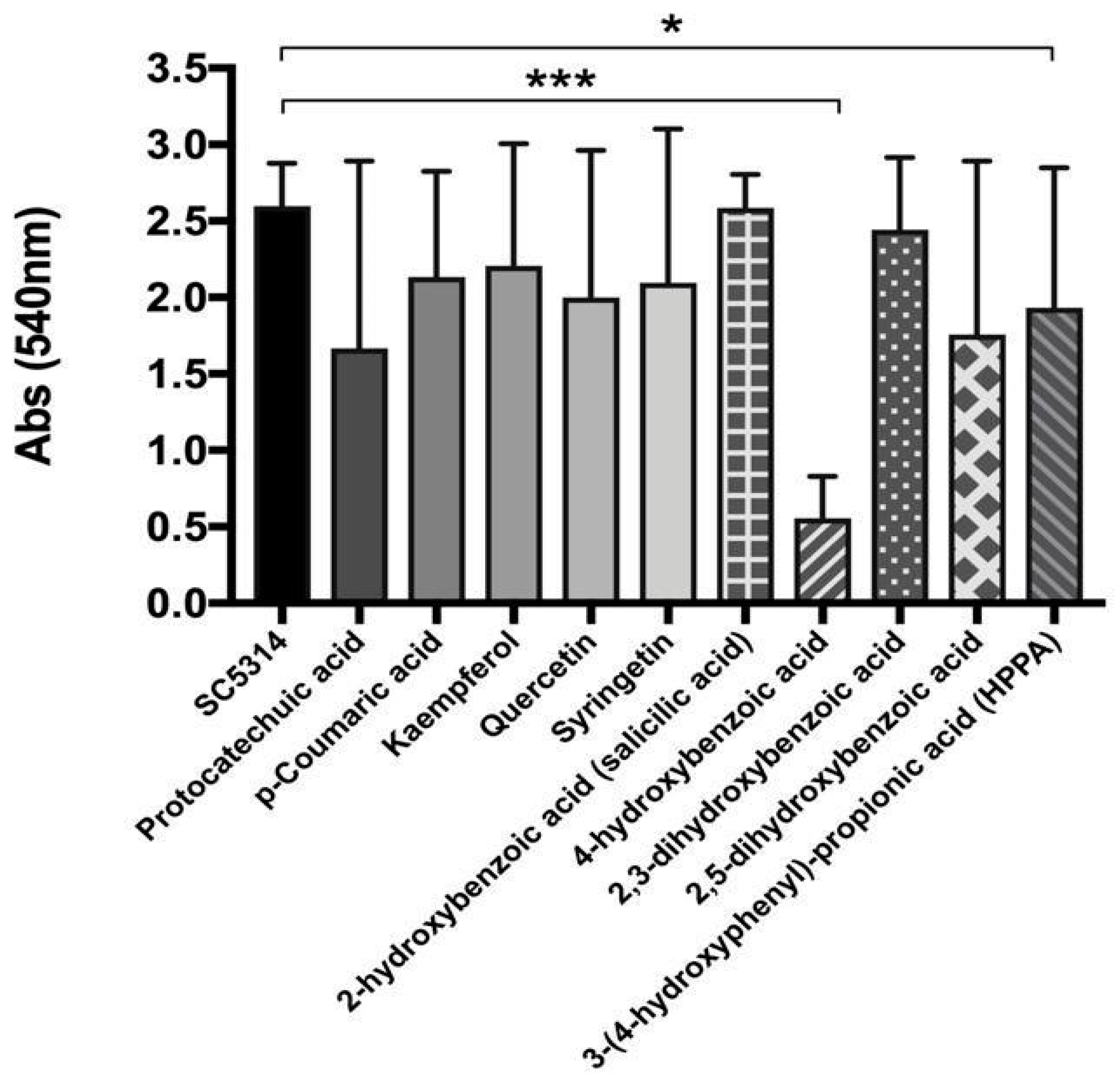
3.2. Cranberry Metabolites Activity Against C. albicans Biofilm Formation
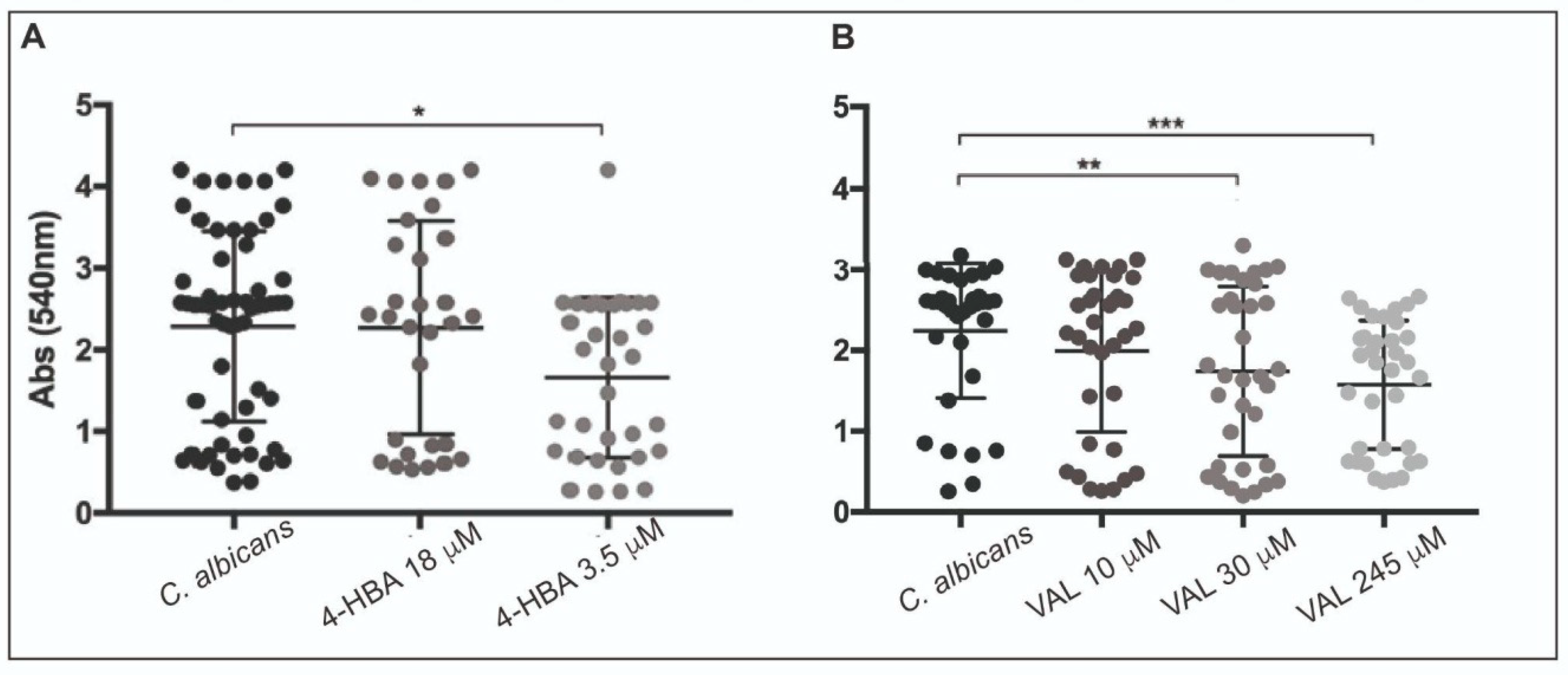
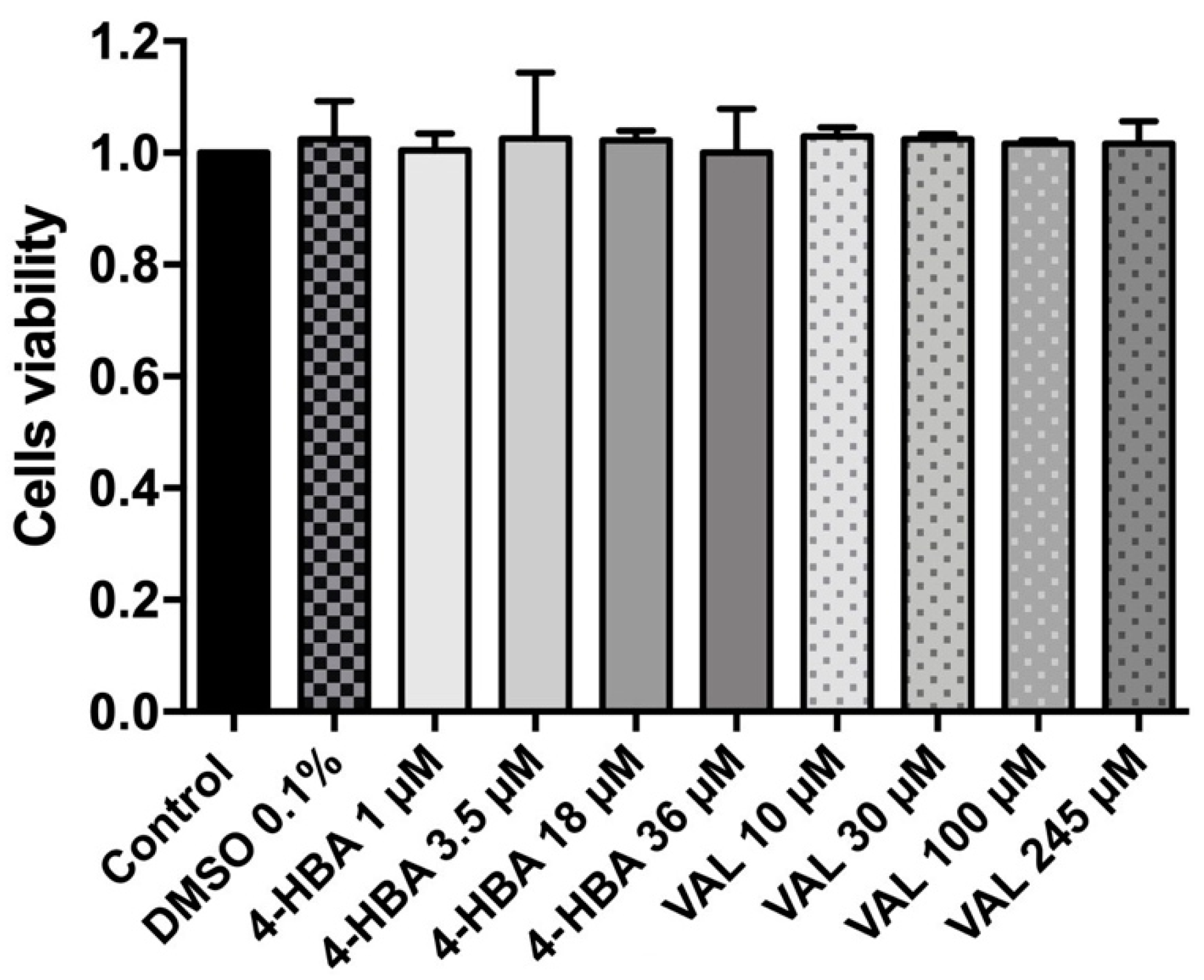
3.3. 4-HBA and VAL Cytotoxicity on Intestinal Cells
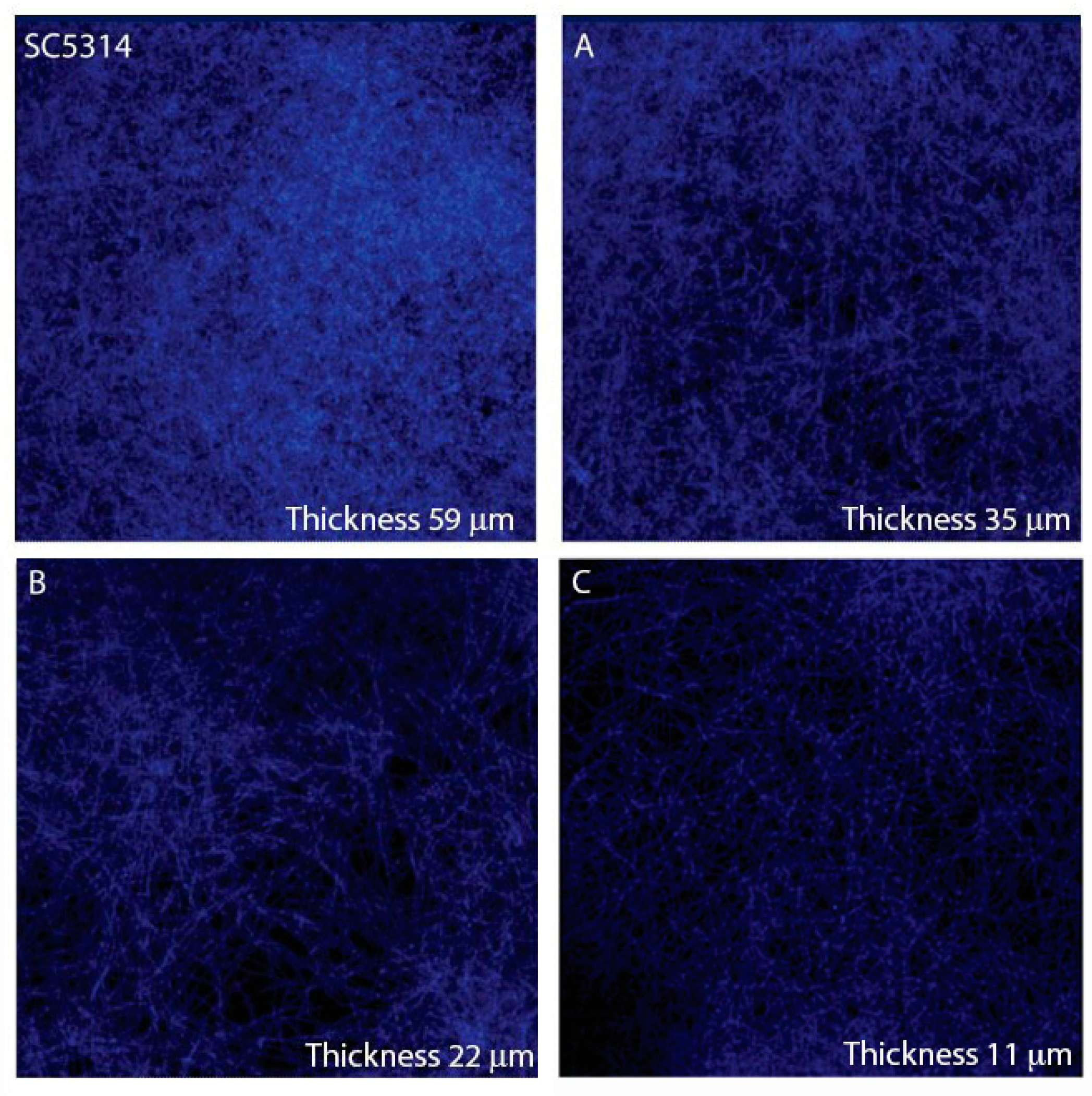
3.4. 4-HBA and VAL Inhibitory Activity on Candida Adhesion
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- d’Enfert, C.; Kaune, A.-K.; Alaban, L.-R.; Chakraborty, S.; Cole, N.; Delavy, M.; Kosmala, D.; Marsaux, B.; Fróis-Martins, R.; Morelli, M.; et al. The Impact of the Fungus-Host-Microbiota Interplay upon Candida Albicans Infections: Current Knowledge and New Perspectives. FEMS Microbiol. Rev. 2020. [Google Scholar] [CrossRef]
- Romani, L. Immunity to Fungal Infections. Nat. Rev. Immunol. 2004, 4, 11–24. [Google Scholar] [CrossRef]
- Mayer, F.L.; Wilson, D.; Hube, B. Candida Albicans Pathogenicity Mechanisms. Virulence 2013, 4, 119–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altomare, A.; Baron, G.; Brioschi, M.; Longoni, M.; Butti, R.; Valvassori, E.; Tremoli, E.; Carini, M.; Agostoni, P.; Vistoli, G.; et al. N-Acetyl-Cysteine Regenerates Albumin Cys34 by a Thiol-Disulfide Breaking Mechanism: An Explanation of Its Extracellular Antioxidant Activity. Antioxidants 2020, 9, 367. [Google Scholar] [CrossRef] [PubMed]
- Calderone, R.A.; Fonzi, W.A. Virulence Factors of Candida Albicans. Trends Microbiol. 2001, 9, 327–335. [Google Scholar] [CrossRef]
- Fisher, J.F.; Kavanagh, K.; Sobel, J.D.; Kauffman, C.A.; Newman, C.A. Candida Urinary Tract Infection: Pathogenesis. Clin. Infect. Dis. 2011, 52, S437–S451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raja, N.S. Epidemiology, Risk Factors, Treatment and Outcome of Candida Bloodstream Infections because of Candida Albicans and Candida Non-Albicans in Two District General Hospitals in the United Kingdom. Int. J. Clin. Pract. 2021, 75, e13655. [Google Scholar] [CrossRef]
- Gonçalves, B.; Ferreira, C.; Alves, C.T.; Henriques, M.; Azeredo, J.; Silva, S. Vulvovaginal Candidiasis: Epidemiology, Microbiology and Risk Factors. Crit. Rev. Microbiol. 2016, 42, 905–927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roselletti, E.; Monari, C.; Sabbatini, S.; Perito, S.; Vecchiarelli, A.; Sobel, J.D.; Cassone, A. A Role for Yeast/Pseudohyphal Cells of Candida albicans in the Correlated Expression of NLRP3 Inflammasome Inducers in Women with Acute Vulvovaginal Candidiasis. Front. Microbiol. 2019, 10, 2669. [Google Scholar] [CrossRef]
- Sobel, J.D. Recurrent Vulvovaginal Candidiasis. Am. J. Obstet. Gynecol. 2016, 214, 15–21. [Google Scholar] [CrossRef]
- Baron, G.; Altomare, A.; Regazzoni, L.; Fumagalli, L.; Artasensi, A.; Borghi, E.; Ottaviano, E.; Del Bo, C.; Riso, P.; Allegrini, P.; et al. Profiling Vaccinium Macrocarpon Components and Metabolites in Human Urine and the Urine Ex-Vivo Effect on Candida albicans Adhesion and Biofilm-Formation. Biochem. Pharmacol. 2020, 173, 113726. [Google Scholar] [CrossRef]
- Morace, G.; Polonelli, L. GISIA Group. Voriconazole Activity against Clinical Yeast Isolates: A Multicentre Italian Study. Int. J. Antimicrob. Agents 2005, 26, 247–253. [Google Scholar] [CrossRef]
- Sherry, L.; Rajendran, R.; Lappin, D.F.; Borghi, E.; Perdoni, F.; Falleni, M.; Tosi, D.; Smith, K.; Williams, C.; Jones, B.; et al. Biofilms Formed by Candida Albicans Bloodstream Isolates Display Phenotypic and Transcriptional Heterogeneity That Are Associated with Resistance and Pathogenicity. BMC Microbiol. 2014, 14, 182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcos-Zambrano, L.J.; Escribano, P.; Bouza, E.; Guinea, J. Production of Biofilm by Candida and Non-Candida Spp. Isolates Causing Fungemia: Comparison of Biomass Production and Metabolic Activity and Development of Cut-off Points. Int. J. Med. Microbiol. 2014, 304, 1192–1198. [Google Scholar] [CrossRef] [PubMed]
- Artasensi, A.; Baron, G.; Vistoli, G.; Aldini, G.; Fumagalli, L. (Z)-5-(3′,4′-Bis(benzyloxy)benzylidene)furan-2(5H)-One. Molbank 2021, 2021, M1193. [Google Scholar] [CrossRef]
- Pierce, C.G.; Uppuluri, P.; Tristan, A.R.; Wormley, F.L., Jr. A Simple and Reproducible 96-Well Plate-Based Method for the Formation of Fungal Biofilms and Its Application to Antifungal Susceptibility Testing. Nat. Protoc. 2008, 3, 1494–1500. [Google Scholar] [CrossRef] [PubMed]
- Peron, G.; Sut, S.; Pellizzaro, A.; Brun, P.; Voinovich, D.; Castagliuolo, I.; Dall’Acqua, S. The Antiadhesive Activity of Cranberry Phytocomplex Studied by Metabolomics: Intestinal PAC-A Metabolites but Not Intact PAC-A Are Identified as Markers in Active Urines against Uropathogenic Escherichia coli. Fitoterapia 2017, 122, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Ermel, G.; Georgeault, S.; Inisan, C.; Besnard, M. Inhibition of Adhesion of Uropathogenic Escherichia Coli Bacteria to Uroepithelial Cells by Extracts from Cranberry. J. Med. Food 2012, 15, 126–134. [Google Scholar] [CrossRef] [PubMed]
- Gupta, K.; Chou, M.Y.; Howell, A.; Wobbe, C.; Grady, R.; Stapleton, A.E. Cranberry Products Inhibit Adherence of P-Fimbriated Escherichia Coli to Primary Cultured Bladder and Vaginal Epithelial Cells. J. Urol. 2007, 177, 2357–2360. [Google Scholar] [CrossRef] [Green Version]
- Feliciano, R.P.; Boeres, A.; Massacessi, L.; Istas, G.; Ventura, M.R.; Nunes Dos Santos, C.; Heiss, C.; Rodriguez-Mateos, A. Identification and Quantification of Novel Cranberry-Derived Plasma and Urinary (poly)phenols. Arch. Biochem. Biophys. 2016, 599, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Iswaldi, I.; Arráez-Román, D.; Gómez-Caravaca, A.M.; Contreras, M.D.M.; Uberos, J.; Segura-Carretero, A.; Fernández-Gutiérrez, A. Identification of Polyphenols and Their Metabolites in Human Urine after Cranberry-Syrup Consumption. Food Chem. Toxicol. 2013, 55, 484–492. [Google Scholar] [CrossRef]
- McKay, D.L.; Chen, C.-Y.O.; Zampariello, C.A.; Blumberg, J.B. Flavonoids and Phenolic Acids from Cranberry Juice Are Bioavailable and Bioactive in Healthy Older Adults. Food Chem. 2015, 168, 233–240. [Google Scholar] [CrossRef]
- Walsh, J.M.; Ren, X.; Zampariello, C.; Polasky, D.A.; McKay, D.L.; Blumberg, J.B.; Chen, C.-Y.O. Liquid Chromatography with Tandem Mass Spectrometry Quantification of Urinary Proanthocyanin A2 Dimer and Its Potential Use as a Biomarker of Cranberry Intake. J. Sep. Sci. 2016, 39, 342–349. [Google Scholar] [CrossRef] [PubMed]
- Phua, A.I.-H.; Hon, K.Y.; Holt, A.; O’Callaghan, M.; Bihari, S. Candida Catheter-Related Bloodstream Infection in Patients on Home Parenteral Nutrition—Rates, Risk Factors, Outcomes, and Management. Clin. Nutr. ESPEN 2019, 31, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Tsui, C.; Kong, E.F.; Jabra-Rizk, M.A. Pathogenesis of Candida Albicans Biofilm. Pathog. Dis. 2016, 74, ftw018. [Google Scholar] [CrossRef] [Green Version]
- Janek, T.; Łukaszewicz, M.; Krasowska, A. Antiadhesive Activity of the Biosurfactant Pseudofactin II Secreted by the Arctic Bacterium Pseudomonas Fluorescens BD5. BMC Microbiol. 2012, 12, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piktel, E.; Suprewicz, Ł.; Depciuch, J.; Chmielewska, S.; Skłodowski, K.; Daniluk, T.; Król, G.; Kołat-Brodecka, P.; Bijak, P.; Pajor-Świerzy, A.; et al. Varied-Shaped Gold Nanoparticles with Nanogram Killing Efficiency as Potential Antimicrobial Surface Coatings for the Medical Devices. Sci. Rep. 2021, 11, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Vicariotto, F.; Del Piano, M.; Mogna, L.; Mogna, G. Effectiveness of the Association of 2 Probiotic Strains Formulated in a Slow Release Vaginal Product, in Women Affected by Vulvovaginal Candidiasis: A Pilot Study. J. Clin. Gastroenterol. 2012, 46, S73–S80. [Google Scholar] [CrossRef] [Green Version]
- Feldman, M.; Tanabe, S.; Howell, A.; Grenier, D. Cranberry Proanthocyanidins Inhibit the Adherence Properties of Candida Albicans and Cytokine Secretion by Oral Epithelial Cells. BMC Complement. Altern. Med. 2012, 12, 6. [Google Scholar]
- Sundararajan, A.; Rane, H.S.; Ramaraj, T.; Sena, J.; Howell, A.B.; Bernardo, S.M.; Schilkey, F.D.; Lee, S.A. Cranberry-Derived Proanthocyanidins Induce a Differential Transcriptomic Response within Candida Albicans Urinary Biofilms. PLoS ONE 2018, 13, e0201969. [Google Scholar] [CrossRef] [Green Version]
- Ou, K.; Sarnoski, P.; Schneider, K.R.; Song, K.; Khoo, C.; Gu, L. Microbial Catabolism of Procyanidins by Human Gut Microbiota. Mol. Nutr. Food Res. 2014, 58, 2196–2205. [Google Scholar] [CrossRef]
- Mena, P.; de Llano, D.G.; Brindani, N.; Esteban-Fernández, A.; Curti, C.; Moreno-Arribas, M.V.; Del Rio, D.; Bartolomé, B. 5-(3′,4′-Dihydroxyphenyl)-γ-Valerolactone and Its Sulphate Conjugates, Representative Circulating Metabolites of Flavan-3-Ols, Exhibit Anti-Adhesive Activity against Uropathogenic Escherichia Coli in Bladder Epithelial Cells. J. Funct. Foods 2017, 29, 275–280. [Google Scholar] [CrossRef]
- Zida, A.; Bamba, S.; Yacouba, A.; Ouedraogo-Traore, R.; Guiguemdé, R.T. Substances Naturelles Actives Sur Candida Albicans, Sources de Nouveaux Médicaments Antifongiques: Revue de La Littérature. J. Mycol. Med. 2017, 27, 1–19. [Google Scholar] [CrossRef]
- Nett, J.; Lincoln, L.; Marchillo, K.; Massey, R.; Holoyda, K.; Hoff, B.; VanHandel, M.; Andes, D. Putative Role of Beta-1,3 Glucans in Candida Albicans Biofilm Resistance. Antimicrob. Agents Chemother. 2007, 51, 510–520. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Rubio, R.; de Oliveira, H.C.; Rivera, J.; Trevijano-Contador, N. The Fungal Cell Wall: Candida, Cryptococcus, and Aspergillus Species. Front. Microbiol. 2019, 10, 2993. [Google Scholar] [CrossRef]
- Yoshijima, Y.; Murakami, K.; Kayama, S.; Liu, D.; Hirota, K.; Ichikawa, T.; Miyake, Y. Effect of Substrate Surface Hydrophobicity on the Adherence of Yeast and Hyphal Candida. Mycoses 2010, 53, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Silva-Dias, A.; Miranda, I.M.; Branco, J.; Monteiro-Soares, M.; Pina-Vaz, C.; Rodrigues, A.G. Adhesion, Biofilm Formation, Cell Surface Hydrophobicity, and Antifungal Planktonic Susceptibility: Relationship among Candida Spp. Front. Microbiol. 2015, 6, 205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borghi, E.; Sciota, R.; Biassoni, C.; Cirasola, D.; Cappelletti, L.; Vizzini, L.; Boracchi, P.; Morace, G. Cell Surface Hydrophobicity: A Predictor of Biofilm Production in Candida Isolates? J. Med. Microbiol. 2011, 60, 689–690. [Google Scholar] [CrossRef] [Green Version]
- Nobile, C.J.; Schneider, H.A.; Nett, J.E.; Sheppard, D.C.; Filler, S.G.; Andes, D.R.; Mitchell, A.P. Complementary Adhesin Function in C. albicans Biofilm Formation. Curr. Biol. 2008, 18, 1017–1024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nobile, C.J.; Nett, J.E.; Andes, D.R.; Mitchell, A.P. Function of Candida albicans Adhesin Hwp1 in Biofilm Formation. Eukaryot. Cell 2006, 5, 1604–1610. [Google Scholar] [CrossRef] [Green Version]
- Orsi, C.F.; Borghi, E.; Colombari, B.; Neglia, R.G.; Quaglino, D.; Ardizzoni, A.; Morace, G.; Blasi, E. Impact of Candida Albicans Hyphal Wall Protein 1 (HWP1) Genotype on Biofilm Production and Fungal Susceptibility to Microglial Cells. Microb. Pathog. 2014, 69–70, 20–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Staab, J.F.; Datta, K.; Rhee, P. Niche-Specific Requirement for Hyphal Wall Protein 1 in Virulence of Candida albicans. PLoS ONE 2013, 8, e80842. [Google Scholar]
- Silva, S.; Henriques, M.; Martins, A.; Oliveira, R.; Williams, D.; Azeredo, J. Biofilms of Non-Candida Albicans Candidaspecies: Quantification, Structure and Matrix Composition. Med. Mycol. 2009, 47, 681–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCall, A.D.; Pathirana, R.U.; Prabhakar, A.; Cullen, P.J.; Edgerton, M. Candida Albicans Biofilm Development Is Governed by Cooperative Attachment and Adhesion Maintenance Proteins. NPJ Biofilms Microbiomes 2019, 5, 21. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID Strain | Species | Source | Biofilm Production * |
|---|---|---|---|
| SC5314 | C. albicans | Reference strain | +++ |
| g11 | C. albicans | Vaginal swab | +++ |
| g23 | C. albicans | Vaginal swab | ++ |
| g29 | C. albicans | Vaginal swab | +++ |
| g35 | C. albicans | Vaginal swab | ++ |
| g44 | C. albicans | Vaginal swab | +++ |
| g49 | C. albicans | Vaginal swab | +/− |
| g67 | C. albicans | Vaginal swab | ++ |
| g69 | C. albicans | Urinary tract infection | +++ |
| g14 | C. albicans | Urinary tract infection | +++ |
| g53 | C. albicans | Urinary tract infection | +++ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ottaviano, E.; Baron, G.; Fumagalli, L.; Leite, J.; Colombo, E.A.; Artasensi, A.; Aldini, G.; Borghi, E. Candida albicans Biofilm Inhibition by Two Vaccinium macrocarpon (Cranberry) Urinary Metabolites: 5-(3′,4′-DihydroxyPhenyl)-γ-Valerolactone and 4-Hydroxybenzoic Acid. Microorganisms 2021, 9, 1492. https://doi.org/10.3390/microorganisms9071492
Ottaviano E, Baron G, Fumagalli L, Leite J, Colombo EA, Artasensi A, Aldini G, Borghi E. Candida albicans Biofilm Inhibition by Two Vaccinium macrocarpon (Cranberry) Urinary Metabolites: 5-(3′,4′-DihydroxyPhenyl)-γ-Valerolactone and 4-Hydroxybenzoic Acid. Microorganisms. 2021; 9(7):1492. https://doi.org/10.3390/microorganisms9071492
Chicago/Turabian StyleOttaviano, Emerenziana, Giovanna Baron, Laura Fumagalli, Jessica Leite, Elisa Adele Colombo, Angelica Artasensi, Giancarlo Aldini, and Elisa Borghi. 2021. "Candida albicans Biofilm Inhibition by Two Vaccinium macrocarpon (Cranberry) Urinary Metabolites: 5-(3′,4′-DihydroxyPhenyl)-γ-Valerolactone and 4-Hydroxybenzoic Acid" Microorganisms 9, no. 7: 1492. https://doi.org/10.3390/microorganisms9071492