
Predictive Modeling and Validation on Growth, Production of Asexual Spores and Ochratoxin A of Aspergillus Ochraceus Group under Abiotic Climatic Variables

 , , ,
, , ,  ,
,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Strains
2.2. Media Preparations
2.3. Culturing and Growth Rate Assessment
2.4. Measurements of Asexual Spores
2.5. Estimation of Ochratoxin A
2.6. Modeling of Fungal Growth, Sporulation, and Ochratoxin A
2.6.1. Normalization
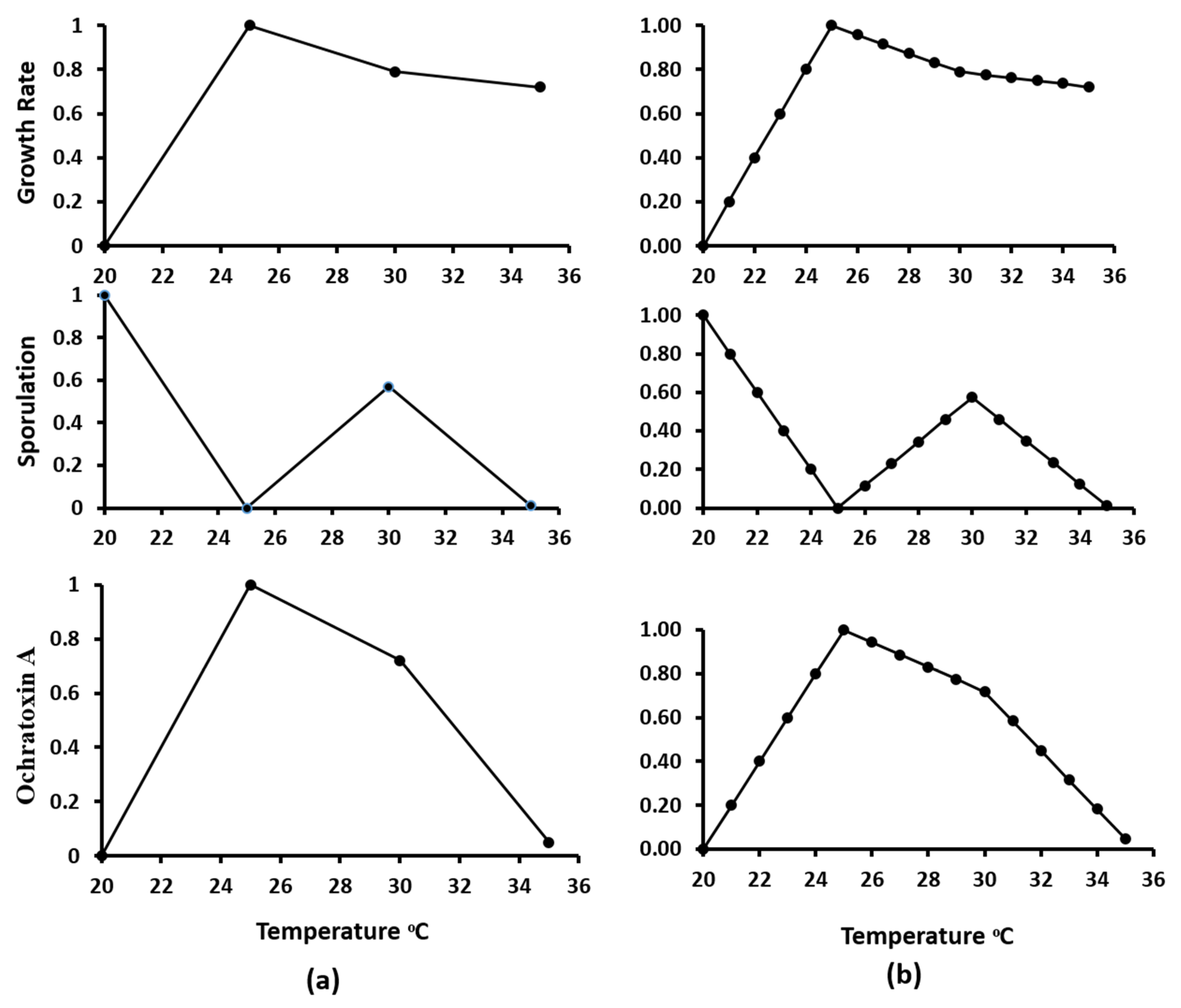
2.6.2. Linear Interpolation
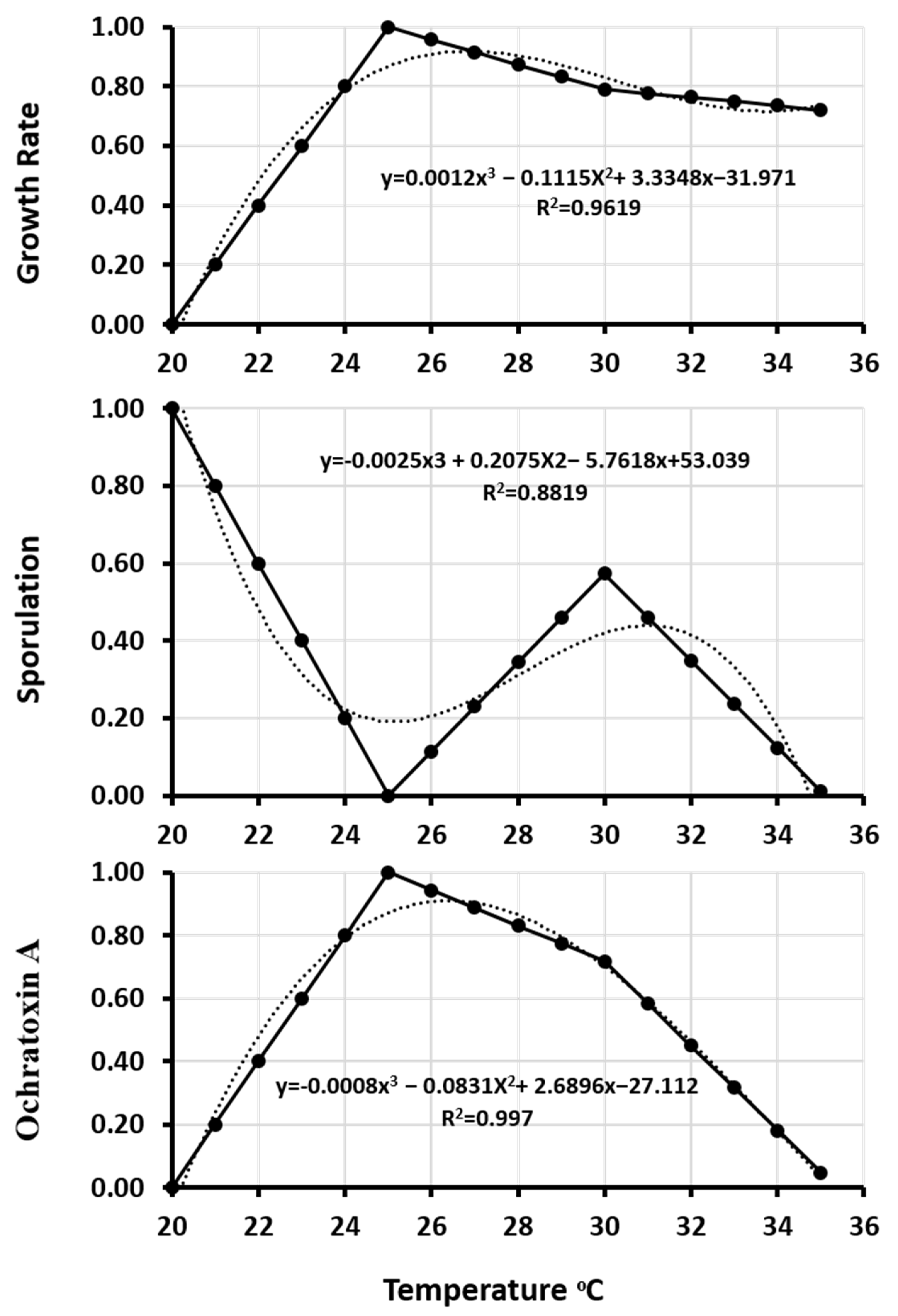
2.6.3. Polynomial Regression
2.6.4. Nonlinear Regression
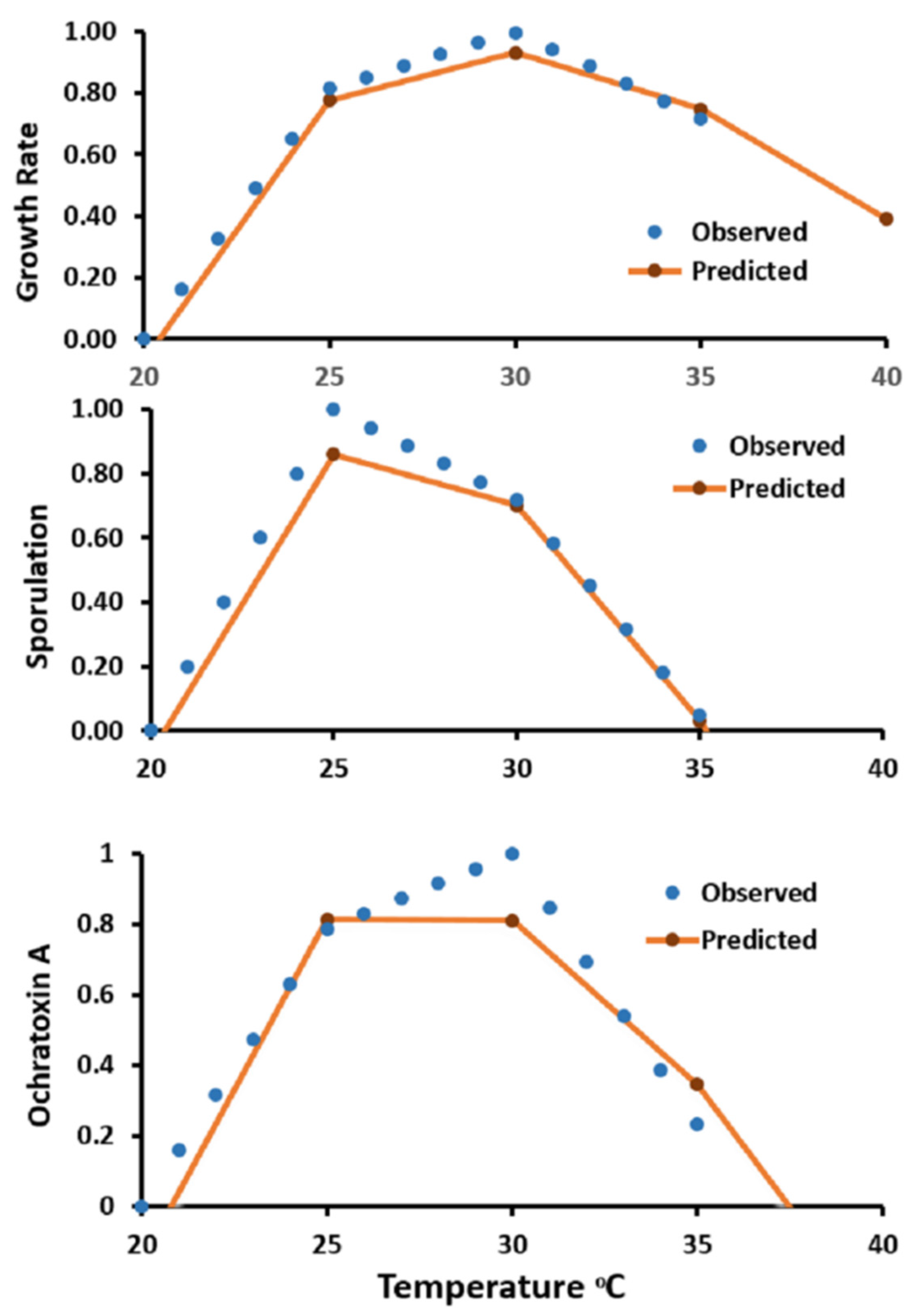
2.7. Model Testing and Validation
2.8. Statistical Analysis
3. Results and Discussion
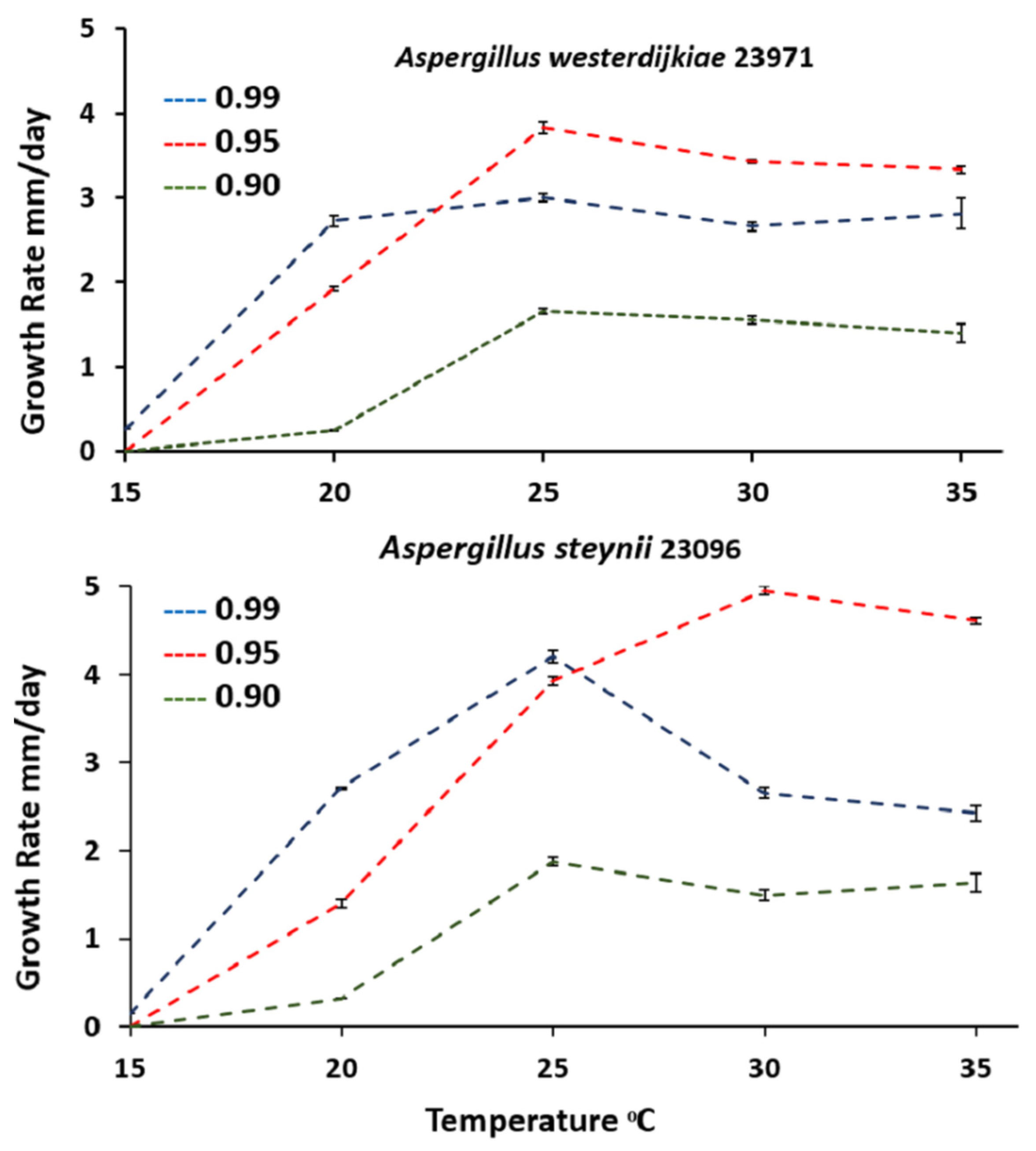
3.1. Growth Assessment in Relation to Abiotic Climatic Variables
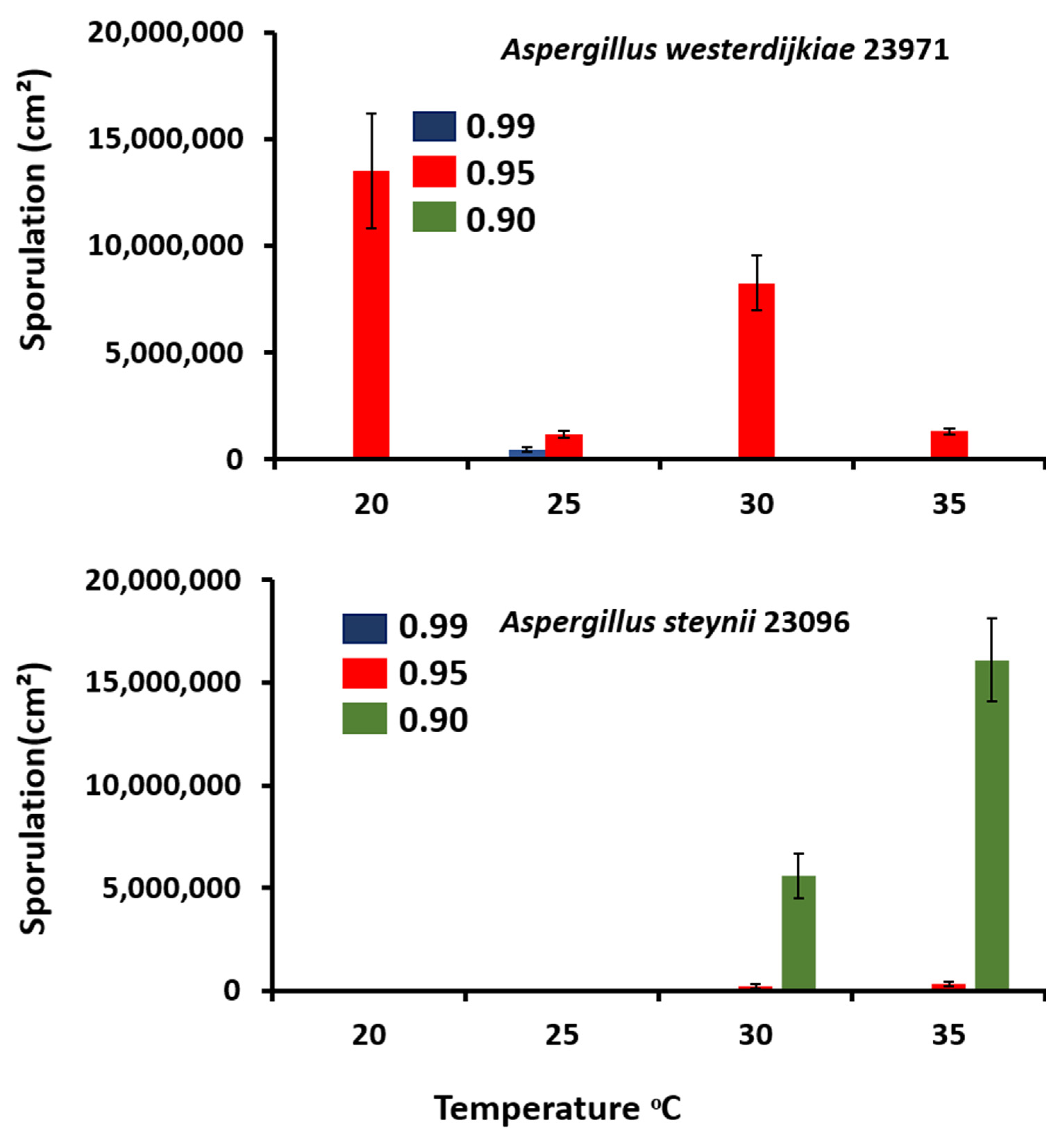
3.2. Sporulation Assessment in Relation to Abiotic Climatic Variables
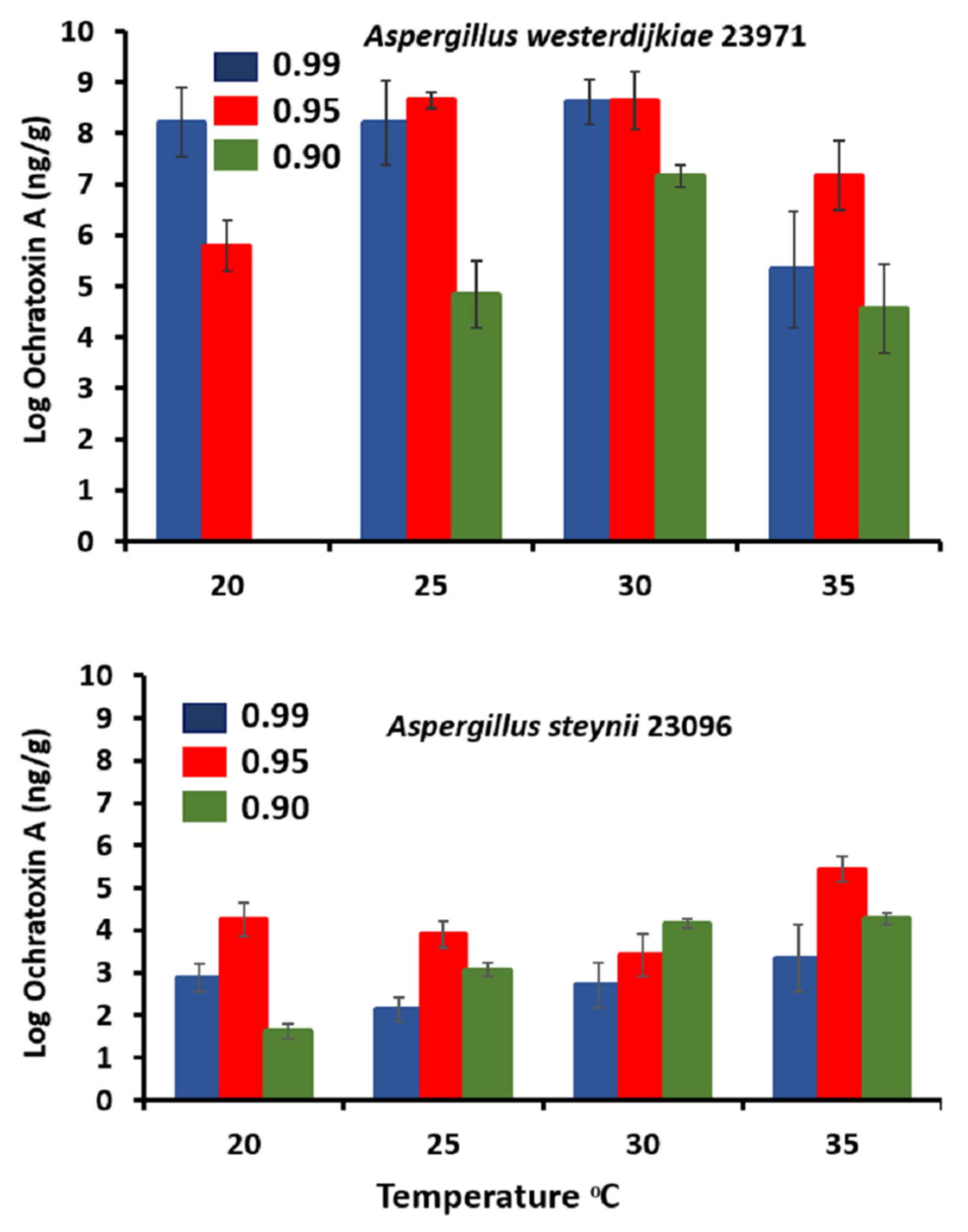
3.3. Ochratoxin A Assessment in Relation to Abiotic Climatic Variables
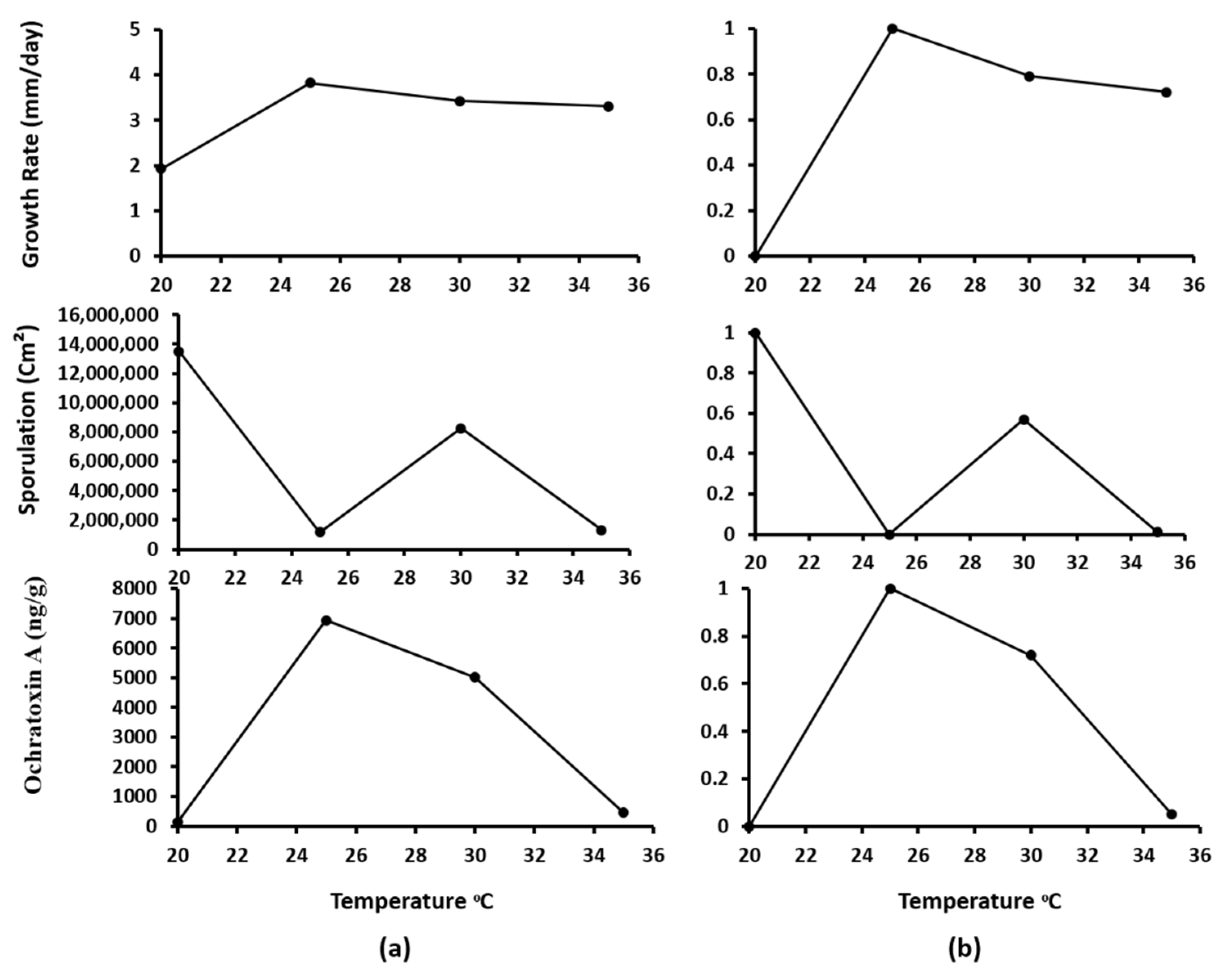
3.4. Predictive Modeling and Validation
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Duarte, S.C.; Pena, A.; Lino, C.M. A review on ochratoxin A occurrence and effects of processing of cereal and cereal derived food products. Food Microbiol. 2010, 27, 187–198. [Google Scholar] [CrossRef]
- Pietri, A.; Rastelli, S.; Bertuzzi, T. Ochratoxin A and aflatoxins in liquorice products. Toxins 2010, 2, 758–770. [Google Scholar] [CrossRef]
- Zaied, C.; Abid, S.; Bouaziz, C.; Chouchane, S.; Jomaa, M.; Bacha, H. Ochratoxin A levels in spices and dried nuts consumed in Tunisia. Food Addit. Contam. Part B 2010, 3, 52–57. [Google Scholar] [CrossRef]
- Frisvad, J.C.; Frank, J.M.; Houbraken, J.A.M.P.; Kuijpers, A.F.A.; Samson, R.A. New ochratoxin A producing species of Aspergillus section circumdati. Stud. Mycol. 2004, 50, 23–43. [Google Scholar]
- Gil-Serna, J.; Vazquez, C.; Sandino, F.G.; Valle, A.M.; Gonzalez-Jáen, M.T.; Patiño, B. Evaluation of growth and ochratoxin A production by Aspergillus steynii and Aspergillus westerdijkiae in green-coffee based medium under different environmental conditions. Food Res. Int. 2014, 61, 127–131. [Google Scholar] [CrossRef] [Green Version]
- Taniwaki, M.H.; Teixeira, A.A.; Teixeira, A.R.; Copetti, M.V.; Iamanaka, B.T. Ochratoxigenic fungi and ochratoxin A in defective coffee beans. Food Res. Int. 2014, 61, 161–166. [Google Scholar] [CrossRef]
- Leitao, A.L. Occurrence of ochratoxin A in coffee: Threads and solutions—A mini-review. Beverages 2019, 5, 36. [Google Scholar] [CrossRef] [Green Version]
- Díaz, G.A.; Torres, R.; Vega, M.; Latorre, B.A. Ochratoxigenic Aspergillus species on grapes from Chilean vineyards and Aspergillus threshold levels on grapes. Int. J. Food Microbiol. 2009, 133, 195–199. [Google Scholar] [CrossRef]
- Santos, L.; Marín, S.; Mateo, E.M.; Gil-Serna, J.; Valle-Algarra, F.M.; Patiño, B.; Ramos, A.J. Mycobiota and co-occurrence of mycotoxins in capsicum powder. Int. J. Food Microbiol. 2011, 151, 270–276. [Google Scholar] [CrossRef]
- Mateo, E.M.; Gil-Serna, J.; Patiño, B.; Jiménez, M. Aflatoxins and ochratoxin A in stored barley grain in Spain and impact of PCR-based strategies to assess the occurrence of aflatoxigenic and ochratoxigenic Aspergillus spp. Int. J. Food Microbiol. 2011, 149, 118–126. [Google Scholar] [CrossRef]
- Vipotnik, Z.; Rodriguez, A.; Rodrigues, P. Aspergillus westerdijkiae as a major ochratoxin A risk in dry-cured ham based-media. Int. J. Food Microbiol. 2017, 16, 244–251. [Google Scholar] [CrossRef]
- Gil-Serna, J.; Vázquez, C.; Sardiñas, N.; González-Jaén, M.T.; Patiño, B. Revision of ochratoxin A production capacity by the main species of Aspergillus section circumdati. Aspergillus steynii revealed as the main risk of OTA contamination. Food Control 2011, 22, 343–345. [Google Scholar] [CrossRef]
- Abdel-Hadi, A.; Magan, N. Influence of physiological factors on growth, sporulation and ochratoxin A/B production of the new Aspergillus ochraceus grouping. World Mycotoxin J. 2009, 2, 429–434. [Google Scholar] [CrossRef] [Green Version]
- Gil-Serna, J.; Patiño, B.; Cortes, L.; Gonzalez-Jaen, M.T.; Vazquez, C. Aspergillus steynii and Aspergillus westerdijkiae as potential risk of OTA contamination in food products in warm climates. Food Microbiol. 2015, 46, 168–175. [Google Scholar] [CrossRef] [Green Version]
- Akbar, A. Growth and Ochratoxin A Production by Aspergillus Species in Coffee Beans: Impact of Climate Change and Control Using O3. Ph.D. Thesis, Cranfield University, Bedford, UK, 2015. [Google Scholar]
- Akbar, A.; Medina, A.; Magan, N. Impact of climate change factors on growth and ochratoxin A production by Aspergillus sections Circumdati and Nigri species on coffee. World Mycotoxin J. 2016, 9, 863–874. [Google Scholar] [CrossRef] [Green Version]
- Medina, A.; Akbar, A.; Baazeem, A.; Rodriguez, A.; Magan, N. Climate change, food security and mycotoxins: Do we know enough? Fungal Biol. Rev. 2017, 31, 143–154. [Google Scholar] [CrossRef] [Green Version]
- Akbar, A.; Medina, A.; Magan, N. Resilience of Aspergillus westerdijkiae strains to interacting climate-related abiotic factors: Effects on growth and ochratoxin A production on coffee-based medium and in stored coffee. Microorganisms 2020, 8, 1268. [Google Scholar] [CrossRef]
- Abdelmohsen, S.; Verheecke-Vaessen, C.; Garcia-Cela, E.; Medina, A.; Magan, N. Dynamics of solute/matric stress interactions with climate change abiotic factors on growth, gene expression and ochratoxin A production by Penicillium verrucosum on a wheat-based matrix. Fungal Biol. 2021, 125, 62–68. [Google Scholar] [CrossRef]
- Baazeem, A.; Garcia-Cela, E.; Medina, A.; Magan, N. Interacting abiotic factors affect growth and aflatoxin b1 production profiles of Aspergillus flavus strains on pistachio-based matrices and pistachio nuts. Front. Microbiol. 2021, 11, 624007. [Google Scholar] [CrossRef]
- Dantigny, P.; Guilmart, A.; Radoi, F.; Bensoussan, M.; Zwietering, M. Modelling the effect of ethanol on growth rate of food spoilage moulds. Int. J. Food Microbiol. 2005, 98, 261–269. [Google Scholar] [CrossRef]
- Garcia, D.; Ramos, A.J.; Sanchis, V.; Marin, S. Predicting mycotoxins in foods: A review. Food Microbiol. 2009, 26, 757–769. [Google Scholar] [CrossRef]
- Whiting, R.C. Microbial modelling in foods. Crit. Rev. Food Sci. Nut. 1995, 6, 467–494. [Google Scholar] [CrossRef]
- Marks, B.P. Status of microbial modelling in food process models. Comp. Rev. Food Sci. Food Saf. 2007, 7, 137–143. [Google Scholar] [CrossRef]
- Kapetanakou, A.E.; Ampavi, A.; Yanniotis, S.; Drosinos, E.H.; Skandamis, P.N. Development of a model describing the effect of temperature, water activity and (gel) structure on growth and ochratoxin A production by Aspergillus carbonarius in vitro and evaluation in food matrices of different viscosity. Food Microbiol. 2011, 28, 727–735. [Google Scholar] [CrossRef]
- Paola, B.; Camardo, L. OTA-grapes: A mechanistic model to predict ochratoxin A risk in grapes, a step beyond the systems approach. Toxins 2015, 7, 3012–3029. [Google Scholar] [CrossRef] [Green Version]
- Kaminiaris, M.D.; Camardo Leggieri, M.; Tsitsigiannis, D.I.; Battilani, P. Afla-pistachio: Development of a mechanistic model to predict the aflatoxin contamination of pistachio nuts. Toxins 2020, 12, 445. [Google Scholar] [CrossRef]
- Gil-Serna, J.; Vazquez, C.; Patiño, B. The genomic regions that contain ochratoxin A biosynthetic genes widely differ in Aspergillus section Circumdati species. Toxins 2020, 12, 754. [Google Scholar] [CrossRef]
- Marin, S.; Sanchis, V.; Magan, N. Water activity, temperature, and pH effects on growth of Fusarium moniliforme and Fusarium proliferatum isolates from maize. Can. J. Microbiol. 1995, 41, 1063–1070. [Google Scholar] [CrossRef]
- Aldred, D.; Magan, N.; Lane, B.S. Influence of water activity and nutrient on growth and production of squalestatin S1 by a Phoma sp. J. Appl. Microbiol. 1999, 87, 842–848. [Google Scholar] [CrossRef]
- Patriarca, A.; Vaamonde, G.; Fernandez, V.; Comerio, R. Influence of water activity and temperature on the growth of Wallemia sebi: Application of a predictive model. Int. J. Food Microbiol. 2001, 68, 61–67. [Google Scholar] [CrossRef]
- Levine, D.M.; Stephan, D.; Szabat, K.A. Statistics for Managers Using Microsoft Excel, 8th ed.; 2017; Available online: http://elibrary.gci.edu.np/handle/123456789/813 (accessed on 6 April 2021).
- Motulsky, H. Essential Biostatistics: A Nonmathematical Approach; Oxford University Press: New York, NY, USA, 2016. [Google Scholar]
- Ross, T. Indices for performance evaluation of predictive models in food microbiology. J. Appl. Microbiol. 1996, 81, 501–508. [Google Scholar] [CrossRef]
- Wawrzyniak, J.; Ryniecki, A.; Gawrysiak-Witulska, M. Kinetics of mould growth in the stored barley ecosystem contaminated with Aspergillus westerdijkiae, Penicillium viridicatum and Fusarium poae at 23–30 °C. J. Sci. Food Agric. 2013, 93, 895–901. [Google Scholar] [CrossRef]
- European Commission. Adapting to Climate Change in Europe—Options for EU Action; Green paper from the Commission to the Council, the European Parliament, the European Economic and Social Committee of the Regions, SEC 849, COM 354 Final; European Commission: Brussels, Belgium, 2007. [Google Scholar]
- Parra, R.; Aldred, D.; Archer, D.; Magan, N. Water activity, solute and temperature modify growth and spore production of wild type and genetically engineered Aspergillus niger strains. Enzyme Microb. Technol. 2004, 35, 232–237. [Google Scholar] [CrossRef] [Green Version]
- Kapetanakou, A.E.; Panagou, E.Z.; Gialitaki, M.; Drosinos, E.H.; Skandamis, P.N. Evaluating the combined effect of water activity, pH and temperature on ochratoxin A production by Aspergillus ochraceus and Aspergillus carbonarius on culture medium and corinth raisins. Food Control 2009, 20, 725–732. [Google Scholar] [CrossRef]
- Rossi, V.; Scandolara, A.; Battilani, P. Effect of environmental conditions on spore production by Fusarium verticillioides, the causal agent of maize ear rot. Europ. J. Plant Pathol. 2009, 123, 159–169. [Google Scholar] [CrossRef]
- Giorni, P.; Camardo Leggieri, M.; Magan, N.; Battilani, P. Comparison of ecological needs for sporulation of Aspergillus flavus sclerotia on natural and artificial substrates. Fungal Biol. 2012, 116, 637–642. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Guo, Z.; Zhong, H.; Wang, S.; Yang, W.; Liu, Y.; Wang, S. RNA-Seq-based transcriptome analysis of aflatoxigenic Aspergillus flavus in response to water activity. Toxins 2014, 6, 3187–3207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lappa, I.K.; Kizis, D.; Panagou, E.Z. Monitoring the temporal expression of genes involved in ochratoxin A production of Aspergillus carbonarius under the influence of temperature and water activity. Toxins 2017, 9, 296. [Google Scholar] [CrossRef] [Green Version]
- Cervini, C.; Gallo, A.; Piemontese, L.; Magistà, D.; Logrieco, A.F.; Ferrara, M.; Solfrizzo, M.; Perrone, G. Effects of temperature and water activity change on ecophysiology of ochratoxigenic Aspergillus carbonarius in field-simulating conditions. Int. J. Food Microbiol. 2020, 315, 108420. [Google Scholar] [CrossRef] [PubMed]
- Perrone, G.; Ferrara, M.; Medina, A.; Pascale, M.; Magan, N. Toxigenic fungi and mycotoxins in a climate change scenario: Ecology, genomics, distribution, prediction and prevention of the risk. Microorganisms 2020, 8, 1496. [Google Scholar] [CrossRef]
- Dantigny, P.; Bensoussan, M. The logarithmic transformation should be avoided for stabilising the variance of mould growth rate. Int. J. Food Microbiol. 2008, 121, 225–228. [Google Scholar] [CrossRef]
- Marin, S.; Ramos, A.J.; Sanchis, V. Modelling Aspergillus flavus growth and aflatoxins production in pistachio nuts. Food Microbiol. 2012, 32, 378–388. [Google Scholar] [CrossRef] [PubMed]
- Marin, S.; Colom, C.; Sanchis, V.; Ramos, A.J. Modelling of growth of aflatoxigenic A. flavus isolates from red chili powder as a function of water availability. Int. J. Food Microbiol. 2009, 128, 491–496. [Google Scholar] [CrossRef]
- Medina, A.; Mateo, R.; Valle-Algarra, F.M.; Mateo, E.M.; Misericordia, J. Effect of carbendazim and physicochemical factors on the growth and ochratoxin A production of Aspergillus carbonarius isolated from grapes. Int. J. Food Microbiol. 2007, 119, 230–235. [Google Scholar] [CrossRef] [PubMed]
- Garcia, D.; Ramos, A.J.; Sanchis, V.; Marin, S. Modeling kinetics of aflatoxin production by Aspergillus flavus in maize-based medium and maize grain. Int. J Food Microbiol. 2013, 162, 182–189. [Google Scholar] [CrossRef] [PubMed]
- Brocklehurst, T. Challenge of food and the environment. In Modelling Microbial Responses in Food; McKellar, R.C., Lu, X., Eds.; CRC Press: Boca Raton, FL, USA, 2004; pp. 197–232. [Google Scholar]
- Marín, S.; Hodzic, I.; Ramos, A.J.; Sanchis, V. Predicting the growth/no growth boundary and ochratoxin A production by Aspergillus carbonarius in pistachio nuts. Food Microbiol. 2008, 25, 683–689. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Aspergillus Westerdijkiae 23971 | Aspergillus Steynii 23096 | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Factor | DF | MS | F | P | DF | MS | F | P | |
| Growth rate | Temperature | 4 | 11.2329 | 2693.48 | 0.00 | 4 | 17.0717 | 6257.19 | 0.00 |
| aw | 2 | 10.2443 | 2456.42 | 0.00 | 2 | 14.5386 | 5328.76 | 0.00 | |
| Temperature × aw | 8 | 0.9489 | 225.61 | 0.00 | 8 | 2.7519 | 1008.65 | 0.00 | |
| Sporulation | Temperature | 3 | 3.420 × 1013 | 3.0773 | 0.046 | 3 | 5.988 × 1013 | 137.093 | 0.00 |
| aw | 2 | 1.438 × 1014 | 12.9402 | 0.00 | 2 | 1.145 × 1014 | 262.299 | 0.00 | |
| Temperature × aw | 6 | 3.632 × 1013 | 3.26787 | 0.017 | 6 | 5.654 × 1013 | 129.375 | 0.00 | |
| Ochratoxin A | Temperature | 3 | 108.844 | 368.25 | 0.00 | 3 | 96.501 | 251.404 | 0.00 |
| aw | 2 | 66.608 | 225.35 | 0.00 | 2 | 70.678 | 182.566 | 0.00 | |
| Temperature × aw | 6 | 24.262 | 82.69 | 0.00 | 6 | 8.064 | 21.008 | 0.00 | |
| Strains | Equation | R2 | |
|---|---|---|---|
| Aspergillus westerdijkiae 23971 | Growth rate | y = 0.0012x3 − 0.1115x2 + 3.3348x − 31.971 | 0.9619 |
| Sporulation | y = −0.0025x3 + 0.2075x2 − 5.7618x + 53.039 | 0.8819 | |
| Ochratoxin A | y = 0.0008x3 − 0.0831x2 + 2.6896x − 27.112 | 0.977 | |
| Aspergillus westerdijkiae 10738 | Growth rate | y = 0.0001x3 − 0.0227x2 + 0.954x − 11.222 | 0.9927 |
| Sporulation | y = −0.0002x3 + 0.0042x2 + 0.3017x + 5.9932 | 0.9871 | |
| Ochratoxin A | y = −0.0004x3 + 0.0153x2 + 0.0258x − 3.7228 | 0.9848 | |
| Aspergillus steynii 22339 | Growth rate | y = 0.001x3 − 0.0904x2 + 2.673x −25.514 | 0.9797 |
| Sporulation | y = −0.0009x3 + 0.0654x2 +1.5539x + 112.197 | 0.984 | |
| Ochratoxin A | y = 0.0008x3 − 0.0687x2 + 2.09792x − 20.24 | 0.9883 | |
| Aspergillus steynii 23096 | Growth rate | y = 0.00003x3 − 0.0106x2 + 0.5627x − 7.321 | 0.9967 |
| Sporulation | y = −0.001x3 + 0.0849x2 − 2.2961x + 19.965 | 0.9903 | |
| Ochratoxin A | y = 0.0004x3 − 0.0302x2 + 0.8883x − 8.5945 | 0.9978 | |
| Aspergillus ochraceus 11952 | Growth rate | y = 0.0011x3 − 0.0934x2 + 2.6015x − 23.685 | 0.9734 |
| Sporulation | y = 0.0013x3 − 0.0997x2 − 2.4917x + 20.127 | 0.9805 | |
| Ochratoxin A | y = 0.0009x3 − 0.062x2 + 1.4539x − 11.226 | 0.9844 |
| Strain | Growth Rate | Sporulation | Ochratoxin A | |||
|---|---|---|---|---|---|---|
| MSE | RMSE | MSE | RMSE | MSE | RMSE | |
| Aspergillus steynii 23096 | 0.0015 | 0.039 | 0.0029 | 0.0539 | 0.0083 | 0.0912 |
| Aspergillus westerdijkia 23971 | 0.0273 | 0.1654 | 0.3094 | 0.5562 | 0.1877 | 0.4332 |
| Aspergillus ochraceus 11952 | 0.0527 | 0.2296 | 0.056 | 0.2367 | 0.1516 | 0.3894 |
| Aspergillus steynii 22339 | 0.0058 | 0.0764 | 0.0832 | 0.2885 | 0.045 | 0.2123 |
| Aspergillus westerdijkia 10738 | 0.0087 | 0.0934 | 0.2217 | 0.4708 | 0.1138 | 0.3373 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdel-Hadi, A.; Alshehri, B.; Waly, M.; Aboamer, M.; Banawas, S.; Alaidarous, M.; Palanisamy, M.; Awad, M.; Baazeem, A. Predictive Modeling and Validation on Growth, Production of Asexual Spores and Ochratoxin A of Aspergillus Ochraceus Group under Abiotic Climatic Variables. Microorganisms 2021, 9, 1321. https://doi.org/10.3390/microorganisms9061321
Abdel-Hadi A, Alshehri B, Waly M, Aboamer M, Banawas S, Alaidarous M, Palanisamy M, Awad M, Baazeem A. Predictive Modeling and Validation on Growth, Production of Asexual Spores and Ochratoxin A of Aspergillus Ochraceus Group under Abiotic Climatic Variables. Microorganisms. 2021; 9(6):1321. https://doi.org/10.3390/microorganisms9061321
Chicago/Turabian StyleAbdel-Hadi, Ahmed, Bader Alshehri, Mohammed Waly, Mohammed Aboamer, Saeed Banawas, Mohammed Alaidarous, Manikandan Palanisamy, Mohamed Awad, and Alaa Baazeem. 2021. "Predictive Modeling and Validation on Growth, Production of Asexual Spores and Ochratoxin A of Aspergillus Ochraceus Group under Abiotic Climatic Variables" Microorganisms 9, no. 6: 1321. https://doi.org/10.3390/microorganisms9061321