
In Vitro Selection of Probiotics, Prebiotics, and Antioxidants to Develop an Innovative Synbiotic (NatuREN G) and Testing Its Effect in Reducing Uremic Toxins in Fecal Batches from CKD Patients

 ,
,  , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganisms and Culture Conditions
2.2. Chemical Characterization of Food Matrices
2.3. Evaluation of Effects Exerted by Antioxidants and Prebiotics on Probiotics Growth
2.4. Fecal Media from Healthy Subjects and Chronic Kidney Disease Patients
2.5. Amino Acids Extraction and Detection
2.6. Short Chain Fatty Acids Detection
2.7. In Vitro Effects of the Innovative Synbiotic Formulation on the Fecal Microbiota of HC and CKD Subjects
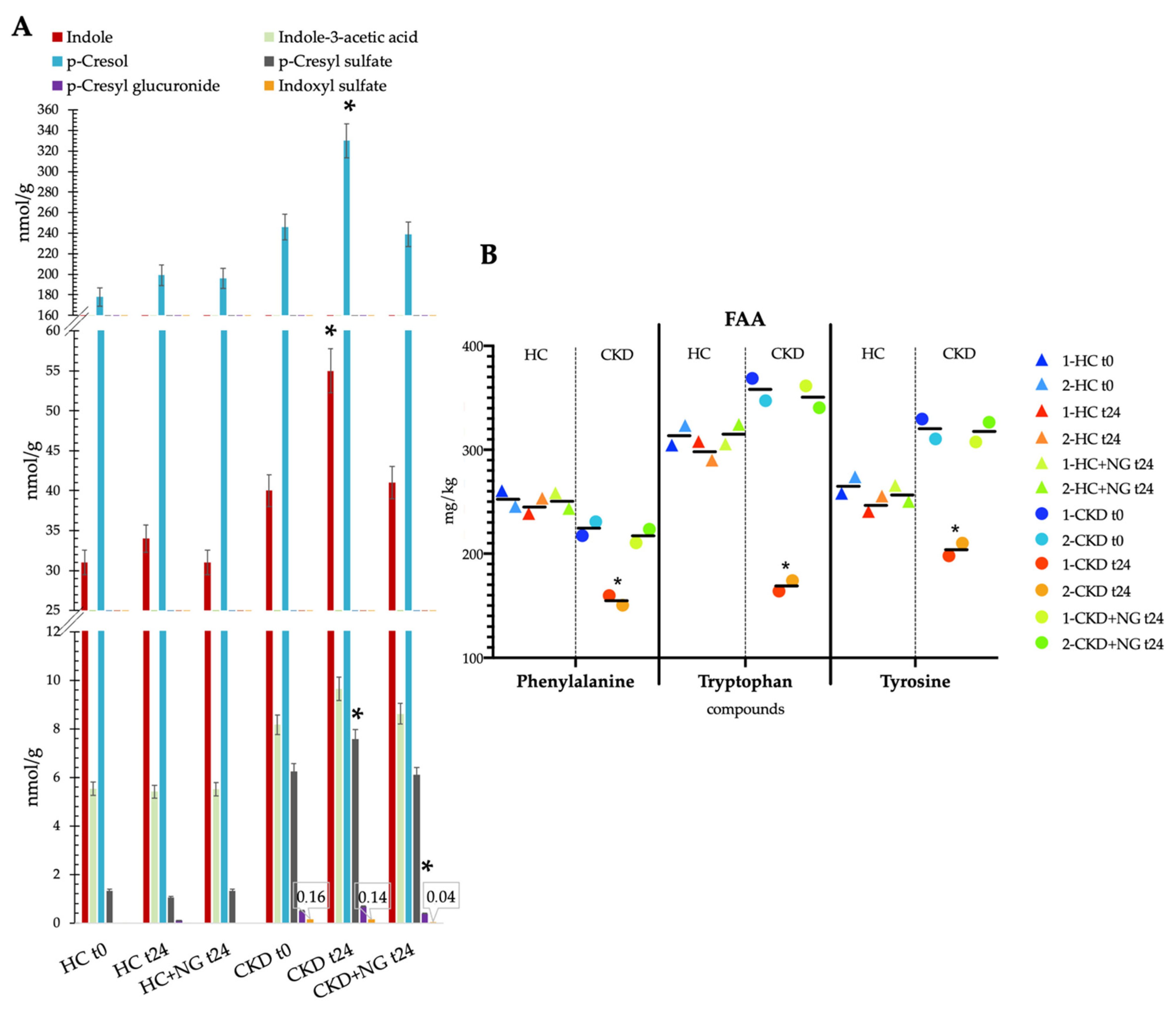
2.8. Uremic Toxins Detection
2.9. Statistical Analyses
3. Results
3.1. Characterization of Food Matrices Rich in Antioxidants
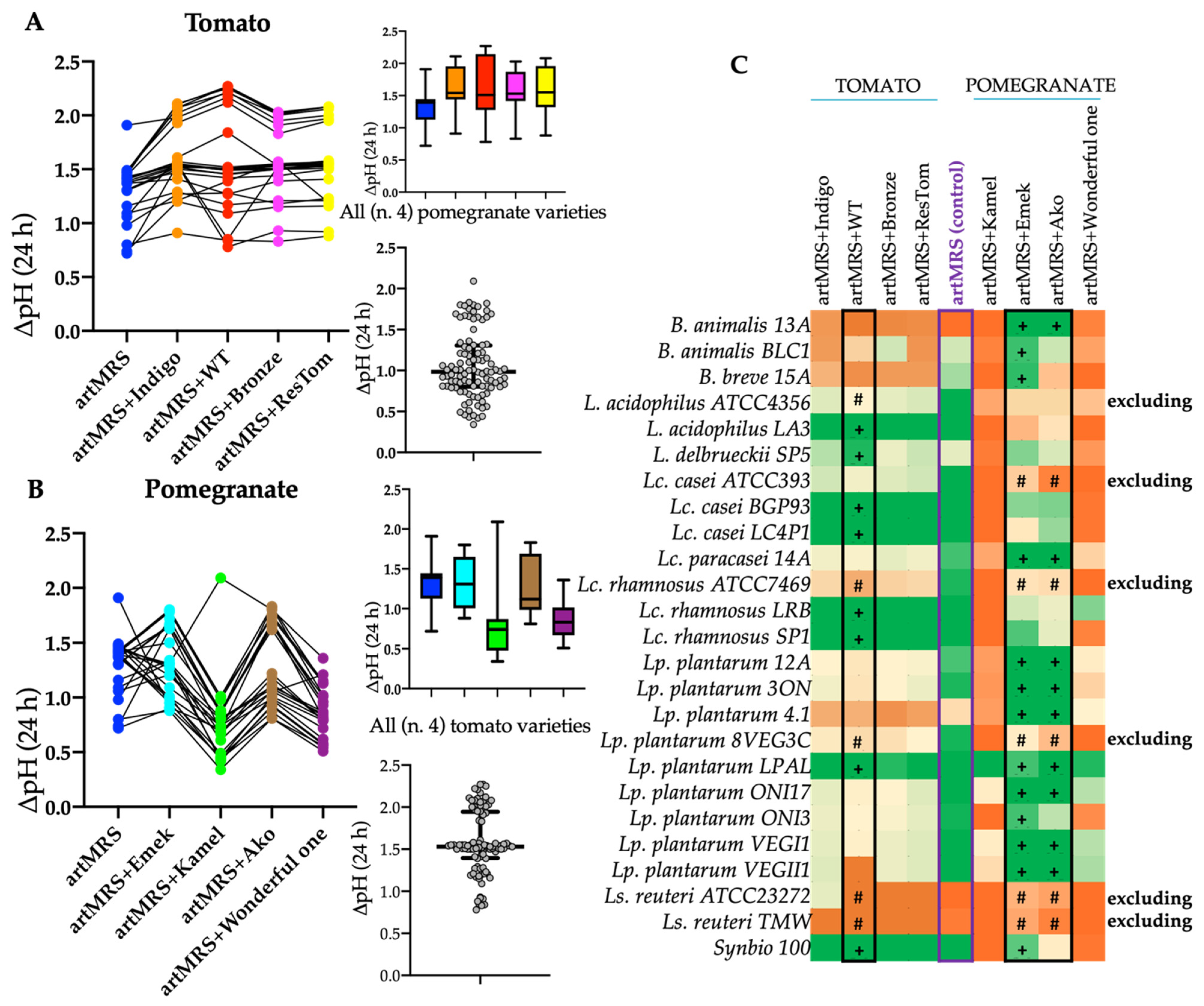
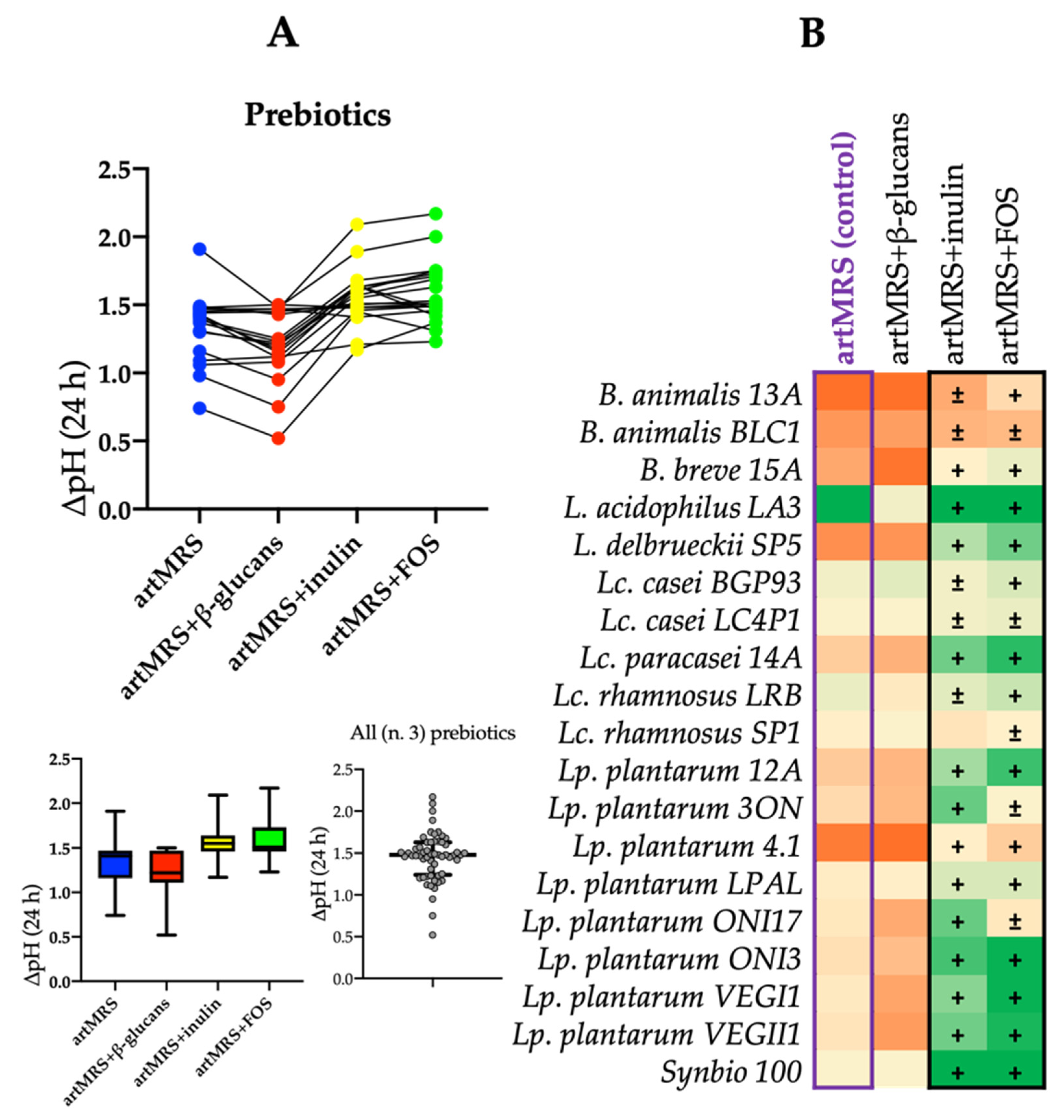
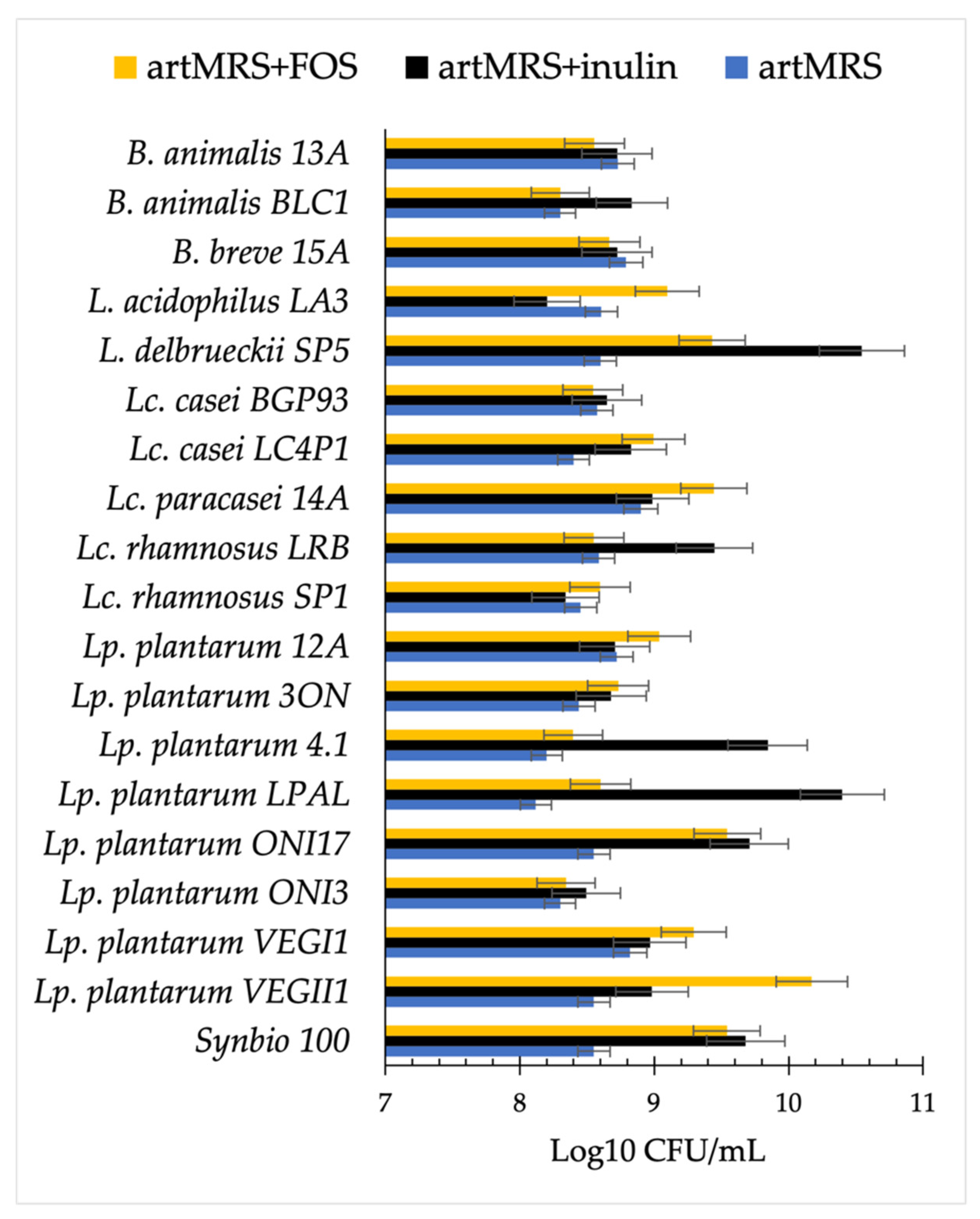
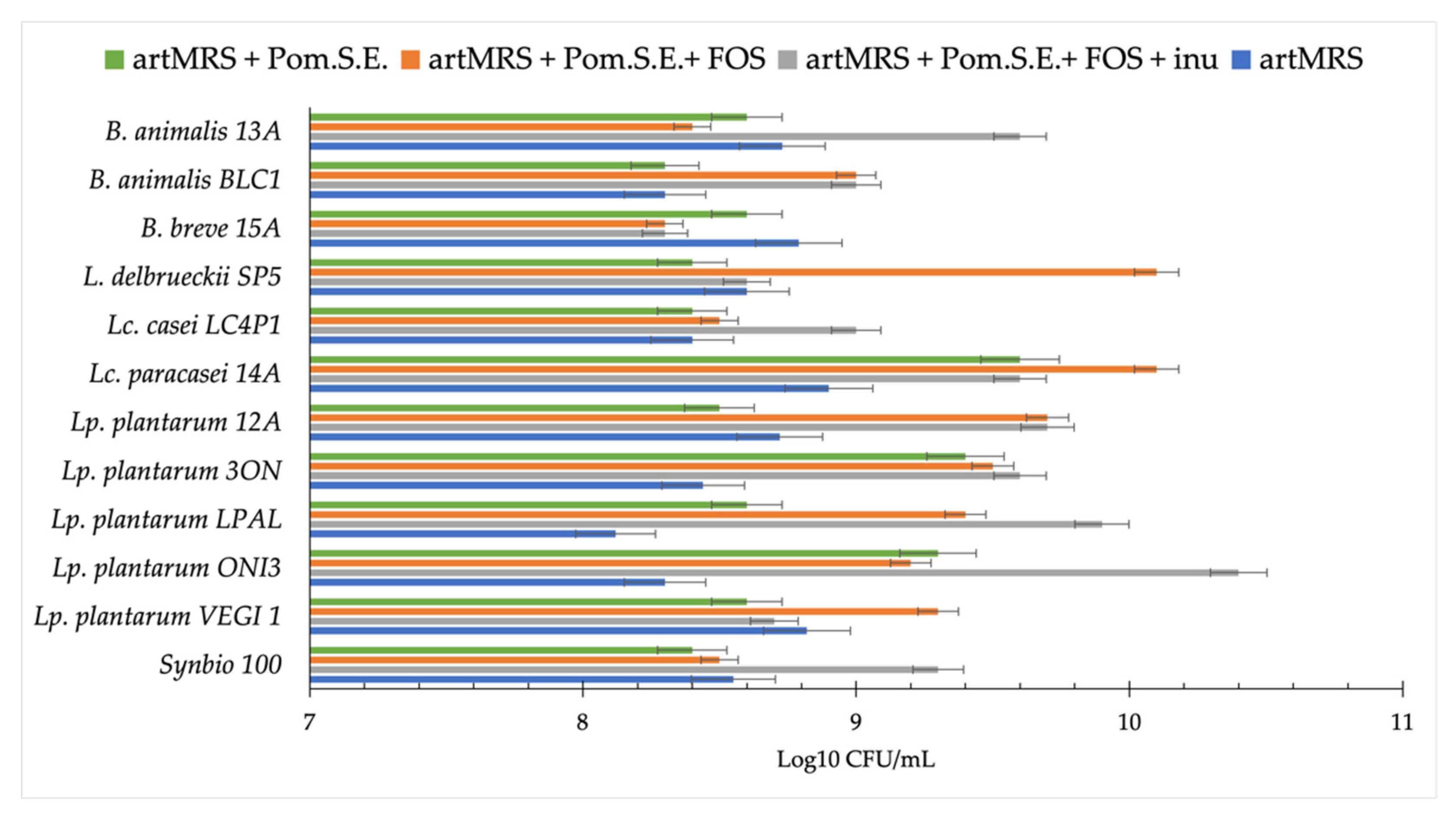
3.2. Acidification and Growth of Probiotics in Presence of Antioxidants and Food Matrices
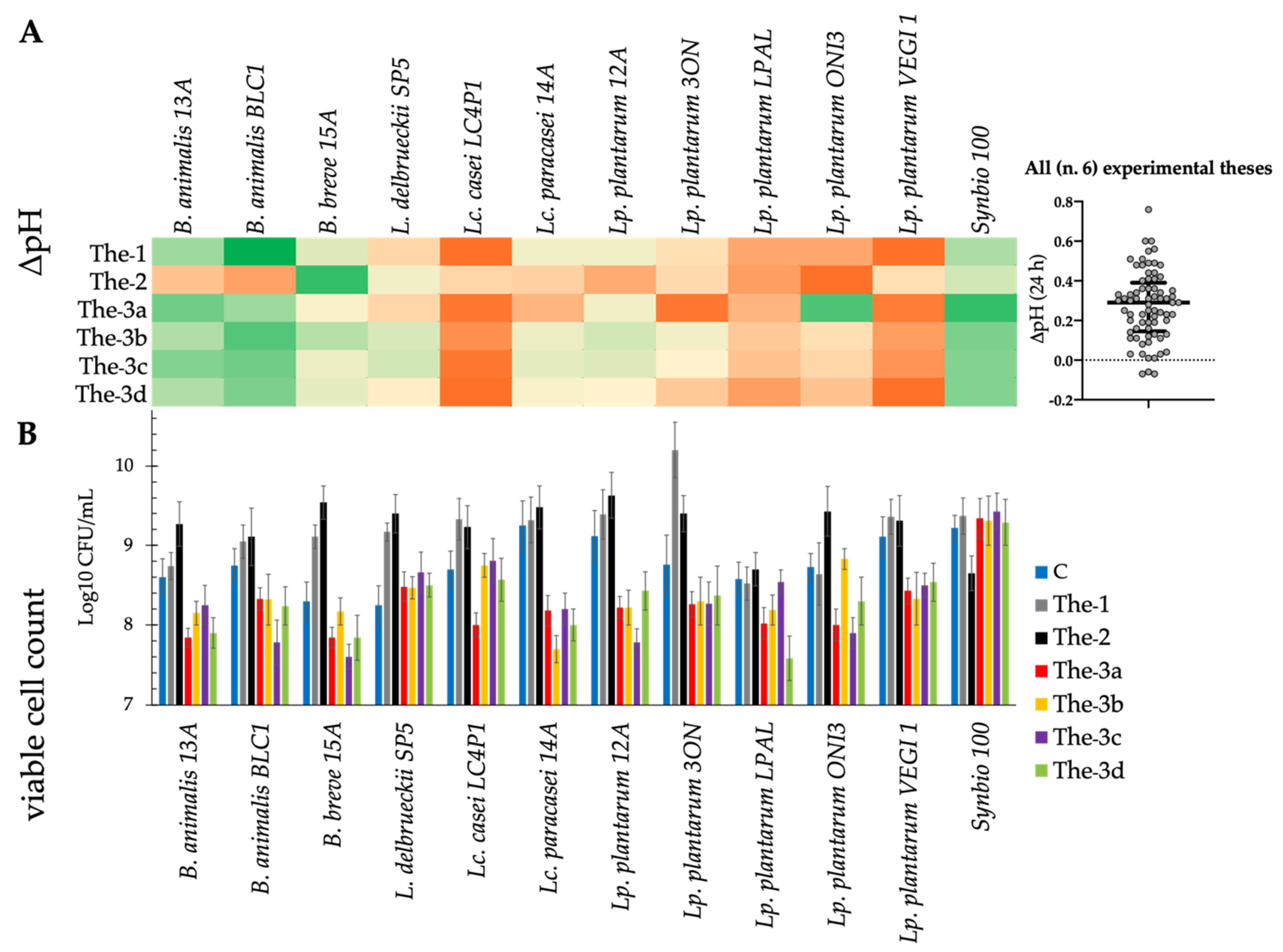
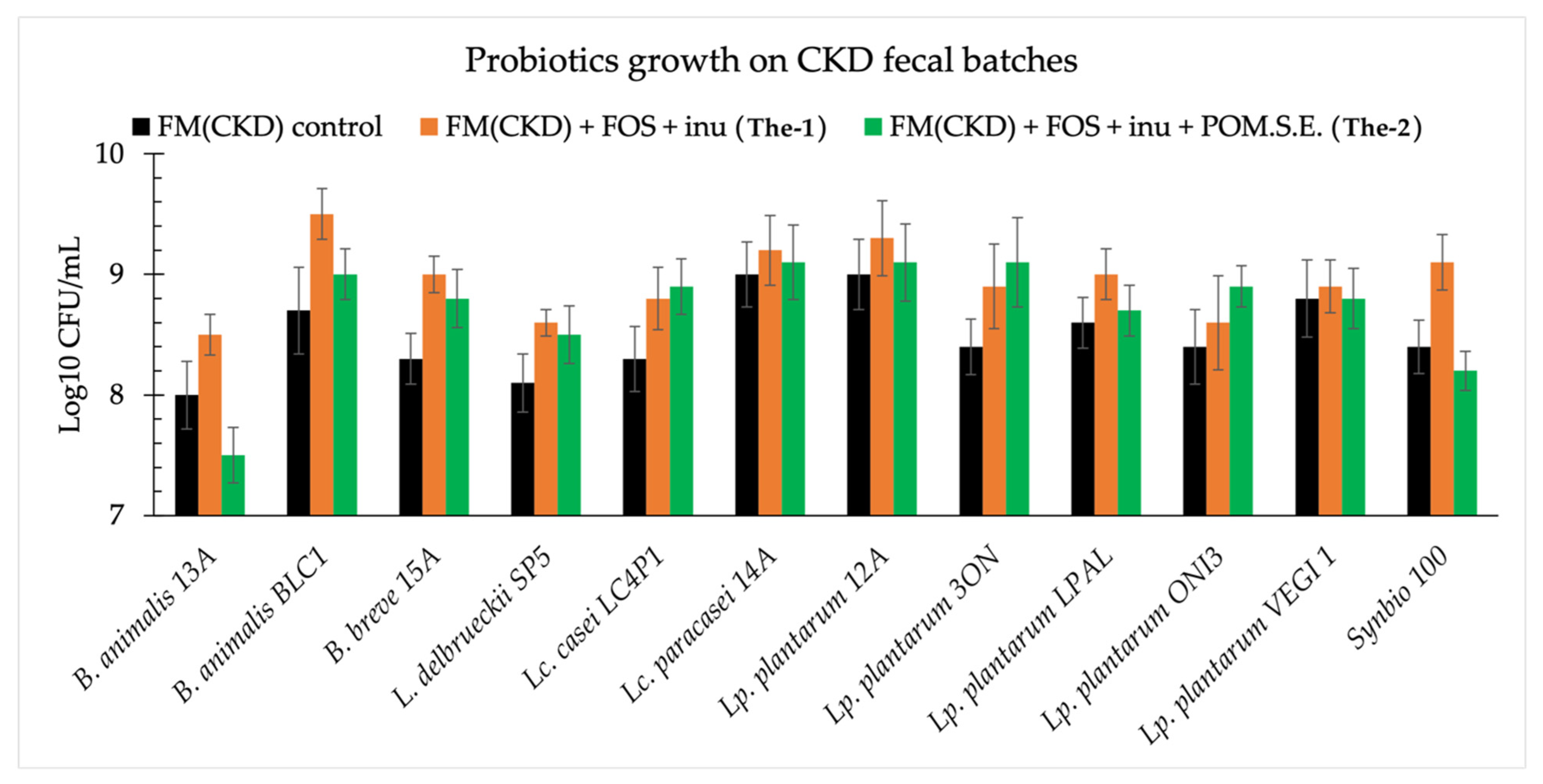
3.3. Acidification and Growth of Probiotics on Fecal Media from Healthy and Chronic Kidney Disease Subjects
- The-1: HC fecal media + FOS + inulin.
- The-2: HC fecal media + FOS inulin + Pom.S.E.
- The-3a: HC fecal media + FOS + inulin + tomato extract variety ResTom.
- The-3b: HC fecal media + FOS + inulin + tom. ex. var. Indigo.
- The-3c: HC fecal media + FOS + inulin + tom. ex. var. Bronze.
- The-3d: HC fecal media + FOS + inulin + tom. ex. from WT tomato.
- FM(CKD) + FOS + inu (The-1) = CKD fecal extract + glucose (2 g/L) + FOS (2.5 g/L) + inulin (2.5 g/L);
- FM(CKD) + FOS + inu + Pom.S.E. (The-2) = CKD fecal extract + glucose (2 g/L) + FOS (2.5 g/L) + inulin (2.5 g/L) + pomegranate seed extract (5 g/L).
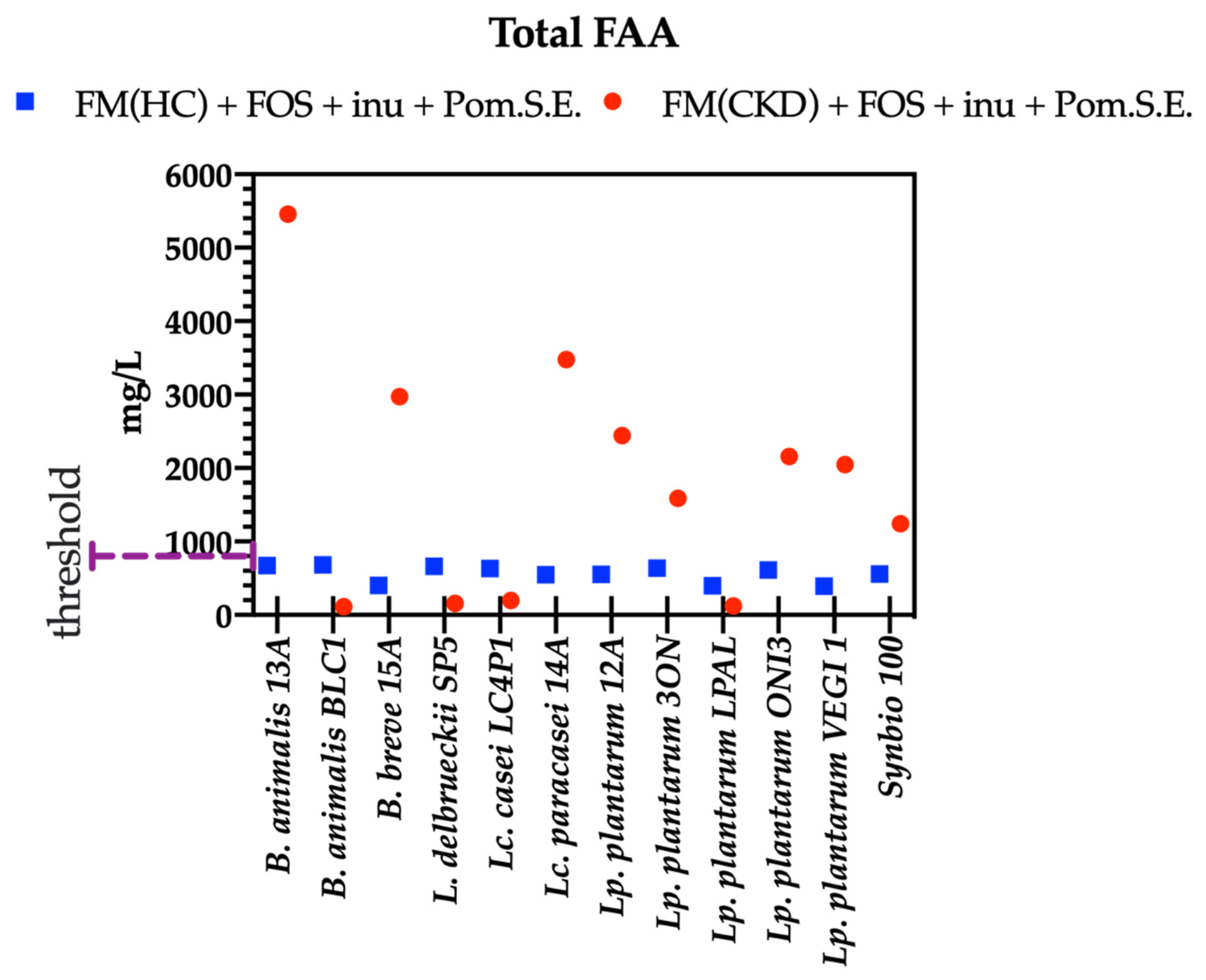
3.4. Concentrations of Total Free Amino Acids
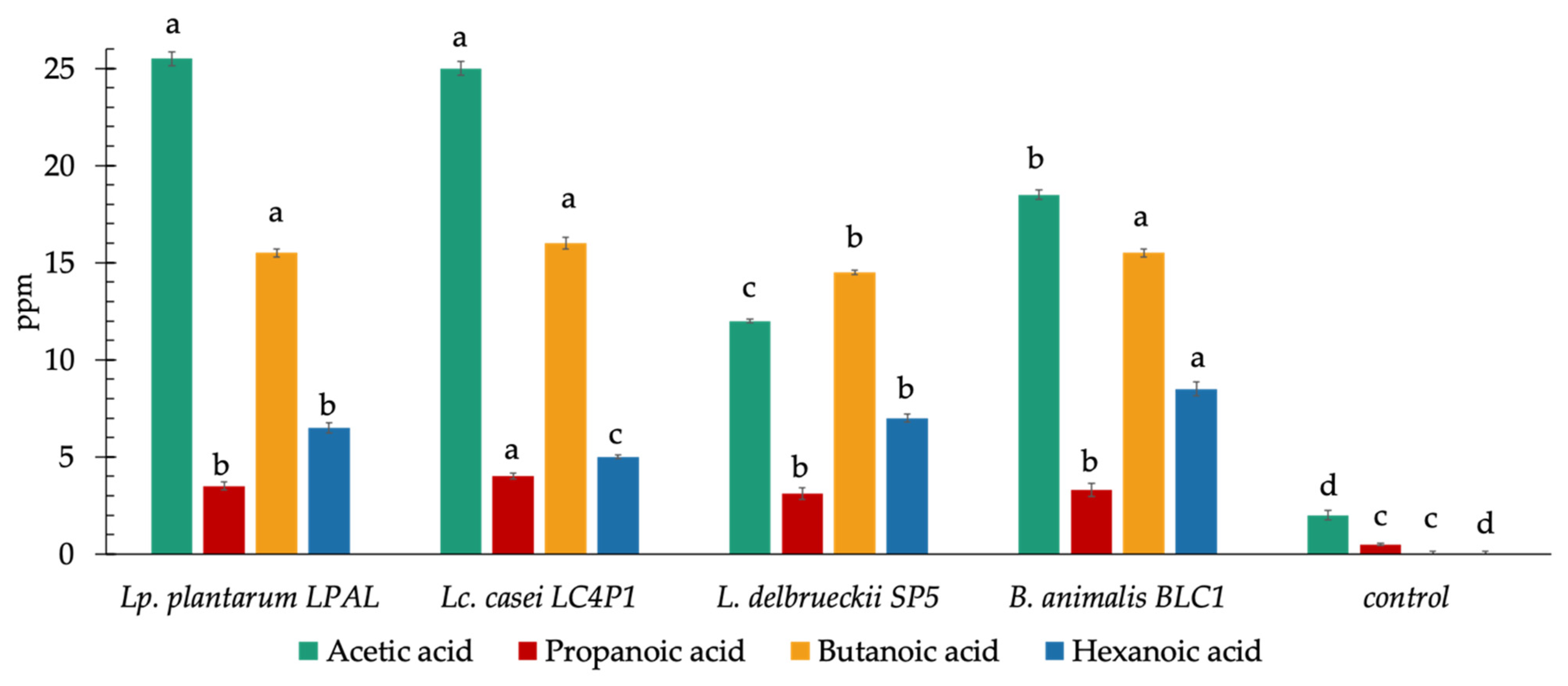
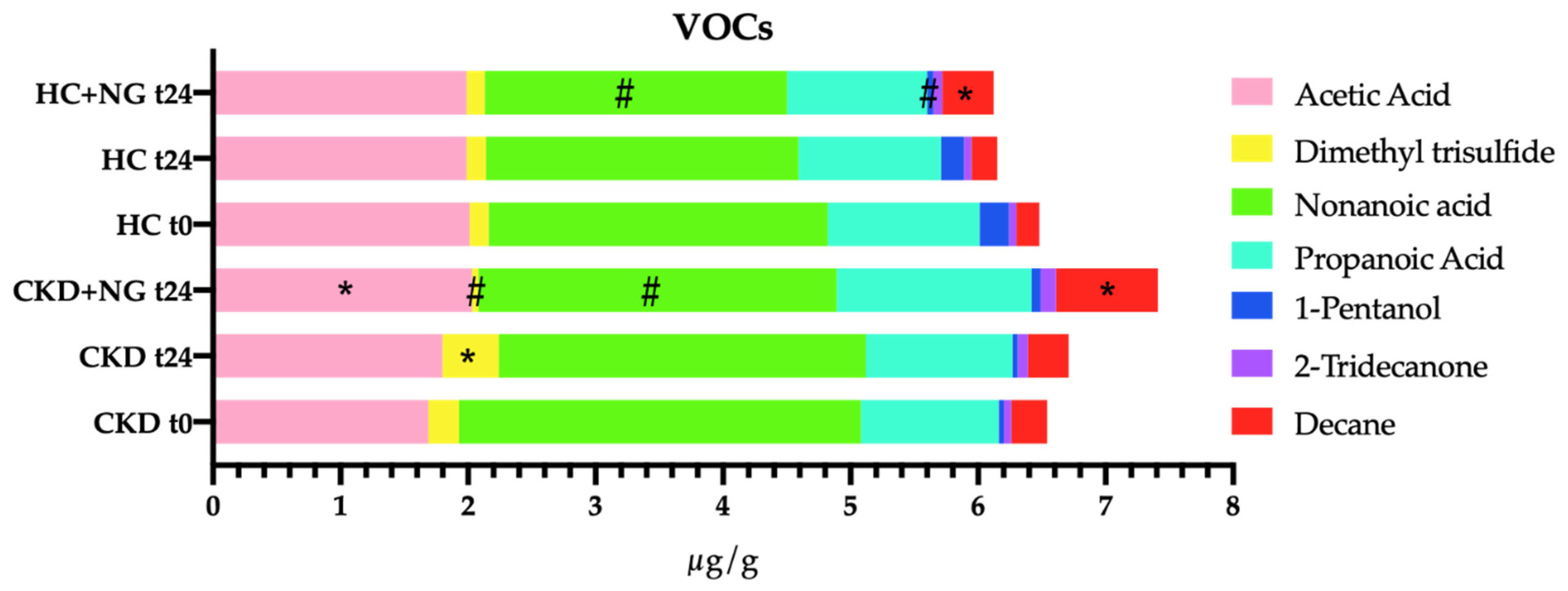
3.5. Concentration of Short Chain Fatty Acids
3.6. Stability Test of the Innovative Synbiotic Formulation
- B. animalis BLC1 (ca. 0.24 g = 109 cells);
- Lc. casei LC4P1 (ca. 0.24 g = 109 cells);
- FOS (2.5 g);
- Inulin (2.5 g);
- Pomegranate seed extract (0.2 g);
- Maltodextrins from corn (0.5 g);
- Sodium cyclamate (0.01 g).
- B. animalis BLC1 (ca. 0.24 g = 109 cells);
- Lc. casei LC4P1 (ca. 0.24 g = 109 cells);
- FOS (2.5 g);
- Inulin (2.5 g);
- Quercetin (0.064 g);
- Resveratrol (0.023 g);
- Grapeseed (Vitis vinifera L.) powder extract (0.013 g), proanthocyanidins (tit. 95% d.extr.);
- Maltodextrins from corn (0.5 g);
- Sodium cyclamate (0.01 g).
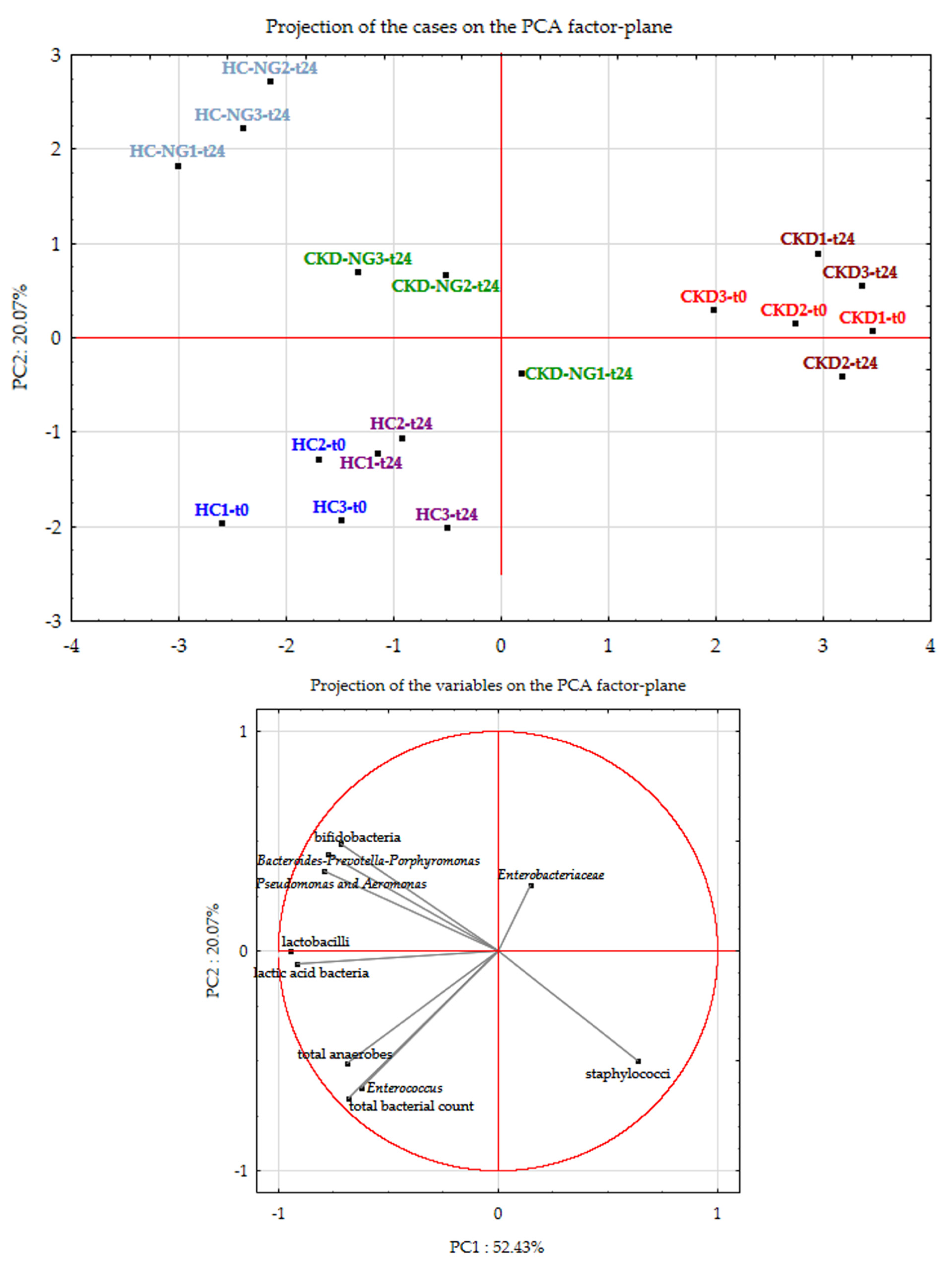
3.7. Effects of NatuREN G on Fecal Microbiota of HC and CKD Subjects
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
Appendix A.1. Pomegranate Juice Extraction
Appendix A.2. Determination of Moisture, Ash, Total Soluble Solids, and pH
Appendix A.3. Total Carbohydrates, Total Polysaccharides, Pectins, and Starch Determination
Appendix A.4. Analysis of Soluble Sugars and Organic Acids
Appendix A.5. Ascorbic Acid (AsA) and Dehydroascorbic Acid (DHA) Determination
Appendix A.6. Extraction and Determination of Soluble and Insoluble-Bound Phenolics
Appendix A.7. Extraction and Determination of Flavonoids and Proanthocyanidins
Appendix A.8. Extraction and Determination of Total Anthocyanins
Appendix A.9. Antioxidant Activity Determination
References
- Levey, A.S.; Eckardt, K.U.; Tsukamoto, Y.; Levin, A.; Coresh, J.; Rossert, J.; Zeeuw, D.D.E.; Hostetter, T.H.; Lameire, N.; Eknoyan, G. Definition and classification of chronic kidney disease: A position statement from Kidney Disease: Improving Global Outcomes (KDIGO). Kidney Int. 2005, 67, 2089–2100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, C.Y.; Iribarren, C.; McCulloch, C.E.; Darbinian, J.; Go, A.S. Risk factors for end-stage renal disease: 25-year follow-up. Arch. Intern. Med. 2009, 169, 342–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stubbs, J.R.; House, J.A.; Ocque, A.J.; Zhang, S.; Johnson, C.; Kimber, C.; Schmidt, K.; Gupta, A.; Wetmore, J.B.; Nolin, T.D.; et al. Serum trimethylamine-N-oxide is elevated in CKD and correlates with coronary atherosclerosis burden. J. Am. Soc. Nephrol. 2016, 27, 305–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anders, H.J.; Andersen, K.; Stecher, B. The intestinal microbiota, a leaky gut, and abnormal immunity in kidney disease. Kidney Int. 2013, 83, 1010–1016. [Google Scholar] [CrossRef] [Green Version]
- Mikusic, N.L.R.; Kouyoumdzian, N.M.; Choi, M.R. Gut microbiota and chronic kidney disease: Evidence and mechanisms that mediate a new communication in the gastrointestinal-renal axis. Pflügers Arch. Eur. J. Physiol. 2020, 472, 303–320. [Google Scholar] [CrossRef]
- Claro, L.M.; Moreno-Amaral, A.N.; Gadotti, A.C.; Dolenga, C.J.; Nakao, L.S.; Azevedo, M.L.; De Noronha, L.; Olandoski, M.; De Moraes, T.P.; Stinghen, A.E.M.; et al. The impact of uremic toxicity induced inflammatory response on the cardiovascular burden in chronic kidney disease. Toxins 2018, 10, 384. [Google Scholar] [CrossRef] [Green Version]
- Wu, P.; Lin, T.; Ho, H.J.; Tseng, C.; Lin, Y.; Liang, S.; Lee, H.; Kuo, M.; Hung, S.; Chiu, Y.; et al. Differences in Gut Microbiota proles and Functions between End-Stage Renal Disease and Healthy Populations. Available online: https://www.researchsquare.com/article/rs-82652/v1 (accessed on 4 February 2021).
- Gryp, T.; Huys, G.R.; Joossens, M.; Van Biesen, W.; Glorieux, G.; Vaneechoutte, M. Isolation and quantification of uremic toxin precursor-generating gut bacteria in chronic kidney disease patients. Int. J. Mol. Sci. 2020, 21, 1986. [Google Scholar] [CrossRef] [Green Version]
- Evenepoel, P.; Poesen, R.; Meijers, B. The gut–kidney axis. Pediatr. Nephrol. 2017, 32, 2005–2014. [Google Scholar] [CrossRef]
- Lehto, M.; Groop, P.H. The gut-kidney axis: Putative interconnections between gastrointestinal and renal disorders. Front. Endocrinol. 2018, 9, 553. [Google Scholar] [CrossRef]
- Di Iorio, B.R.; Rocchetti, M.T.; De Angelis, M.; Cosola, C.; Marzocco, S.; Di Micco, L.; di Bari, I.; Accetturo, M.; Vacca, M.; Gobbetti, M.; et al. Nutritional therapy modulates intestinal microbiota and reduces serum levels of total and free indoxyl sulfate and p-cresyl sulfate in chronic kidney disease (Medika Study). J. Clin. Med. 2019, 8, 1424. [Google Scholar] [CrossRef] [Green Version]
- Fernandez-Prado, R.; Esteras, R.; Perez-Gomez, M.V.; Gracia-Iguacel, C.; Gonzalez-Parra, E.; Sanz, A.B.; Ortiz, A.; Sanchez-Nino, M.D. Nutrients turned into toxins: Microbiota modulation of nutrient properties in chronic kidney disease. Nutrients 2017, 9, 489. [Google Scholar] [CrossRef] [Green Version]
- Rocchetti, M.T.; Di Iorio, B.R.; Vacca, M.; Cosola, C.; Marzocco, S.; Bari, I.D.; Calabrese, F.M.; Ciarcia, R.; De Angelis, M.; Gesualdo, L. Ketoanalogs’ effects on intestinal microbiota modulation and uremic toxins serum levels in chronic kidney disease (Medika2 Study). J. Clin. Med. 2021, 10, 840. [Google Scholar] [CrossRef] [PubMed]
- Borges, N.A.; Carmo, F.L.; Stockler-Pinto, M.B.; de Brito, J.S.; Dolenga, C.J.; Ferreira, D.C.; Nakao, L.S.; Rosado, A.; Fouque, D.; Mafra, D. Probiotic supplementation in chronic kidney disease: A double-blind, randomized, placebo-controlled trial. J. Ren. Nutr. 2018, 28, 28–36. [Google Scholar] [CrossRef] [Green Version]
- Natarajan, R.; Pechenyak, B.; Vyas, U.; Ranganathan, P.; Weinberg, A.; Liang, P.; Mallappallil, M.C.; Norin, A.J.; Friedman, E.A.; Saggi, S.J. Randomized controlled trial of strain-specific probiotic formulation (Renadyl) in dialysis patients. BioMed Res. Int. 2014, 2014, 568571. [Google Scholar] [CrossRef]
- Dehghani, H.; Heidari, F.; Mozaffari-Khosravi, H.; Nouri-Majelan, N.; Dehghani, A. Synbiotic supplementations for azotemia in patients with chronic kidney disease: A randomized controlled trial. Iran. J. Kidney Dis. 2016, 10, 351–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cosola, C.; De Angelis, M.; Rocchetti, M.T.; Montemurno, E.; Maranzano, V.; Dalfino, G.; Manno, C.; Zito, A.; Gesualdo, M.; Ciccone, M.M.; et al. Beta-glucans supplementation associates with reduction in p-cresyl sulfate levels and improved endothelial vascular reactivity in healthy individuals. PLoS ONE 2017, 12, e0169635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Den Besten, G.; van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid. Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campos-Perez, W.; Martinez-Lopez, E. Effects of short chain fatty acids on metabolic and inflammatory processes in human health. BBA Mol. Cell Biol. Lipids 2021, 1866, 158900. [Google Scholar] [CrossRef]
- Pluznick, J.L. Gut microbiota in renal physiology: Focus on short-chain fatty acids and their receptors. Kidney Int. 2016, 90, 1191–1198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Ma, L.; Fu, P. Gut microbiota-derived short-chain fatty acids and kidney diseases. Drug Des. Dev. Ther. 2017, 11, 3531. [Google Scholar] [CrossRef] [Green Version]
- Takayama, F.; Taki, K.; Niwa, T. Bifidobacterium in gastro-resistant seamless capsule reduces serum levels of indoxyl sulfate in patients on hemodialysis. Am. J. Kidney Dis. 2003, 41 (Suppl. 1), S142–S145. [Google Scholar] [CrossRef]
- Borges, N.A.; Stenvinkel, P.; Bergman, P.; Qureshi, A.R.; Lindholm, B.; Moraes, C.; Stockler-Pinto, B.; Mafra, D. Effects of probiotic supplementation on trimethylamine-n-oxide plasma levels in hemodialysis patients: A pilot study. Probiotics Antimicrob. Proteins 2019, 11, 648–654. [Google Scholar] [CrossRef]
- Nakabayashi, I.; Nakamura, M.; Kawakami, K.; Ohta, T.; Kato, I.; Uchida, K.; Yoshida, M. Effects of synbiotic treatment on serum level of p-cresol in haemodialysis patients: A preliminary study. Nephrol. Dial. Transpl. 2011, 26, 1094–1098. [Google Scholar] [CrossRef] [Green Version]
- Guida, B.; Germanò, R.; Trio, R.; Russo, D.; Memoli, B.; Grumetto, L.; Barbati, F.; Cataldi, M. Effect of short-term synbiotic treatment on plasma p-cresol levels in patients with chronic renal failure: A randomized clinical trial. Nutr. Metab. Cardiovasc. Dis. 2014, 24, 1043–1049. [Google Scholar] [CrossRef]
- Rossi, M.; Johnson, D.W.; Morrison, M.; Pascoe, E.M.; Coombes, J.S.; Forbes, J.M.; Szeto, C.C.; McWhinney, B.C.; Ungerer, J.P.J.; Campbell, K.L. Synbiotics easing renal failure by improving gut microbiology (SYNERGY): A randomized trial. Clin. J. Am. Soc. Nephrol. 2016, 11, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Scarano, A.; Butelli, E.; De Santis, S.; Cavalcanti, E.; Hill, L.; De Angelis, M.; Giovinazzo, G.; Chieppa, M.; Martin, C.; Santino, A. Combined dietary anthocyanins, flavonols, and stilbenoids alleviate inflammatory bowel disease symptoms in mice. Front. Nutr. 2018, 4, 75. [Google Scholar] [CrossRef] [Green Version]
- Filannino, P.; De Angelis, M.; Di Cagno, R.; Gozzi, G.; Riciputi, Y.; Gobbetti, M. How Lactobacillus plantarum shapes its transcriptome in response to contrasting habitats. Env. Microbiol. 2018, 20, 3700–3716. [Google Scholar] [CrossRef] [PubMed]
- De Angelis, M.; Montemurno, E.; Piccolo, M.; Vannini, L.; Lauriero, G.; Maranzano, V.; Gozzi, G.; Serrazanetti, D.; Dalfino, G.; Gobbetti, M.; et al. Microbiota and metabolome associated with immunoglobulin A nephropathy (IgAN). PLoS ONE 2014, 9, e99006. [Google Scholar] [CrossRef] [Green Version]
- Dixon, E.; Clubb, C.; Pittman, S.; Ammann, L.; Rasheed, Z.; Kazmi, N.; Keshavarzian, A.; Pat Gillevet, P.; Huzefa Rangwala, H.; Couch, R.D. Solid-phase microextraction and the human fecal VOC metabolome. PLoS ONE 2011, 6, e18471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Angelis, M.; Piccolo, M.; Vannini, L.; Siragusa, S.; De Giacomo, A.; Serrazzanetti, D.I.; Cristofori, F.; Guerzoni, M.E.; Gobbetti, M.; Francavilla, R. Fecal microbiota and metabolome of children with autism and pervasive developmental disorder not otherwise specified. PLoS ONE 2013, 8, e76993. [Google Scholar] [CrossRef] [Green Version]
- Cammarota, G.; Ianiro, G.; Tilg, H.; Rajilić-Stojanović, M.; Kump, P.; Satokari, R.; Sokol, H.; Arkkila, P.; Pintus, C.; Hart, A.; et al. European consensus conference on faecal microbiota transplantation in clinical practice. Gut 2017, 66, 569–580. [Google Scholar] [CrossRef]
- Beaumont, M.; Portune, K.J.; Steuer, N.; Lan, A.; Cerrudo, V.; Audebert, M.; Dumont, F.; Mancano, G.; Khodorova, N.; Andriamihaja, M.; et al. Quantity and source of dietary protein influence metabolite production by gut microbiota and rectal mucosa gene expression: A randomized, parallel, double-blind trial in overweight humans. Am. J. Clin. Nutr. 2017, 106, 1005–1019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liso, M.; De Santis, S.; Scarano, A.; Verna, G.; Dicarlo, M.; Galleggiante, V.; Campiglia, P.; Mastronardi, M.; Lippolis, A.; Vacca, M.; et al. A bronze-tomato enriched diet affects the intestinal microbiome under homeostatic and inflammatory conditions. Nutrients 2018, 10, 1862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dounousi, E.; Papavasiliou, E.; Makedou, A.; Ioannou, K.; Katopodis, K.P.; Tselepis, A.; Siamopoulos, K.C.; Tsakiris, D. Oxidative stress is progressively enhanced with advancing stages of CKD. Am. J. Kidney Dis. 2006, 48, 752–760. [Google Scholar] [CrossRef] [Green Version]
- Gosmanova, E.O.; Le, N.A. Cardiovascular complications in CKD patients: Role of oxidative stress. Cardiol. Res. Pract. 2011, 2011, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozdal, T.; Sela, D.A.; Xiao, J.; Boyacioglu, D.; Chen, F.; Capanoglu, E. The reciprocal interactions between polyphenols and gut microbiota and effects on bioaccessibility. Nutrients 2016, 8, 78. [Google Scholar] [CrossRef]
- Yahia, Y.; Benabderrahim, M.A.; Tlili, N.; Bagues, M.; Nagaz, K. Bioactive compounds, antioxidant and antimicrobial activities of extracts from different plant parts of two Ziziphus Mill. species. PLoS ONE 2020, 15, e0232599. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Huang, S.Y.; Deng, Q.; Li, G.; Su, G.; Liu, J.; Wang, H.M.D. Extraction and characterization of phenolic compounds with antioxidant and antimicrobial activities from pickled radish. Food Chem. Toxicol. 2020, 136, 111050. [Google Scholar] [CrossRef]
- Zhang, Z.; Lv, J.; Pan, L.; Zhang, Y. Roles and applications of probiotic Lactobacillus strains. Appl. Microbiol. Biot. 2018, 102, 8135–8143. [Google Scholar] [CrossRef]
- Aw, W.; Fukuda, S. Protective effects of bifidobacteria against enteropathogens. Microb. Biotechnol. 2019, 12, 1097–1100. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, L.; Delgado, S.; Ruas-Madiedo, P.; Sánchez, B.; Margolles, A. Bifidobacteria and their molecular communication with the immune system. Front. Microbiol. 2017, 8, 2345. [Google Scholar] [CrossRef] [Green Version]
- Papizadeh, M.; Rohani, M.; Nahrevanian, H.; Javadi, A.; Pourshafie, M.R. Probiotic characters of Bifidobacterium and Lactobacillus are a result of the ongoing gene acquisition and genome minimization evolutionary trends. Microb. Pathog. 2017, 111, 118–131. [Google Scholar] [CrossRef] [PubMed]
- Bellali, S.; Lagier, J.C.; Raoult, D.; Bou Khalil, J. Among live and dead bacteria, the optimization of sample collection and processing remains essential in recovering gut microbiota components. Front. Microbiol. 2019, 10, 1606. [Google Scholar] [CrossRef] [PubMed]
- Montemurno, E.; Cosola, C.; Dalfino, G.; Daidone, G.; De Angelis, M.; Gobbetti, M.; Gesualdo, L. What would you like to eat, Mr CKD microbiota? A Mediterranean diet, please! Kidney Blood Press. Res. 2014, 39, 114–123. [Google Scholar] [CrossRef]
- Lau, W.L.; Vaziri, N.D. The leaky gut and altered microbiome in chronic kidney disease. J. Ren. Nutr. 2017, 27, 458–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ardö, Y.; Pettersson, H.E. Accelerated cheese ripening with heat treated cells of Lactobacillus helveticus and a commercial proteolytic enzyme. J. Dairy Res. 1988, 55, 239–245. [Google Scholar] [CrossRef]
- Francavilla, R.; Cristofori, F.; Vacca, M.; Barone, M.; De Angelis, M. Advances in understanding the potential therapeutic applications of gut microbiota and probiotic mediated therapies in celiac disease. Expert Rev. Gastroent. 2020, 14, 323–333. [Google Scholar] [CrossRef] [PubMed]
- De Angelis, M.; Siragusa, S.; Vacca, M.; Di Cagno, R.; Cristofori, F.; Schwarm, M.; Pelzer, S.; Flügel, M.; Speckmann, B.; Francavilla, R.; et al. Selection of gut-resistant bacteria and construction of microbial consortia for improving gluten digestion under simulated gastrointestinal conditions. Nutrients 2021, 13, 992. [Google Scholar] [CrossRef]
- Esgalhado, M.; Kemp, J.A.; Azevedo, R.; Stockler-Pinto, M.B.; Dolenga, C.J.; Borges, N.A.; Nakao, L.S.; Mafra, D. Could resistant starch supplementation improve inflammatory and oxidative stress biomarkers and uremic toxins levels in hemodialysis patients? A pilot randomized controlled trial. Food Funct. 2018, 9, 6508–6516. [Google Scholar] [CrossRef]
- Meijers, B.K.I.; De Preter, V.; Verbeke, K.; Vanrenterghem, Y.; Evenepoel, P. p-Cresyl sulfate serum concentrations in haemodialysis patients are reduced by the prebiotic oligofructose-enriched inulin. Nephrol. Dial. Transplant. 2010, 25, 219–224. [Google Scholar] [CrossRef] [Green Version]
- Ramos, C.I.; Armani, R.G.; Canziani, M.E.F.; Dalboni, M.A.; Dolenga, C.J.R.; Nakao, L.S.; Campbell, K.L.; Cuppari, L. Effect of prebiotic (fructooligosaccharide) on uremic toxins of chronic kidney disease patients: A randomized controlled trial. Nephrol. Dial. Transplant. 2019, 34, 1876–1884. [Google Scholar] [CrossRef]
- Armani, R.G.; Carvalho, A.B.; Ramos, C.I.; Hong, V.; Bortolotto, L.A.; Cassiolato, J.L.; Oliveira, N.F.; Cieslarova, Z.; do Lago, C.L.; Klassen, A.; et al. Effect of fructooligosaccharide on endothelial function in CKD patients: A randomized controlled trial. Nephrol. Dial. Transplant. 2021, 7, gfaa335. [Google Scholar] [CrossRef] [PubMed]
- Association of Official Analytical Chemists. Official Methods of Analysis of AOAC International, 18th ed.; Association of Official Analytical Chemists International: Gaithersburg, MD, USA, 2005. [Google Scholar]
- Nielsen, S.S. Total carbohydrate by phenol-sulfuric acid method. In Food Analysis Laboratory Manual; Food Science Text Series; Springer: Cham, Switzerland, 2017; pp. 137–141. [Google Scholar] [CrossRef]
- Blumenkrantz, N.; Asboe-Hansen, G. New method for quantitative determination of uronic acids. Anal. Biochem. 1973, 54, 484–489. [Google Scholar] [CrossRef]
- McCleary, B.V.; Gibson, T.S.; Mugford, D.C. Measurement of total starch in cereal products by amyloglucosidase-α-amylase method: Collaborative study. J. AOAC Int. 1997, 80, 571–579. [Google Scholar] [CrossRef] [Green Version]
- Castellari, M.; Versari, A.; Spinabelli, U.; Galassi, S.; Amati, A. An improved HPLC method for the analysis of organic acids, carbohydrates and alcohols in grape musts and wines. J. Liq. Chromatogr. Relat. Technol. 2000, 23, 2047–2056. [Google Scholar] [CrossRef]
- Kampfenkel, K.; Van Montagu, M.; Inzè, D. Extraction and determination of ascorbate and dehydroascorbate from plant tissue. Anal. Biochem. 1995, 225, 165–167. [Google Scholar] [CrossRef]
- Adom, K.K.; Sorrells, M.E.; Liu, R.H. Phytochemical profiles and antioxidant activity of wheat varieties. J. Agric. Food Chem. 2003, 51, 7825–7834. [Google Scholar] [CrossRef]
- Xu, G.; Liu, D.; Chen, J.; Ye, X.; Ma, Y.; Shi, J. Juice components and antioxidant capacity of citrus varieties cultivated in China. Food Chem. 2008, 106, 545–551. [Google Scholar] [CrossRef]
- Zhishen, J.; Mengcheng, T.; Jianming, W. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Broadhurst, R.B.; Jones, W.T. Analysis of condensed tannins using acidified vanillin. J. Sci. Food Agric. 1978, 29, 788–794. [Google Scholar] [CrossRef]
- Zhao, X.; Yuan, Z.; Fang, Y.; Yin, Y.; Feng, L. Characterization and evaluation of major anthocyanins in pomegranate (Punica granatum L.) peel of different cultivars and their development phases. Eur. Food Res. Technol. 2013, 236, 109–117. [Google Scholar] [CrossRef]
- Ozgen, M.; Durgaç, C.; Serçe, S.; Kaya, C. Chemical and antioxidant properties of pomegranate cultivars grow in the Mediterranean region of Turkey. Food Chem. 2008, 111, 703–706. [Google Scholar] [CrossRef]
- Hdider, C.; Ilahy, R.; Tlili, I.; Lenucci, M.S.; Dalessandro, G. Effect of the stage of maturity on the antioxidant content and antioxidant activity of high-pigment tomato cultivars grown in Italy. Food 2013, 7, 1–7. [Google Scholar]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying and improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Probiotics | pH artMRS * | pH artMRS + Pom.S.E. | pH artMRS + Pom.S.E. + FOS | pH artMRS + Pom.S.E. + FOS + inu | (Diff.Ac.Deg.) pH artMRS − pH artMRS + Pom.S.E. + FOS + inu |
|---|---|---|---|---|---|
| B. animalis 13A | 5.66 ± 0.12 a | 5.51 ± 0.11 ab | 5.37 ± 0.11 bc | 5.26 ± 0.04 c | 0.40 ± 0.08 |
| B. animalis BLC1 | 5.31 ± 0.10 a | 5.23 ± 0.06 a | 5.02 ± 0.11 b | 4.86 ± 0.08 b | 0.45 ± 0.09 |
| B. breve 15A | 5.24 ± 0.07 a | 5.15 ± 0.10 a | 4.91 ± 0.12 bc | 4.77 ± 0.03 c | 0.47 ± 0.05 |
| L. acidophilus LA3 | 4.83 ± 0.09 a | 4.78 ± 0.10 ab | 4.65 ± 0.04 b | 4.59 ± 0.11 b | 0.24 ± 0.10 |
| L. delbrueckii SP5 | 5.34 ± 0.11 a | 5.21 ± 0.03 a | 5.02 ± 0.12 b | 4.86 ± 0.09 b | 0.48 ± 0.10 |
| Lc. casei BGP93 | 4.92 ± 0.06 a | 4.94 ± 0.08 a | 4.81 ± 0.11 ab | 4.74 ± 0.07 b | 0.18 ± 0.07 |
| Lc. casei LC4P1 | 4.93 ± 0.08 a | 4.95 ± 0.06 a | 4.70 ± 0.11 b | 4.56 ± 0.05 b | 0.37 ± 0.07 |
| Lc. paracasei 14A | 5.09 ± 0.05 a | 4.92 ± 0.05 b | 4.80 ± 0.04 c | 4.69 ± 0.11 c | 0.40 ± 0.08 |
| Lc. rhamnosus LRB | 4.91 ± 0.10 a | 4.99 ± 0.08 a | 4.88 ± 0.08 ab | 4.81 ± 0.04 b | 0.10 ± 0.07 |
| Lc. rhamnosus SP1 | 4.95 ± 0.10 a | 4.98 ± 0.02 a | 4.88 ± 0.11 ab | 4.80 ± 0.11 b | 0.15 ± 0.11 |
| Lp. plantarum 12A | 5.10 ± 0.10 a | 4.92 ± 0.02 b | 4.81 ± 0.03 c | 4.69 ± 0.12 c | 0.41 ± 0.11 |
| Lp. plantarum 3ON | 5.03 ± 0.08 a | 4.95 ± 0.10 ab | 4.84 ± 0.05 bc | 4.69 ± 0.10 c | 0.34 ± 0.09 |
| Lp. plantarum 4.1 | 5.42 ± 0.10 a | 5.37 ± 0.10 ab | 5.26 ± 0.06 bc | 5.19 ± 0.04 c | 0.23 ± 0.07 |
| Lp. plantarum LPAL | 4.96 ± 0.07 a | 4.91 ± 0.10 ab | 4.78 ± 0.06 b | 4.66 ± 0.02 c | 0.30 ± 0.05 |
| Lp. plantarum ONI17 | 4.98 ± 0.11 a | 4.97 ± 0.07 a | 4.90 ± 0.11 ab | 4.83 ± 0.05 b | 0.15 ± 0.08 |
| Lp. plantarum ONI3 | 5.01 ± 0.06 a | 4.91 ± 0.05 ab | 4.78 ± 0.08 bc | 4.67 ± 0.03 c | 0.34 ± 0.05 |
| Lp. plantarum VEGI1 | 4.97 ± 0.12 a | 4.95 ± 0.02 a | 4.78 ± 0.10 ab | 4.67 ± 0.11 b | 0.30 ± 0.12 |
| Lp. plantarum VEGII1 | 4.99 ± 0.05 a | 5.02 ± 0.11 ab | 4.89 ± 0.04 b | 4.83 ± 0.12 b | 0.16 ± 0.09 |
| Synbio 100 | 4.93 ± 0.10 a | 5.06 ± 0.10 a | 4.34 ± 0.04 b | 4.10 ± 0.10 c | 0.83 ± 0.10 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vacca, M.; Celano, G.; Lenucci, M.S.; Fontana, S.; Forgia, F.M.l.; Minervini, F.; Scarano, A.; Santino, A.; Dalfino, G.; Gesualdo, L.; et al. In Vitro Selection of Probiotics, Prebiotics, and Antioxidants to Develop an Innovative Synbiotic (NatuREN G) and Testing Its Effect in Reducing Uremic Toxins in Fecal Batches from CKD Patients. Microorganisms 2021, 9, 1316. https://doi.org/10.3390/microorganisms9061316
Vacca M, Celano G, Lenucci MS, Fontana S, Forgia FMl, Minervini F, Scarano A, Santino A, Dalfino G, Gesualdo L, et al. In Vitro Selection of Probiotics, Prebiotics, and Antioxidants to Develop an Innovative Synbiotic (NatuREN G) and Testing Its Effect in Reducing Uremic Toxins in Fecal Batches from CKD Patients. Microorganisms. 2021; 9(6):1316. https://doi.org/10.3390/microorganisms9061316
Chicago/Turabian StyleVacca, Mirco, Giuseppe Celano, Marcello Salvatore Lenucci, Sergio Fontana, Flavia Maria la Forgia, Fabio Minervini, Aurelia Scarano, Angelo Santino, Giuseppe Dalfino, Loreto Gesualdo, and et al. 2021. "In Vitro Selection of Probiotics, Prebiotics, and Antioxidants to Develop an Innovative Synbiotic (NatuREN G) and Testing Its Effect in Reducing Uremic Toxins in Fecal Batches from CKD Patients" Microorganisms 9, no. 6: 1316. https://doi.org/10.3390/microorganisms9061316