
Characterisation of the Antibiotic Profile of Lysobacter capsici AZ78, an Effective Biological Control Agent of Plant Pathogenic Microorganisms

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Maintenance of Microorganisms
2.2. Matrix-Assisted Laser Desorption/Ionisation-Orthogonal Time of Flight Mass Spectrometric Imaging (MALDI-qTOF-MSI)
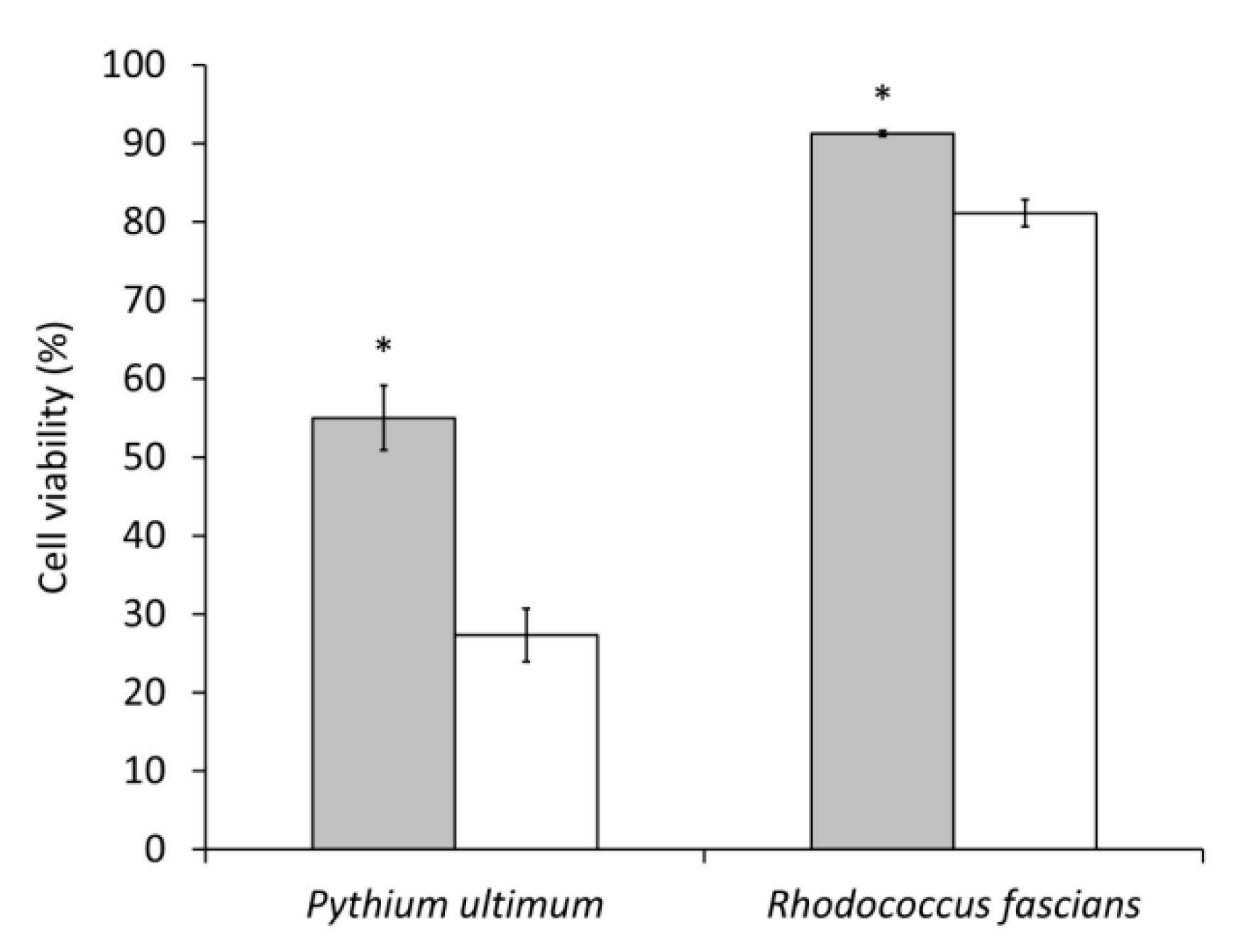
2.3. Evaluation of Lysobacter capsici AZ78 Cell-Free Extracts against Pythium ultimum and Rhodococcus fascians
2.4. LC-HRMS/MS Analysis of Lysobacter capsici AZ78 Cell-Free Extracts
2.4.1. Sample Preparation
2.4.2. LC-HRMS/MS Measurement
2.4.3. LC-HRMS/MS Data Analysis
2.5. Impact of High-Temperature Exposure on Lysobacter capsici AZ78 Cell-Free Supernatant
2.6. Statistical Analyses
3. Results
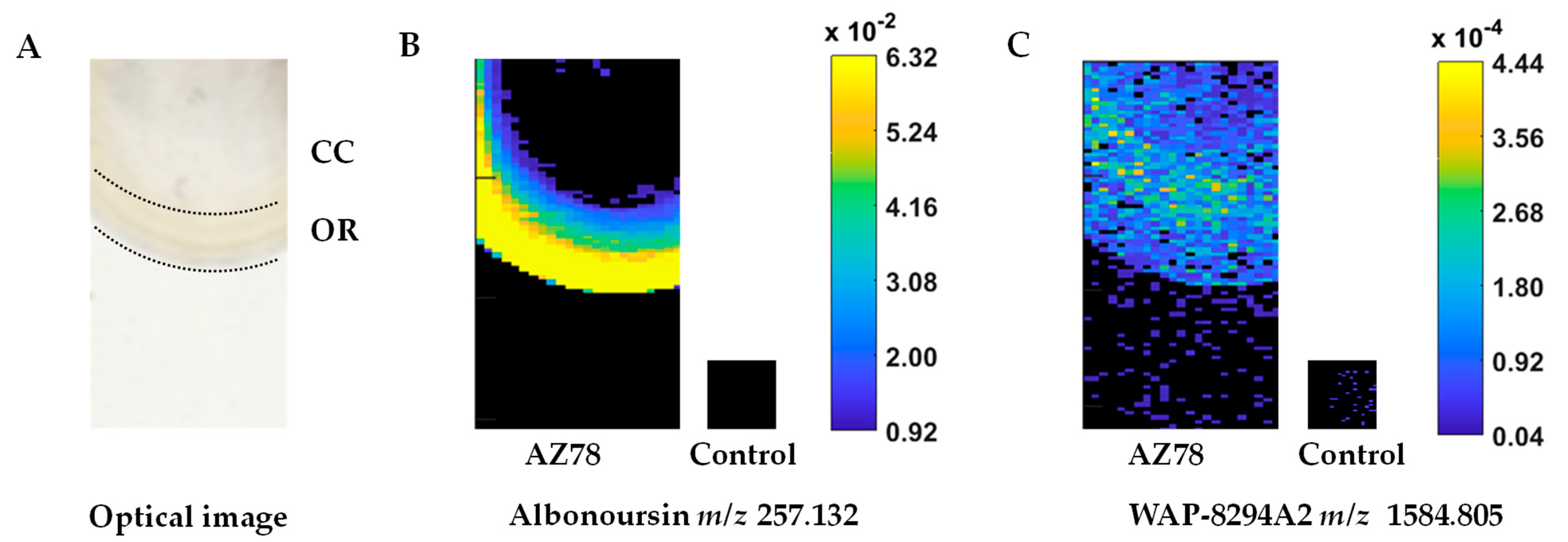
3.1. Mass Spectrometric Imaging Reveals a High Diversity of Antimicrobial Analytes in Lysobacter capsici AZ78 Metabolic Profile and an Intense Metabolic Activity in the Macrocolony Outer Ring
3.2. Lysobacter capsici AZ78 Produces Secondary Metabolites with Antioomycete and Antibacterial Activity
3.3. The Cyclic Lipodepsipeptide WAP-8294A2, the Polycyclic Tetramic Acid Macrolactam Dihydromaltophilin and Four Diketopiperazines Are Confirmed as Antimicrobial Constituents of Lysobacter capsici AZ78 Metabolome
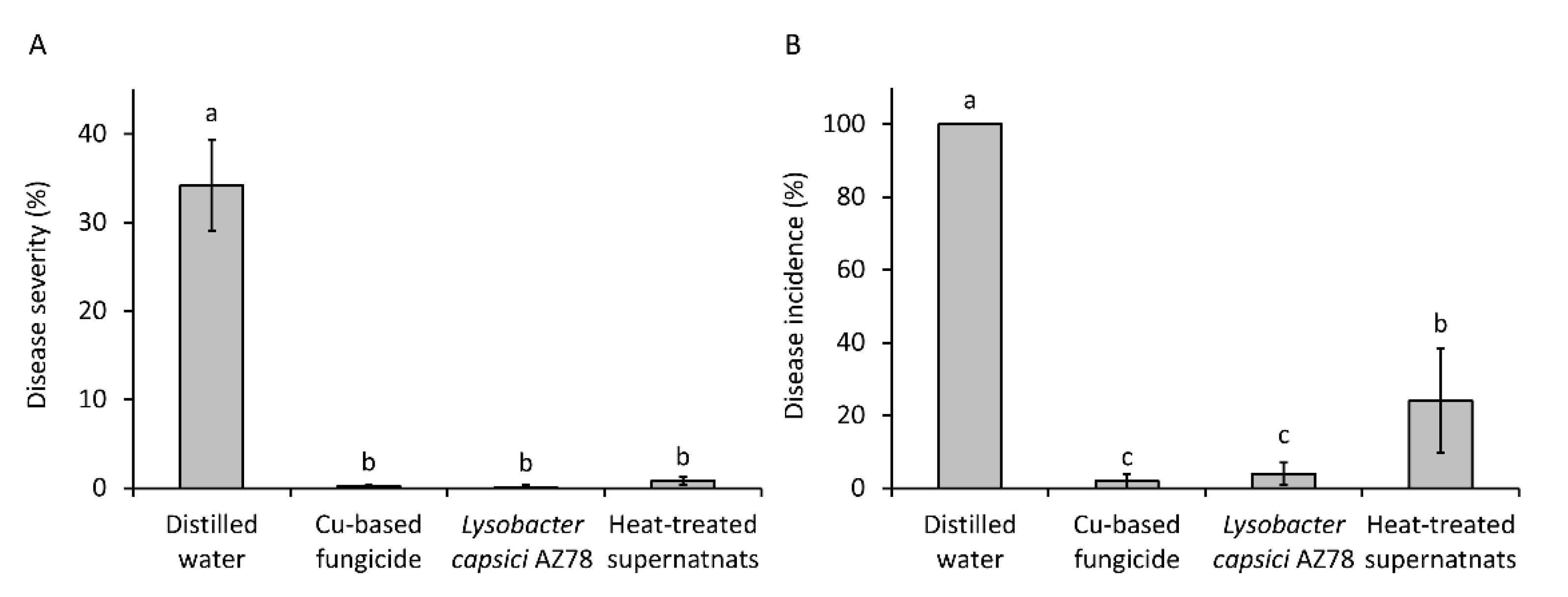
3.4. Lysobacter capsici AZ78 Produces Dihydromaltophilin Resistant to Heat Shock and Active against Plasmopara viticola on Grapevine Leaf Disk
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Agrios, G.N. Plant Pathology, 5th ed.; Academic: Oxford, UK, 2005; ISBN 9780120445653. [Google Scholar]
- Colla, P.; Gilardi, G.; Gullino, M.L. A review and critical analysis of the European situation of soilborne disease management in the vegetable sector. Phytoparasitica 2012, 40, 515–523. [Google Scholar] [CrossRef]
- Köhl, J.; Kolnaar, R.; Ravensberg, W.J. Mode of action of microbial biological control agents against plant diseases: Relevance beyond efficacy. Front. Plant Sci. 2019, 10, 845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Commission Regulation (EU). No 546/2011 of 10 June 2011 implementing Regulation (EC) No 1107/2009 of the European Parliament and of the Council as regards uniform principles for evaluation and authorisation of plant protection products. Off. J. Eur. Union L 2009, 155, 127–175. [Google Scholar]
- Commission Regulation (EU). No 283/2013 of 1 March 2013 setting out the data requirements for active sub-stances, in accordance with Regulation (EC) No 1107/2009 of the European Parliament and of the Council concerning the placing of plant protection products on the market. Off. J. Eur. Union L 2013, 93, 1–84. [Google Scholar]
- Commission Regulation (EU). No 284/2013 of 1 March 2013 setting out the data requirements for plant pro-tec-tion products, in accordance with Regulation (EC) No 1107/2009 of the European Parliament and of the Council concerning the placing of plant protection products on the market. Off. J. Eur. Union L 2013, 93, 85–152. [Google Scholar]
- Jochum, C.; Osborne, L.; Yuen, G. Fusarium head blight biological control with Lysobacter enzymogenes strain C3. Biol. Control. 2006, 39, 336–344. [Google Scholar] [CrossRef]
- Ji, G.-H.; Wei, L.-F.; He, Y.-Q.; Wu, Y.-P.; Bai, X.-H. Biological control of rice bacterial blight by Lysobacter antibioticus strain 13-1. Biol. Control. 2008, 45, 288–296. [Google Scholar] [CrossRef]
- Puopolo, G.; Raio, A.; Zoina, A. Identification and characterization of Lysobacter capsici strain PG4: A new plant health-promoting rhizobacterium. J. Plant Pathol. 2010, 92, 157–164. [Google Scholar] [CrossRef]
- Puopolo, G.; Tomada, S.; Pertot, I. The impact of the omics era on the knowledge and use of Lysobacter species to control phytopathogenic microorganisms. J. Appl. Microbiol. 2018, 124, 15–27. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Qian, G.; Ye, Y.; Wright, S.; Chen, H.; Shen, Y.; Liu, F.; Du, L. Heterocyclic aromaticn-oxidation in the biosynthesis of phenazine antibiotics from Lysobacter antibioticus. Org. Lett. 2016, 18, 2495–2498. [Google Scholar] [CrossRef]
- Panthee, S.; Hamamoto, H.; Paudel, A.; Sekimizu, K. Lysobacter species: A potential source of novel antibiotics. Arch. Microbiol. 2016, 198, 839–845. [Google Scholar] [CrossRef]
- Meyers, D.; Cooper, R.; Dean, L.; Johnson, J.H.; Slusarchyk, D.S.; Trejo, W.H.; Singh, P.D. Catacandins, novel anticandidal antibiotics of bacterial origin. J. Antibiot. (Tokyo) 1985, 38, 1642–1648. [Google Scholar] [CrossRef] [Green Version]
- Nakayama, T.; Homma, Y.; Hashidoko, Y.; Mizutani, J.; Tahara, S. Possible role of xanthobaccins produced by Stenotrophomonas sp. strain SB-K88 in suppression of sugar beet damping-off disease. Appl. Environ. Microbiol. 1999, 65, 4334–4339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, L.; Wu, P.; Wright, S.J.; Du, L.; Wei, X. Bioactive polycyclic tetramate macrolactams from Lysobacter enzymogenes and their absolute configurations by theoretical ECD calculations. J. Nat. Prod. 2015, 78, 1841–1847. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Li, Y.; Li, Z.; Zhang, J.; Lu, C.; Wang, H.; Shen, Y.; Du, L. Alteramide B is a microtubule antagonist of inhibiting Candida albicans. Biochim. Biophys. Acta (BBA) Gen. Subj. 2016, 1860, 2097–2106. [Google Scholar] [CrossRef] [Green Version]
- Yu, F.; Zaleta-Rivera, K.; Zhu, X.; Huffman, J.; Millet, J.C.; Harris, S.D.; Yuen, G.; Li, X.-C.; Du, L. Structure and biosynthesis of heat-stable antifungal factor (HSAF), a broad-spectrum antimycotic with a novel mode of action. Antimicrob. Agents Chemother. 2007, 51, 64–72. [Google Scholar] [CrossRef] [Green Version]
- Puopolo, G.; Giovannini, O.; Pertot, I. Lysobacter capsici AZ78 can be combined with copper to effectively control Plasmopara viticola on grapevine. Microbiol. Res. 2014, 169, 633–642. [Google Scholar] [CrossRef]
- Puopolo, G.; Cimmino, A.; Palmieri, M.; Giovannini, O.; Evidente, A.; Pertot, I. Lysobacter capsici AZ78 produces cyclo(l -Pro-l -Tyr), a 2,5-diketopiperazine with toxic activity against sporangia of Phytophthora infestans and Plasmopara viticola. J. Appl. Microbiol. 2014, 117, 1168–1180. [Google Scholar] [CrossRef]
- Puopolo, G.; Tomada, S.; Sonego, P.; Moretto, M.; Engelen, K.; Perazzolli, M.; Pertot, I. The Lysobacter capsici AZ78 genome has a gene pool enabling it to interact successfully with phytopathogenic microorganisms and environmental factors. Front. Microbiol. 2016, 7, 96. [Google Scholar] [CrossRef] [Green Version]
- Puopolo, G.; Pertot, I. 2014. A New Bacterial Lysobacter Capsici Strain and Uses Thereof. WO/2014/173906, PCT/EP2014/058151 U.S. Patent No. 9,756,863 Patent and Trademark Office: Washington, DC, USA. 12 September 2017. [Google Scholar]
- Tomada, S.; Sonego, P.; Moretto, M.; Engelen, K.; Pertot, I.; Perazzolli, M.; Puopolo, G. Dual RNA-Seq of Lysobacter capsici AZ78-Phytophthora infestans interaction shows the implementation of attack strategies by the bacterium and unsuccessful oomycete defense responses. Environ. Microbiol. 2017, 19, 4113–4125. [Google Scholar] [CrossRef] [PubMed]
- Cimmino, A.; Bejarano, A.; Masi, M.; Puopolo, G.; Evidente, A. Isolation of 2,5-diketopiperazines from Lysobacter capsici AZ78 with activity against Rhodococcus fascians. Nat. Prod. Res. 2020, 1–9. [Google Scholar] [CrossRef]
- Brescia, F.; Marchetti-Deschmann, M.; Musetti, R.; Perazzolli, M.; Pertot, I.; Puopolo, G. The rhizosphere signature on the cell motility, biofilm formation and secondary metabolite production of a plant-associated Lysobacter strain. Microbiol. Res. 2020, 234, 126424. [Google Scholar] [CrossRef]
- Vlassi, A.; Nesler, A.; Parich, A.; Puopolo, G.; Schuhmacher, R. Volatile-mediated inhibitory activity of rhizobacteria as a result of multiple factors interaction: The case of Lysobacter capsici AZ78. Microorganisms 2020, 8, 1761. [Google Scholar] [CrossRef] [PubMed]
- Vlassi, A.; Nesler, A.; Perazzolli, M.; Lazazzara, V.; Büschl, C.; Parich, A.; Puopolo, G.; Schuhmacher, R. Volatile organic compounds from Lysobacter capsici AZ78 as potential candidates for biological control of soilborne plant pathogens. Front. Microbiol. 2020, 11, 1748. [Google Scholar] [CrossRef] [PubMed]
- Holzlechner, M.; Reitschmidt, S.; Gruber, S.; Zeilinger, S.; Marchetti-Deschmann, M. Visualizing fungal metabolites during mycoparasitic interaction by MALDI mass spectrometry imaging. Proteomics 2016, 16, 1742–1746. [Google Scholar] [CrossRef] [Green Version]
- Klinkert, I.; Chughtai, K.; Ellis, S.R.; Heeren, R. Methods for full resolution data exploration and visualization for large 2D and 3D mass spectrometry imaging datasets. Int. J. Mass Spectrom. 2014, 362, 40–47. [Google Scholar] [CrossRef]
- Robichaud, G.; Garrard, K.P.; Barry, J.A.; Muddiman, D.C. Msireader: An open-source interface to view and analyze high resolving power MS imaging files on matlab platform. J. Am. Soc. Mass Spectrom. 2013, 24, 718–721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sud, M.; Fahy, E.; Cotter, D.; Azam, K.; Vadivelu, I.; Burant, C.; Edison, A.; Fiehn, O.; Higashi, R.; Nair, K.S.; et al. Metabolomics Workbench: An international repository for metabolomics data and metadata, metabolite standards, protocols, tutorials and training, and analysis tools. Nucleic Acids Res. 2016, 44, D463–D470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pluskal, T.; Castillo, S.; Villar-Briones, A.; Orešič, M. MZmine 2: Modular framework for processing, visualizing, and analyzing mass spectrometry-based molecular profile data. BMC Bioinform. 2010, 11, 395. [Google Scholar] [CrossRef] [Green Version]
- Smith, C.A.; Want, E.J.; O’Maille, G.; Abagyan, R.; Siuzdak, G. XCMS: Processing mass spectrometry data for metabolite profiling using nonlinear peak alignment, matching, and identification. Anal. Chem. 2006, 78, 779–787. [Google Scholar] [CrossRef]
- Dodder, N. OrgMassSpecR: Organic Mass Spectrometry. R Package Version 0.5-3. 2017. Available online: https://CRAN.R-project.org/package=OrgMassSpecR (accessed on 15 June 2021).
- Sumner, L.W.; Lei, Z.; Nikolau, B.J.; Saito, K.; Roessner, U.; Trengove, R. Proposed quantitative and alphanumeric metabolite identification metrics. Metabolomics 2014, 10, 1047–1049. [Google Scholar] [CrossRef]
- Bofinger, M.R.; De Sousa, L.S.; Fontes, J.E.N.; Marsaioli, A.J. Diketopiperazines as cross-communication quorum-sensing signals between Cronobacter sakazakii and Bacillus cereus. ACS Omega 2017, 2, 1003–1008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giebel, R.; Worden, C.; Rust, S.; Kleinheinz, G.; Robbins, M.; Sandrin, T. Microbial Fingerprinting using Matrix-Assisted Laser Desorption Ionization Time-Of-Flight Mass Spectrometry (MALDI-TOF MS); Elsevier B.V.: Amsterdam, The Netherlands, 2010; Volume 71, pp. 149–184. ISBN 9780123809933. [Google Scholar]
- Molina-Santiago, C.; Pearson, J.R.; Navarro, Y.; Berlanga-Clavero, M.V.; Caraballo-Rodriguez, A.M.; Petras, D.; García-Martín, M.L.; Lamon, G.; Haberstein, B.; Cazorla, F.M.; et al. The extracellular matrix protects Bacillus subtilis colonies from Pseudomonas invasion and modulates plant co-colonization. Nat. Commun. 2019, 10, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziegler, D.; Mariotti, A.; Pflüger, V.; Saad, M.; Vogel, G.; Tonolla, M.; Perret, X. In situ identification of plant-invasive bacteria with MALDI-TOF mass spectrometry. PLoS ONE 2012, 7, e37189. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, L.; Sánchez-Juanes, F.; García-Fraile, P.; Rivas, R.; Mateos, P.F.; Martínez-Molina, E.; González-Buitrago, J.M.; Velazquez, E. MALDI-TOF mass spectrometry is a fast and reliable platform for identification and ecological studies of species from family Rhizobiaceae. PLoS ONE 2011, 6, e20223. [Google Scholar] [CrossRef] [Green Version]
- Itoh, H.; Tokumoto, K.; Kaji, T.; Paudel, A.; Panthee, S.; Hamamoto, H.; Sekimizu, K.; Inoue, M. Total synthesis and biological mode of action of WAP-8294A2: A menaquinone-targeting antibiotic. J. Org. Chem. 2017, 83, 6924–6935. [Google Scholar] [CrossRef]
- Chen, X.; Li, S.; Yu, L.; Miller, A.; Du, L. Systematic optimization for production of the anti- MRSA antibiotics WAP -8294A in an engineered strain of Lysobacter enzymogenes. Microb. Biotechnol. 2019, 12, 1430–1440. [Google Scholar] [CrossRef] [Green Version]
- Yu, L.; Su, W.; Fey, P.D.; Liu, F.; Du, L. Yield improvement of the Anti-MRSA antibiotics WAP-8294A by CRISPR/dCas9 combined with refactoring self-protection genes in Lysobacter enzymogenes OH11. ACS Synth. Biol. 2018, 7, 258–266. [Google Scholar] [CrossRef] [PubMed]
- Gustafsson, J.O.; Eddes, J.S.; Meding, S.; Koudelka, T.; Oehler, M.K.; McColl, S.R.; Hoffmann, P. Internal calibrants allow high accuracy peptide matching between MALDI imaging MS and LC-MS/MS. J. Proteom. 2012, 75, 5093–5105. [Google Scholar] [CrossRef]
- Han-Jen, R.E.; Wai-Fong, Y.; Kok-Gan, C. Pandoraea sp. Rb-44, a novel quorum sensing soil bacterium. Sensors 2013, 13, 14121–14132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quezada, M.; Licona-Cassani, C.; Cruz-Morales, P.; Salim, A.A.; Marcellin, E.; Capon, R.J.; Barona-Gómez, F. Diverse cone-snail species harbor closely related Streptomyces species with conserved chemical and genetic profiles, including polycyclic tetramic acid macrolactams. Front. Microbiol. 2017, 8, 2305. [Google Scholar] [CrossRef] [PubMed]
- Jakobi, M.; Winkelmann, G.; Kaiser, D.; Kempter, C.; Jung, G.; Berg, G.; Bahl, H. Maltophilin: A new antifungal compound produced by Stenotrophomonas maltophilia R3089. J. Antibiot. 1996, 49, 1101–1104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shigemori, H.; Bae, M.A.; Yazawa, K.; Sasaki, T.; Kobayashi, J. Alteramide A, a new tetracyclic alkaloid from a bacterium Alteromonas sp. associated with the marine sponge Halichondria okadai. J. Org. Chem. 1992, 57, 4317–4320. [Google Scholar] [CrossRef]
- Lou, L.; Qian, G.; Du, L.; Xie, Y.; Hang, J.; Chen, H.; Zaleta-Rivera, K.; Li, Y.; Shen, Y.; Dussault, P.H.; et al. Biosynthesis of HSAF, a tetramic acid-containing macrolactam from Lysobacter enzymogenes. J. Am. Chem. Soc. 2010, 133, 643–645. [Google Scholar] [CrossRef] [Green Version]
- Kato, A.; Hirata, H.; Ohashi, Y.; Fujii, K.; Mori, K.; Harada, K.-I. A new anti-MRSA antibiotic complex, WAP-8294A II. Structure characterization of minor components by ESI LCMS and MS/MS. J. Antibiot. (Tokyo) 2011, 64, 373–379. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.; Tian, L.; Po, K.H.L.; Chen, S.; Li, X. Total synthesis and a systematic structure-activity relationship study of WAP-8294A2. Bioorganic Med. Chem. 2020, 28, 115677. [Google Scholar] [CrossRef] [PubMed]
- Kalaiyarasi, M.; Ahmad, P.; Vijayaraghavan, P. Enhanced production antibiotics using green gram husk medium by Streptomyces sp. SD1 using response surface methodology. J. King Saud Univ. Sci. 2020, 32, 2134–2141. [Google Scholar] [CrossRef]
- Tang, B.; Laborda, P.; Sun, C.; Xu, G.; Zhao, Y.; Liu, F. Improving the production of a novel antifungal alteramide B in Lysobacter enzymogenes OH11 by strengthening metabolic flux and precursor supply. Bioresour. Technol. 2019, 273, 196–202. [Google Scholar] [CrossRef]
- Zhao, Y.; Cheng, C.; Jiang, T.; Xu, H.; Chen, Y.; Ma, Z.; Qian, G.; Liu, F. Control of wheat Fusarium head blight by heat-stable antifungal factor (HSAF) from Lysobacter enzymogenes. Plant Dis. 2019, 103, 1286–1292. [Google Scholar] [CrossRef]
- Li, C.; Tang, B.; Cao, S.; Bao, Y.; Sun, W.; Zhao, Y.; Liu, F. Biocontrol ability and action mechanism of dihydromaltophilin against Colletotrichum fructicola causing anthracnose of pear fruit. Pest Manag. Sci. 2021, 77, 1061–1069. [Google Scholar] [CrossRef]
- Yue, H.; Jiang, J.; Taylor, A.J.; Leite, A.D.L.; Dodds, E.D.; Du, L. Outer membrane vesicle-mediated codelivery of the antifungal HSAF metabolites and lytic polysaccharide monooxygenase in the predatory Lysobacter enzymogenes. ACS Chem. Biol. 2021. [Google Scholar] [CrossRef]
- Li, S.; Calvo, A.M.; Yuen, G.Y.; Du, L.; Harris, S.D. Induction of cell wall thickening by the antifungal compound dihydromaltophilin disrupts fungal growth and is mediated by sphingolipid biosynthesis. J. Eukaryot. Microbiol. 2009, 56, 182–187. [Google Scholar] [CrossRef]
- Ding, Y.; Li, Z.; Li, Y.; Lu, C.; Wang, H.; Shen, Y.; Du, L. HSAF-induced antifungal effects in Candida albicans through ROS-mediated apoptosis. RSC Adv. 2016, 6, 30895–30904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rhee, K.-H. Purification and identification of an antifungal agent from Streptomyces sp. KH-614 antagonistic to rice blast fungus, Pyricularia oryzae. J. Microbiol. Biotechnol. 2003, 13, 984–988. [Google Scholar]
- Fukushima, K.; Yazawa, K.; Arai, T. Biological activities of albonoursin. J. Antibiot. 1973, 26, 175–176. [Google Scholar] [CrossRef] [PubMed]
- Lautru, S.; Gondry, M.; Genet, R.; Pernodet, J.-L. The albonoursin gene cluster of S. noursei: Biosynthesis of diketopiperazine metabolites independent of nonribosomal peptide synthetases. Chem. Biol. 2002, 9, 1355–1364. [Google Scholar] [CrossRef] [Green Version]
- Wattana-Amorn, P.; Charoenwongsa, W.; Williams, C.; Crump, M.P.; Apichaisataienchote, B. Antibacterial activity of cyclo(L-Pro-L-Tyr) and cyclo(D-Pro-L-Tyr) from Streptomyces sp. strain 22-4 against phytopathogenic bacteria. Nat. Prod. Res. 2016, 30, 1980–1983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holden, M.; Chhabra, S.R.; De Nys, R.; Stead, P.; Bainton, N.J.; Hill, P.; Manefield, M.; Kumar, N.; Labatte, M.; England, D.; et al. Quorum-sensing cross talk: Isolation and chemical characterization of cyclic dipeptides from Pseudomonas aeruginosa and other Gram-negative bacteria. Mol. Microbiol. 2002, 33, 1254–1266. [Google Scholar] [CrossRef]
- Degrassi, G.; Aguilar, C.; Bosco, M.; Zahariev, S.; Pongor, S.; Venturi, V. Plant growth-promoting Pseudomonas putida WCS358 produces and secretes four cyclic dipeptides: Cross-talk with quorum sensing bacterial sensors. Curr. Microbiol. 2002, 45, 250–254. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Measured a | Annotated | |||
|---|---|---|---|---|
| m/z | Adduct Ion b | Molecular Formula | Compound Name | Compound Class |
| 257.132 | [M+H]+ | C15H17N2O2 | Albonoursin | 2,5-diketopiperazine |
| 748.433 | [M+H]+ | C38H70NO13 | Clarithromycin | Macrolide |
| 1419.866 | [M+Na]+ | C71H132N2O24Na | Mathemycin A | Macrolactone |
| 1447.770 | [M+H]+ | C75H115O27 | Langkolide | - |
| [M+H]+ | C72H111N12O19 | SNA-60-367-14 | Cyclic depsipeptide | |
| 1584.805 | [M+Na]+ | C73H111N17O21Na | WAP-8294A2 | Cyclic lipodepsipeptide |
| 1598.827 | [M+Na]+ | C74H113N17O21Na | WAP-8294A4/WAP-294Ax13 * | - |
| 1600.81 | [M+K]+ | C73H111N17O21K | WAP-8294A2 | - |
| 1614.838 | [M+K]+ | C74H113N17O21K | WAP-8294A4/WAP-8294Ax13 * | - |
| 1650.784 | [M+Na]+ | C72H109N17O26Na | CB-182333 | Cyclic depsipeptide |
| [M+Na]+ | C72H109N17O26Na | CB-182349 | - | |
| [M+K]+ | C72H109N17O25K | CB-182348 | - | |
| Compound Class | Compound Name | Detected Ion Species 3 |
|---|---|---|
| 2,5-diketopiperazine | Albonoursin 1 | [M+H]+, [M-H2O+H]+ |
| - | Cyclo(Pro-Val) 2 | [M+H]+, [M+Na]+ |
| - | Cyclo(Phe-Pro) 2 | [M+H]+, [M+Na]+ |
| - | Cyclo(Pro-Leu) 2 | [M+H]+, [M+Na]+ |
| - | Cyclo(Pro-Tyr) 2 | [M+H]+, [M+Na]+ |
| Polycyclic tetramic acid | Dihydromaltophilin 2 | [M+H]+, [M-H]−, [M+Na]+− |
| Macrolactam | Maltophilin/Alteramide A 2 * | [M+H]+, [M-H]−, [M+K]+, [M+Na]+ |
| - | Alteramide B 2 | [M+H]+, [M-H]−, [M+Na]+, [M+NH4]+ |
| Cyclic lipodepsipeptide | WAP-8294A2 1 | [M+K]+, [M+Na]+, [M+H]+, [M-H]− |
| - | WAP-8294A4/WAP-8294Ax13 1 * | [M+K]+, [M+Na]+, [M+H]+, [M-H]− |
| Compound | Molecular Formula | m/z Deviation [ppm] a | RT Deviation [%] b | Peak Shape Similarity [PCC] c | MS/MS Similarity [cos Φ] d | Compound Identification Score e |
|---|---|---|---|---|---|---|
| Dihydromaltophilin | C29H40N2O6 | 0.61 | 0.09 | 0.91 | 0.97 | Z| |
| WAP-8294A2 | C73H111N17O21 | 0.87 | 0.44 | 0.91 f | 0.97 | 9.0 |
| Cyclo(Pro-Val) | C10H16N2O2 | 0.36 | 0.16 | 0.99 | 0.99 | 9.0 |
| Cyclo(Phe-Pro) | C14H16N2O2 | 0.77 | 0.23 | 0.98 | 1.00 | 9.0 |
| Cyclo(Pro-Leu) putative isomer at RT 12.0 min | C11H18N2O2 | 0.71 | 1.63 | 0.85 f | 1.00 | 8.0 |
| Cyclo(Pro-Leu) putative isomer at RT 12.5 min | C11H18N2O2 | 0.62 | 2.13 | 0.46 f | 1.00 | 8.0 |
| Cyclo(L-Pro-L-Tyr) | C14H16N2O3 | 0.88 | 0.59 | 0.96 | 0.99 | 8.0 |
| Cyclo(D-Pro-L-Tyr) * | C14H16N2O3 | _ | _ | _ | _ | _ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brescia, F.; Vlassi, A.; Bejarano, A.; Seidl, B.; Marchetti-Deschmann, M.; Schuhmacher, R.; Puopolo, G. Characterisation of the Antibiotic Profile of Lysobacter capsici AZ78, an Effective Biological Control Agent of Plant Pathogenic Microorganisms. Microorganisms 2021, 9, 1320. https://doi.org/10.3390/microorganisms9061320
Brescia F, Vlassi A, Bejarano A, Seidl B, Marchetti-Deschmann M, Schuhmacher R, Puopolo G. Characterisation of the Antibiotic Profile of Lysobacter capsici AZ78, an Effective Biological Control Agent of Plant Pathogenic Microorganisms. Microorganisms. 2021; 9(6):1320. https://doi.org/10.3390/microorganisms9061320
Chicago/Turabian StyleBrescia, Francesca, Anthi Vlassi, Ana Bejarano, Bernard Seidl, Martina Marchetti-Deschmann, Rainer Schuhmacher, and Gerardo Puopolo. 2021. "Characterisation of the Antibiotic Profile of Lysobacter capsici AZ78, an Effective Biological Control Agent of Plant Pathogenic Microorganisms" Microorganisms 9, no. 6: 1320. https://doi.org/10.3390/microorganisms9061320