
Two Pathway-Specific Transcriptional Regulators, PltR and PltZ, Coordinate Autoinduction of Pyoluteorin in Pseudomonas protegens Pf-5


Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Cultural Conditions
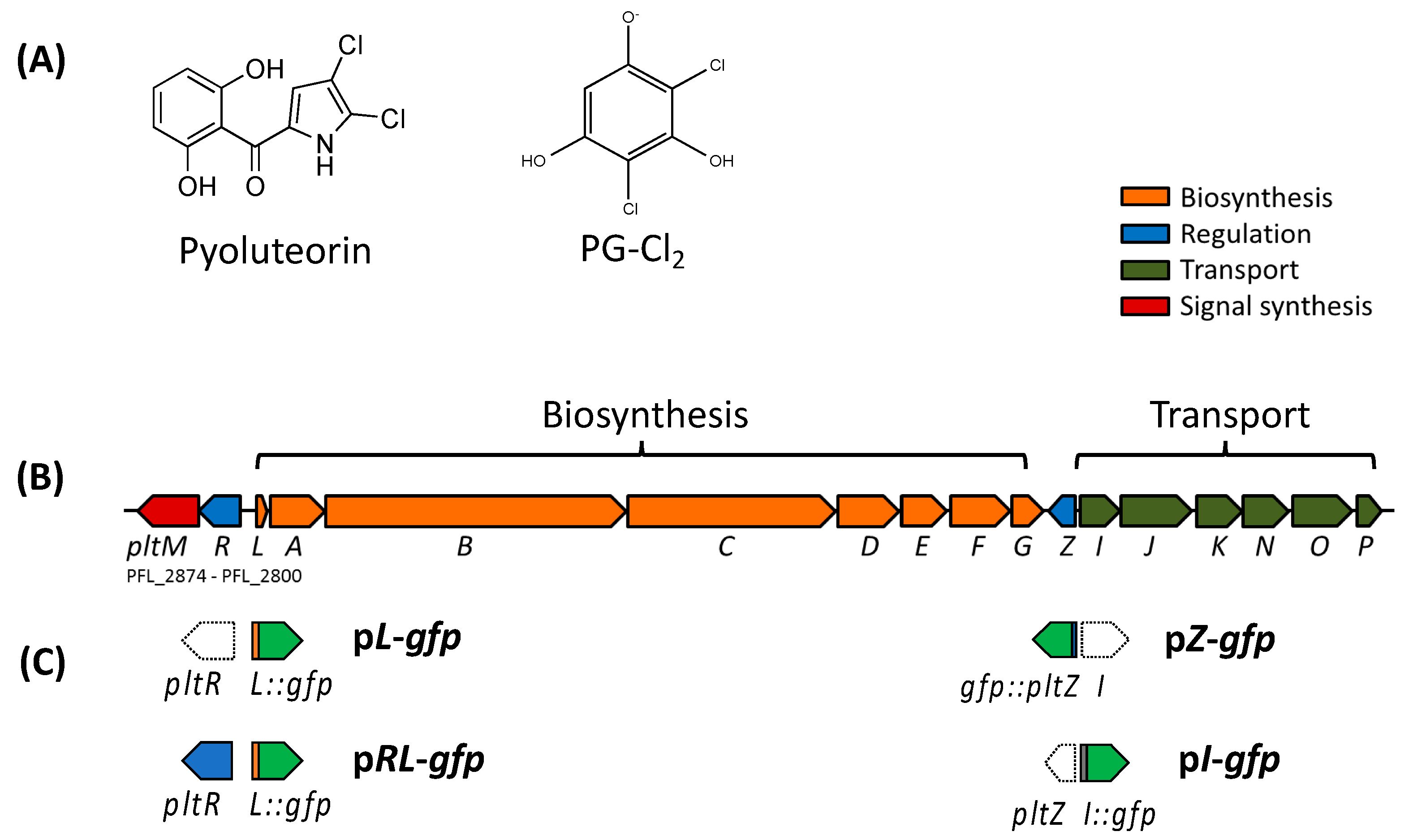
2.2. Construction of GFP Transcriptional Reporters and Expression Constructs
2.3. Construction of Pf-5 Mutants
2.4. Assays for Monitoring GFP Activity of Reporter Constructs
2.5. Protein Purification and In Vitro Activity Assays
3. Results
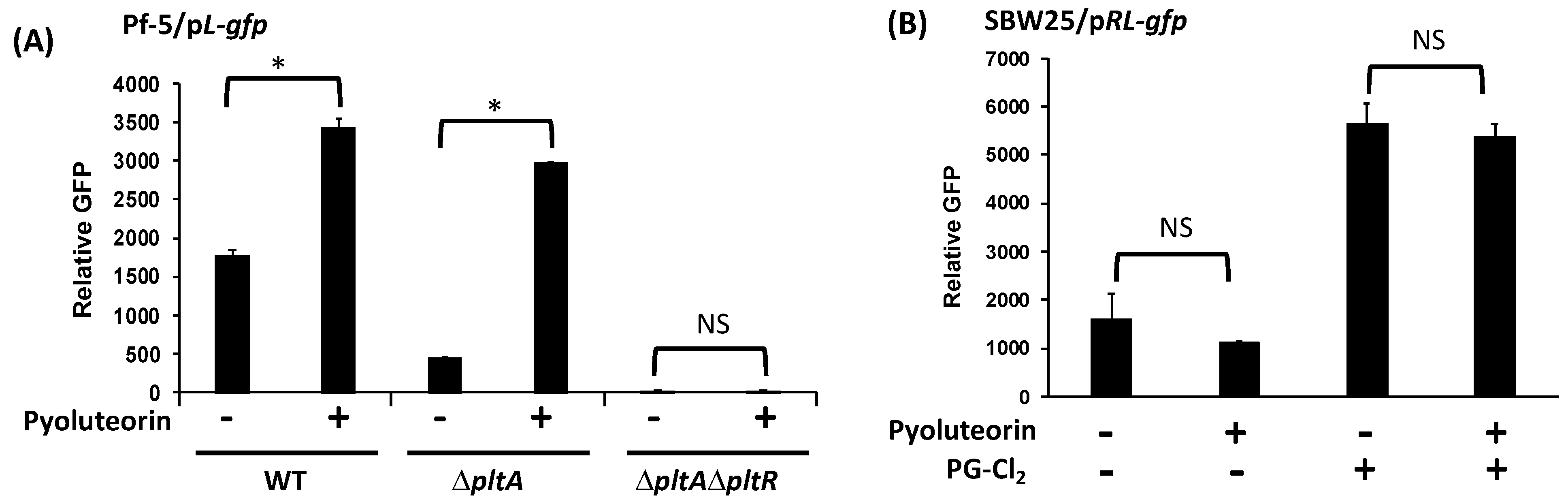
3.1. PltR Is Required But Not Sufficient for the Autoinduction of Pyoluteorin
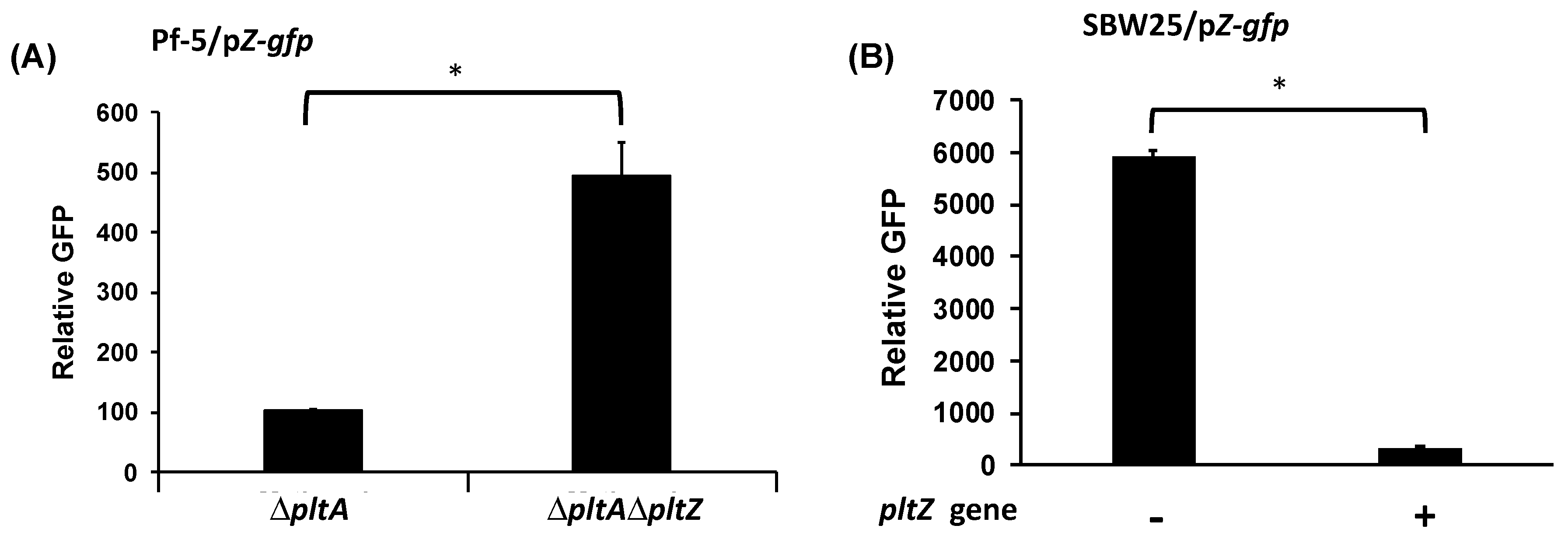
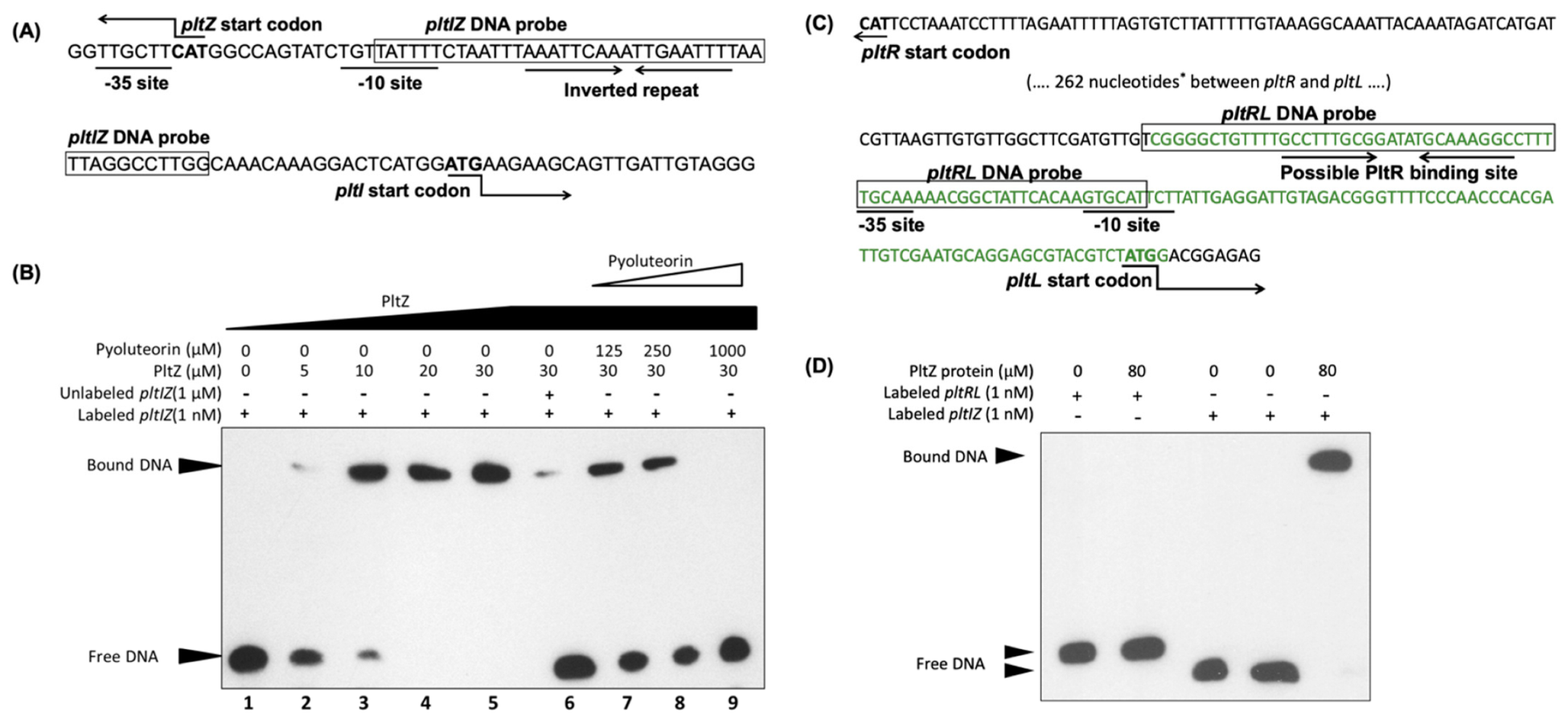
3.2. PltZ Is an Autorepressor and Directly Represses Expression of Linked Transporter Genes
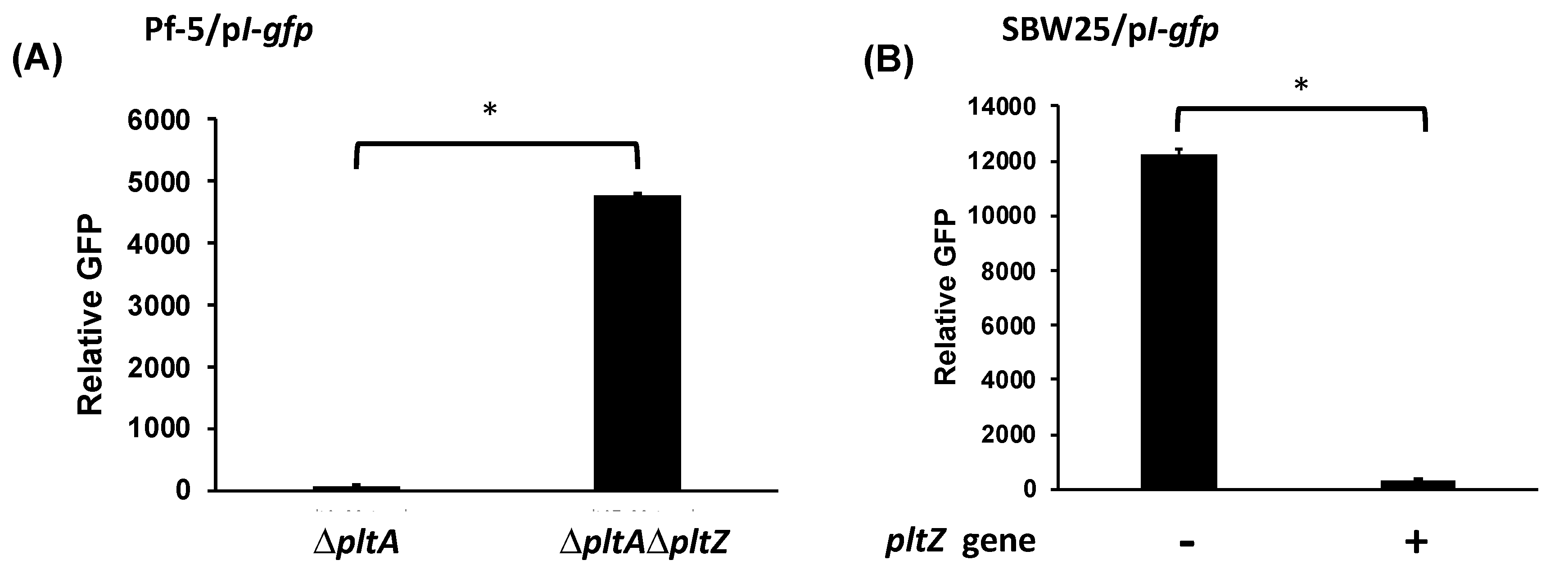
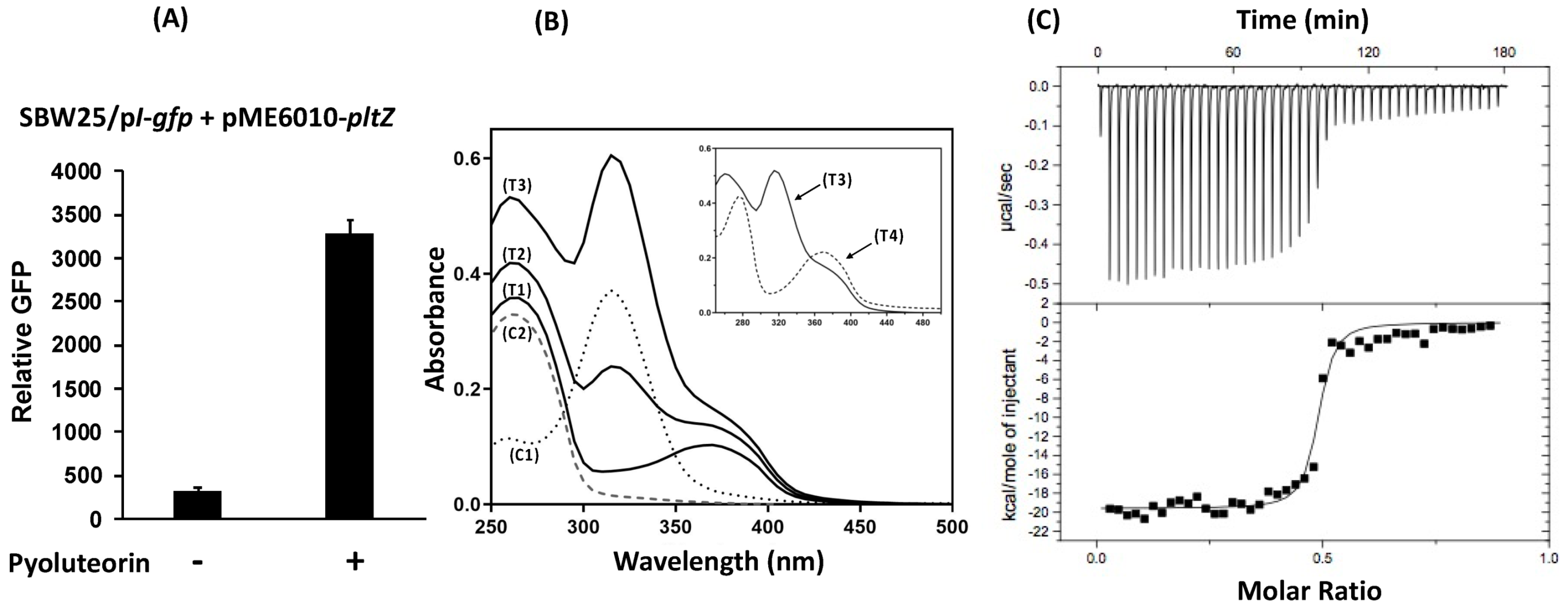
3.3. Pyoluteorin Interacts with PltZ and Relieves PltZ-Mediated Transcriptional Repression of pltIJKNOP Transporter Genes
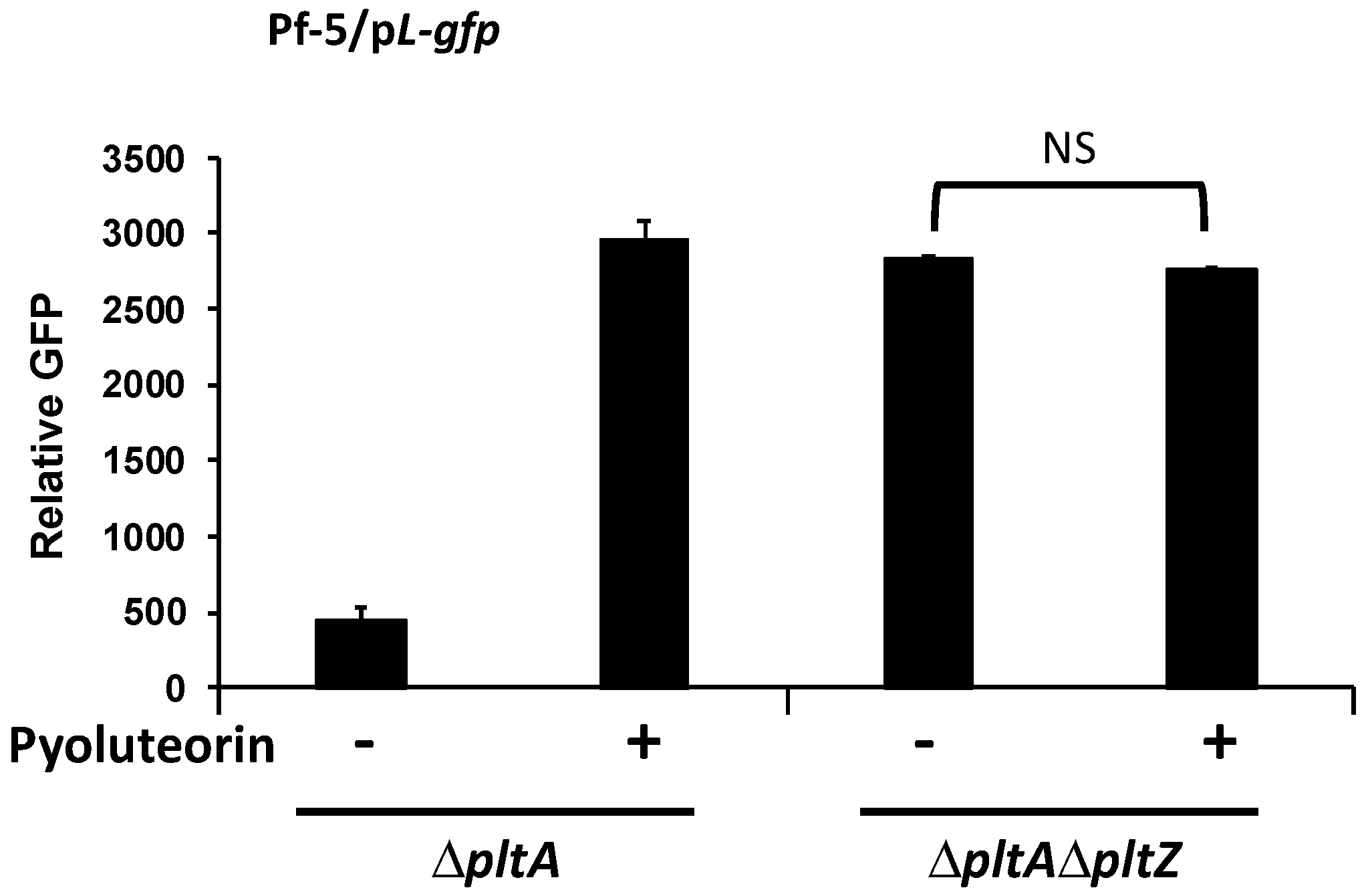
3.4. PltZ Is Involved in the Autoinduction of Pyoluteorin
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Kjelleberg, S.; Molin, S. Is There a Role for Quorum Sensing Signals in Bacterial Biofilms? Curr. Opin. Microbiol. 2002, 5, 254–258. [Google Scholar] [CrossRef]
- Smits, W.K.; Kuipers, O.P.; Veening, J.-W. Phenotypic Variation in Bacteria: The Role of Feedback Regulation. Nat. Rev. Microbiol. 2006, 4, 259–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Groisman, E.A. Feedback Control of Two-Component Regulatory Systems. Annu. Rev. Microbiol. 2016, 70, 103–124. [Google Scholar] [CrossRef] [PubMed]
- Camilli, A.; Bassler, B.L. Bacterial Small-Molecule Signaling Pathways. Science 2006, 311, 1113–1116. [Google Scholar] [CrossRef] [Green Version]
- Kuipers, O.P.; Beerthuyzen, M.M.; De Ruyter, P.G.; Luesink, E.J.; De Vos, W.M. Autoregulation of Nisin Biosynthesis in Lactococcus lactis by Signal Transduction. J. Biol. Chem. 1995, 270, 27299–27304. [Google Scholar] [CrossRef] [Green Version]
- Fish, S.A.; Cundliffe, E. Stimulation of Polyketide Metabolism in Streptomyces fradiae by Tylosin and Its Glycosylated Precursors. Microbiology 1997, 143, 3871–3876. [Google Scholar] [CrossRef] [Green Version]
- Schnider-Keel, U.; Seematter, A.; Maurhofer, M.; Blumer, C.; Duffy, B.; Gigot-Bonnefoy, C.; Reimmann, C.; Notz, R.; Défago, G.; Haas, D.; et al. Autoinduction of 2,4-Diacetylphloroglucinol Biosynthesis in the Biocontrol Agent Pseudomonas fluorescens CHA0 and Repression by the Bacterial Metabolites Salicylate and Pyoluteorin. J. Bacteriol. 2000, 182, 1215–1225. [Google Scholar] [CrossRef] [Green Version]
- Stein, T.; Borchert, S.; Kiesau, P.; Heinzmann, S.; Klöss, S.; Klein, C.; Helfrich, M.; Entian, K.-D. Dual Control of Subtilin Biosynthesis and Immunity in Bacillus subtilis. Mol. Microbiol. 2002, 44, 403–416. [Google Scholar] [CrossRef] [Green Version]
- Brodhagen, M.; Henkels, M.D.; Loper, J.E. Positive Autoregulation and Signaling Properties of Pyoluteorin, an Antibiotic Produced by the Biological Control Organism Pseudomonas fluorescens Pf-5. Appl. Environ. Microbiol. 2004, 70, 1758–1766. [Google Scholar] [CrossRef] [Green Version]
- Inaoka, T.; Takahashi, K.; Yada, H.; Yoshida, M.; Ochi, K. RNA Polymerase Mutation Activates the Production of a Dormant Antibiotic 3, 3′-Neotrehalosadiamine via an Autoinduction Mechanism in Bacillus subtilis. J. Biol. Chem. 2004, 279, 3885–3892. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Kim, J.-G.; Kang, Y.; Jang, J.Y.; Jog, G.J.; Lim, J.Y.; Kim, S.; Suga, H.; Nagamatsu, T.; Hwang, I. Quorum Sensing and the LysR-Type Transcriptional Activator ToxR Regulate Toxoflavin Biosynthesis and Transport in Burkholderia glumae. Mol. Microbiol. 2004, 54, 921–934. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Hutchinson, C.R. Feedback Regulation of Doxorubicin Biosynthesis in Streptomyces peucetius. Res. Microbiol. 2006, 157, 666–674. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, S.; Hoffmann, A.; Szekat, C.; Rudd, B.; Bierbaum, G. The Lantibiotic Mersacidin Is an Autoinducing Peptide. Appl. Environ. Microbiol. 2006, 72, 7270–7277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Tian, X.; Wang, J.; Yang, H.; Fan, K.; Xu, G.; Yang, K.; Tan, H. Autoregulation of Antibiotic Biosynthesis by Binding of the End Product to an Atypical Response Regulator. Proc. Natl. Acad. Sci. USA 2009, 106, 8617–8622. [Google Scholar] [CrossRef] [Green Version]
- Sherwood, E.J.; Bibb, M.J. The Antibiotic Planosporicin Coordinates Its Own Production in the Actinomycete planomonospora Alba. Proc. Natl. Acad. Sci. USA 2013, 110, E2500–E2509. [Google Scholar] [CrossRef] [Green Version]
- Abbas, A.; Morrissey, J.P.; Marquez, P.C.; Sheehan, M.M.; Delany, I.R.; O’Gara, F. Characterization of Interactions between the Transcriptional Repressor PhlF and Its Binding Site at the phlA Promoter in Pseudomonas fluorescens F113. J. Bacteriol. 2002, 184, 3008–3016. [Google Scholar] [CrossRef] [Green Version]
- Takeda, R. Pseudomonas Pigments. I. Pyoluteorin, a New Chlorine-Containing Pigment Produced by Pseudomonas aeruginosa. Hako Kogaku Zasshi 1958, 31, 281–290. [Google Scholar]
- Howell, C.R.; Stipanovic, R.D. Suppression of Pythium Ultimum-Induced Damping-off of Cotton Seedlings by Pseudomonas fluorescens and Its Antibiotic, Pyoluteorin. Phytopathology 1980, 70, 712–715. [Google Scholar] [CrossRef]
- Baehler, E.; Bottiglieri, M.; Péchy-Tarr, M.; Maurhofer, M.; Keel, C. Use of Green Fluorescent Protein-Based Reporters to Monitor Balanced Production of Antifungal Compounds in the Biocontrol Agent Pseudomonas fluorescens CHA0. J. Appl. Microbiol. 2005, 99, 24–38. [Google Scholar] [CrossRef]
- Ge, Y.H.; Zhao, Y.H.; Chen, L.J.; Miao, J.; Wen, L. Autoinduction of Pyoluteorin and Correlation between Phenazine-1-Carboxylic Acid and Pyoluteorin in Pseudomonas sp. M18. Wei Sheng Wu Xue Bao Acta Microbiol. Sin. 2007, 47, 441–446. [Google Scholar]
- Nowak-Thompson, B.; Chaney, N.; Wing, J.S.; Gould, S.J.; Loper, J.E. Characterization of the Pyoluteorin Biosynthetic Gene Cluster of Pseudomonas fluorescens Pf-5. J. Bacteriol. 1999, 181, 2166–2174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Huang, X.; Wang, G.; Xu, Y. Transcriptional Activation of Pyoluteorin Operon Mediated by the LysR-Type Regulator PltR Bound at a 22 Bp lys Box in Pseudomonas aeruginosa M18. PLoS ONE 2012, 7, e39538. [Google Scholar] [CrossRef] [Green Version]
- Maddocks, S.E.; Oyston, P.C. Structure and Function of the LysR-Type Transcriptional Regulator (LTTR) Family Proteins. Microbiology 2008, 154, 3609–3623. [Google Scholar] [CrossRef] [Green Version]
- Yan, Q.; Philmus, B.; Chang, J.H.; Loper, J.E. Novel Mechanism of Metabolic Co-Regulation Coordinates the Biosynthesis of Secondary Metabolites in Pseudomonas protegens. eLife 2017, 6, e22835. [Google Scholar] [CrossRef]
- King, E.O.; Ward, M.K.; Raney, D.E. Two Simple Media for the Demonstration of Pyocyanin and Fluorescin. J. Lab. Clin. Med. 1954, 44, 301–307. [Google Scholar]
- Howell, C.R.; Stipanovic, R.D. Control of Rhizoctonia Solani in Cotton Seedlings with Pseudomonas fluorescens and with an Antibiotic Produced by the Bacterium. Phytopathology 1979, 69, 480–482. [Google Scholar] [CrossRef] [Green Version]
- Henkels, M.D.; Kidarsa, T.A.; Shaffer, B.T.; Goebel, N.C.; Burlinson, P.; Mavrodi, D.V.; Bentley, M.A.; Rangel, L.I.; Davis, E.W.; Thomashow, L.S.; et al. Pseudomonas protegens Pf-5 Causes Discoloration and Pitting of Mushroom Caps Due to the Production of Antifungal Metabolites. Mol. Plant-Microbe Interact. 2014, 27, 733–746. [Google Scholar] [CrossRef] [Green Version]
- Silby, M.W.; Cerdeño-Tárraga, A.M.; Vernikos, G.S.; Giddens, S.R.; Jackson, R.W.; Preston, G.M.; Zhang, X.-X.; Moon, C.D.; Gehrig, S.M.; Godfrey, S.A. Genomic and Genetic Analyses of Diversity and Plant Interactions of Pseudomonas fluorescens. Genome Biol. 2009, 10, 1–16. [Google Scholar] [CrossRef]
- Simon, R.; Priefer, U.; Pühler, A. A Broad Host Range Mobilization System for in vivo Genetic Engineering: Transposon Mutagenesis in Gram Negative Bacteria. Nat. Biotechnol. 1983, 1, 784–791. [Google Scholar] [CrossRef]
- Hoang, T.T.; Karkhoff-Schweizer, R.R.; Kutchma, A.J.; Schweizer, H.P. A Broad-Host-Range Flp-FRT Recombination System for Site-Specific Excision of Chromosomally-Located DNA Sequences: Application for Isolation of Unmarked Pseudomonas aeruginosa Mutants. Gene 1998, 212, 77–86. [Google Scholar] [CrossRef]
- Miller, W.G.; Leveau, J.H.; Lindow, S.E. Improved gfp and inaZ Broad-Host-Range Promoter-Probe Vectors. Mol. Plant-Microbe Interact. 2000, 13, 1243–1250. [Google Scholar] [CrossRef] [Green Version]
- Yan, Q.; Philmus, B.; Hesse, C.; Kohen, M.; Chang, J.H.; Loper, J. The Rare Codon AGA Is Involved in Regulation of Pyoluteorin Biosynthesis in Pseudomonas protegens Pf-5. Front. Microbiol. 2016, 7, 497. [Google Scholar] [CrossRef] [Green Version]
- Heeb, S.; Itoh, Y.; Nishijyo, T.; Schnider, U.; Keel, C.; Wade, J.; Walsh, U.; O’Gara, F.; Haas, D. Small, Stable Shuttle Vectors Based on the Minimal PVS1 Replicon for Use in Gram-Negative, Plant-Associated Bacteria. Mol. Plant-Microbe Interact. 2000, 13, 232–237. [Google Scholar] [CrossRef] [Green Version]
- Loper, J.E.; Hassan, K.A.; Mavrodi, D.V.; Davis, E.W., II; Lim, C.K.; Shaffer, B.T.; Elbourne, L.D.; Stockwell, V.O.; Hartney, S.L.; Breakwell, K. Comparative Genomics of Plant-Associated Pseudomonas spp.: Insights into Diversity and Inheritance of Traits Involved in Multitrophic Interactions. PLoS Genet. 2012, 8, e1002784. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.-Q.; Ge, Y.-H.; Zhang, X.-H.; Xu, Y.Q. Transcriptional Repression of PltZ on Pyoluteorin ABC Transporter of Pseudomonas sp. M18. Wei Sheng Wu Xue Bao Acta Microbiol. Sin. 2005, 45, 344–348. [Google Scholar]
- Guo, D.-D.; Luo, L.-M.; Ma, H.-L.; Zhang, S.-P.; Xu, H.; Zhang, H.; Wang, Y.; Yuan, Y.; Wang, Z.; He, Y.-X. The Regulator PltZ Regulates a Putative ABC Transporter System PltIJKNOP of Pseudomonas aeruginosa ATCC 27853 in Response to the Antimicrobial 2, 4-Diacetylphloroglucinol. Front. Microbiol. 2020, 11, 1423. [Google Scholar] [CrossRef] [PubMed]
- Brodhagen, M.; Paulsen, I.; Loper, J.E. Reciprocal Regulation of Pyoluteorin Production with Membrane Transporter Gene Expression in Pseudomonas fluorescens Pf-5. Appl. Environ. Microbiol. 2005, 71, 6900–6909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuthbertson, L.; Nodwell, J.R. The TetR Family of Regulators. Microbiol. Mol. Biol. Rev. 2013, 77, 440–475. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Zhu, D.; Ge, Y.; Hu, H.; Zhang, X.; Xu, Y. Identification and Characterization of pltZ, a Gene Involved in the Repression of Pyoluteorin Biosynthesis in Pseudomonas sp. M18. FEMS Microbiol. Lett. 2004, 232, 197–202. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Yan, A.; Zhang, X.; Xu, Y. Identification and Characterization of a Putative ABC Transporter PltHIJKN Required for Pyoluteorin Production in Pseudomonas sp. M18. Gene 2006, 376, 68–78. [Google Scholar] [CrossRef]
- Kidarsa, T.A.; Goebel, N.C.; Zabriskie, T.M.; Loper, J.E. Phloroglucinol Mediates Crosstalk between the Pyoluteorin and 2,4-Diacetylphloroglucinol Biosynthetic Pathways in Pseudomonas fluorescens Pf-5. Mol. Microbiol. 2011, 81, 395–414. [Google Scholar] [CrossRef]
- Xu, Y.; Willems, A.; Au-yeung, C.; Tahlan, K.; Nodwell, J.R. A Two-Step Mechanism for the Activation of Actinorhodin Export and Resistance in Streptomyces coelicolor. MBio 2012, 3, e00191-12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, X.; Yang, R.; Zhao, R.-X.; Han, J.-T.; Jia, W.-J.; Li, D.-Y.; Wang, Y.; Zhang, N.; Wu, Y.; Zhang, L.-Q. Transcriptional Regulator PhlH Modulates 2, 4-Diacetylphloroglucinol Biosynthesis in Response to the Biosynthetic Intermediate and End Product. Appl. Environ. Microbiol. 2017, 83, e01419-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, D.; Wang, X.; Nie, J.; Niu, G. Regulation of Antibiotic Production by Signaling Molecules in Streptomyces. Front. Microbiol. 2019, 10, 2927. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains, Plasmids, or Primers | Genotype, Relevant Characteristics or Sequences | Reference or Source |
|---|---|---|
| Strains # | ||
| P. protegens | ||
| LK099 | Wild-type strain Pf-5. | [26] |
| JL4805 | Pf-5 derivative strain contains an in-frame deletion of pltA in the chromosome. | [27] |
| LK530 | Pf-5 derivative strain contains an in-frame deletion of pltA and pltR in the chromosome. This mutant was generated by deleting pltR in JL4805. | This study |
| LK419 | Pf-5 derivative strain contains an in-frame deletion of pltA and pltZ in the chromosome. This mutant was generated by deleting pltZ in JL4805. | This study |
| P. fluorescens SBW25 | SBW25 is a member of the P. fluorescens group that does not contain plt gene cluster. | [28] |
| E. coli | ||
| S17-1 | recA pro hsdR−M+ RP4 2-Tc::Mu-Km::Tn7 Smr Tpr | [29] |
| BL21 (DE3) | ompT hsdSB (rB−mB−) gal dcm | NEB |
| Plasmids | ||
| pEX18Tc | Gene replacement vector with MCS from pUC18, sacB+ Tcr | [30] |
| pEX18Tc-ΔpltR | pEX18Tc containing pltR with a 925-bp in-frame deletion. | [24] |
| pEX18Tc-ΔpltZ | pEX18Tc containing pltZ with a 621-bp in-frame deletion. | This study |
| pPROBE-NT | pBBR1, containing a promoterless gfp, Kmr | [31] |
| pL-gfp | Called ppltL-gfp previously. Contains the intergenic region between pltR and pltL including pltL promoter fused with a promoterless gfp. | [32] |
| pRL-gfp | Contains pltR and the intergenic region between pltR and pltL including pltL promoter fused with a promoterless gfp. | [24] |
| pI-gfp | Contains the intergenic region between pltI and pltZ including pltI promoter fused with a promoterless gfp. | This study |
| pZ-gfp | Contains the intergenic region between pltI and pltZ including pltZ promoter fused with a promoterless gfp. | This study |
| pME6010 | pACYC177-pVS1 shuttle vector, Tcr | [33] |
| PME6010-pltZ | pME6010 with a 697-bp XhoI-KpnI PCR fragment amplified from the genomic DNA of Pf-5, containing a constitutively expressed pltZ gene. | This study |
| Primers * | oligonucleotide sequences (5′to 3′) | |
| pltZ-F2 | ACTCGAGAAAATAACAGATACTGGCCATG | |
| pltZ-R2 | ATAGGTACCGACTATTGGGCAATGGC | |
| pltI-F1 | ATAGGATCCAGCAGACGGAATTTGGGC | |
| pltI-R1 | TATGGTACCCCTACAATCAACTGCTTCTTC | |
| pltI-F4 | TGGCGAATGTCGACGTTGC | |
| pltI-R4 | TATGAATTCCTCGTGGTCTGAGCGG | |
| pltZ 5′primer | GTCATACATATGAAGCAACCCCCCGCTC | |
| pltZ 3′primer | ATGATCTCGAGCCGACTATTGGGCAATG | |
| pltZ UpF-Xba | CTCCTCTCTAGATCAAAACGTGCCTTGGACTG | |
| pltZ UpR | ACTATTGGGCAATGATGTTCCTCGTGGTCTGAG | |
| pltZ DnF | ACGAGGAACATCATTGCCCAATAGTCGGCTCA | |
| pltZ DnR-Xba | CACACCTCTAGAACATCCCCTGCTGTTTCTTC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, Q.; Liu, M.; Kidarsa, T.; Johnson, C.P.; Loper, J.E. Two Pathway-Specific Transcriptional Regulators, PltR and PltZ, Coordinate Autoinduction of Pyoluteorin in Pseudomonas protegens Pf-5. Microorganisms 2021, 9, 1489. https://doi.org/10.3390/microorganisms9071489
Yan Q, Liu M, Kidarsa T, Johnson CP, Loper JE. Two Pathway-Specific Transcriptional Regulators, PltR and PltZ, Coordinate Autoinduction of Pyoluteorin in Pseudomonas protegens Pf-5. Microorganisms. 2021; 9(7):1489. https://doi.org/10.3390/microorganisms9071489
Chicago/Turabian StyleYan, Qing, Mary Liu, Teresa Kidarsa, Colin P. Johnson, and Joyce E. Loper. 2021. "Two Pathway-Specific Transcriptional Regulators, PltR and PltZ, Coordinate Autoinduction of Pyoluteorin in Pseudomonas protegens Pf-5" Microorganisms 9, no. 7: 1489. https://doi.org/10.3390/microorganisms9071489