
Biochemical and Structural Aspects of Cytokinin Biosynthesis and Degradation in Bacteria


Abstract
:1. Introduction
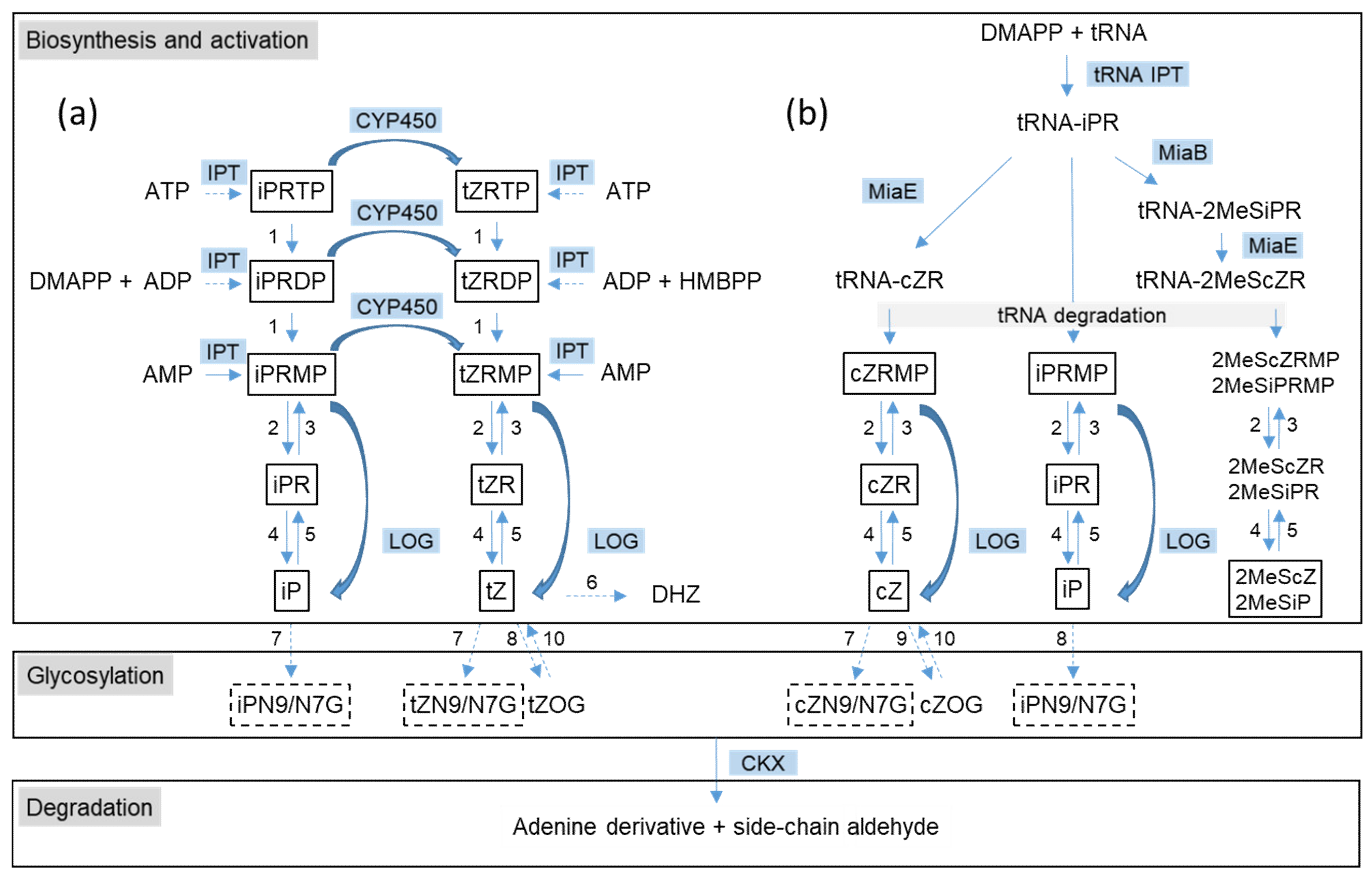
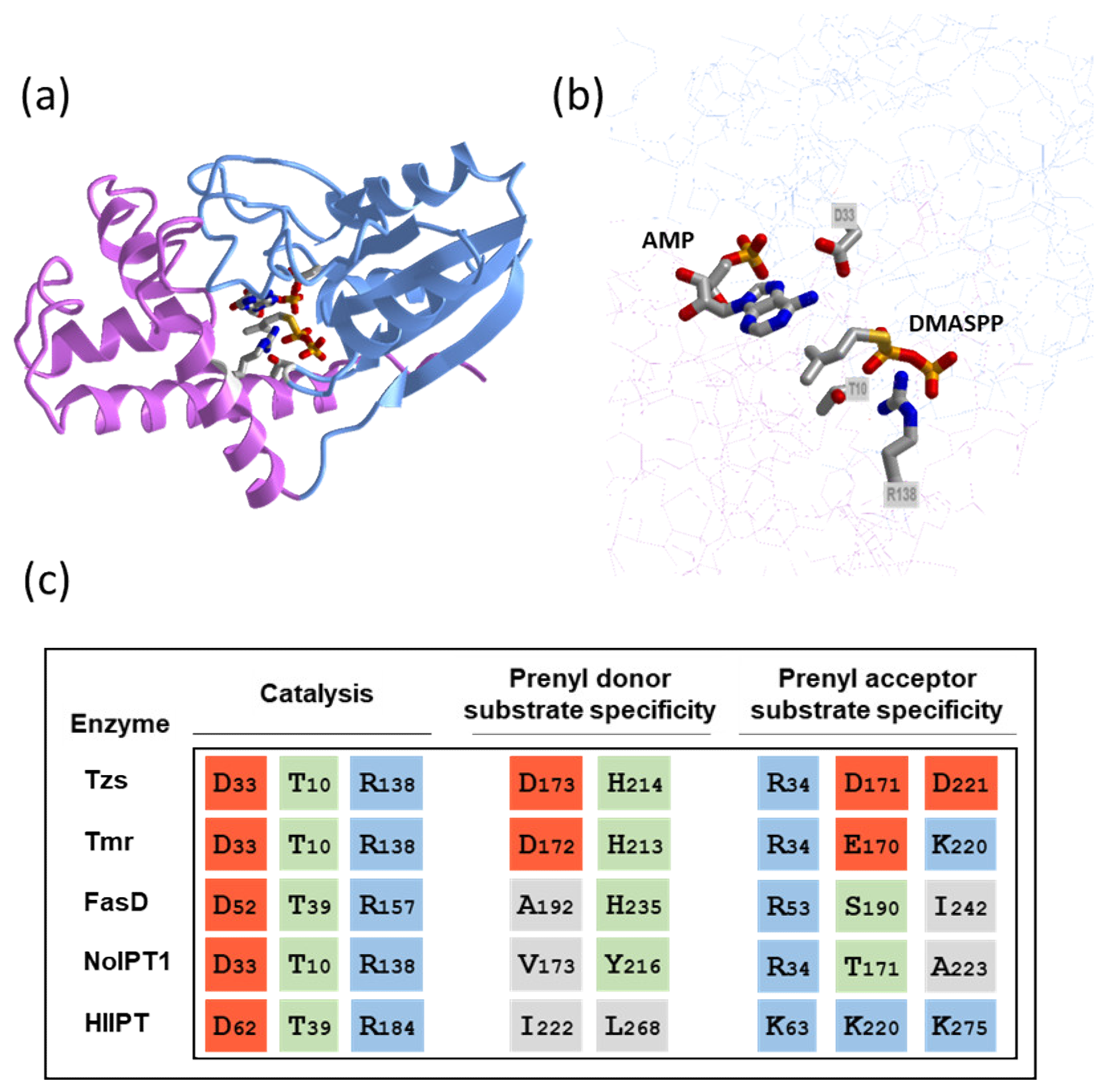
2. The Initial Step of CK Biosynthesis: Two Different Pathways
2.1. De Novo CK Biosynthesis Pathway
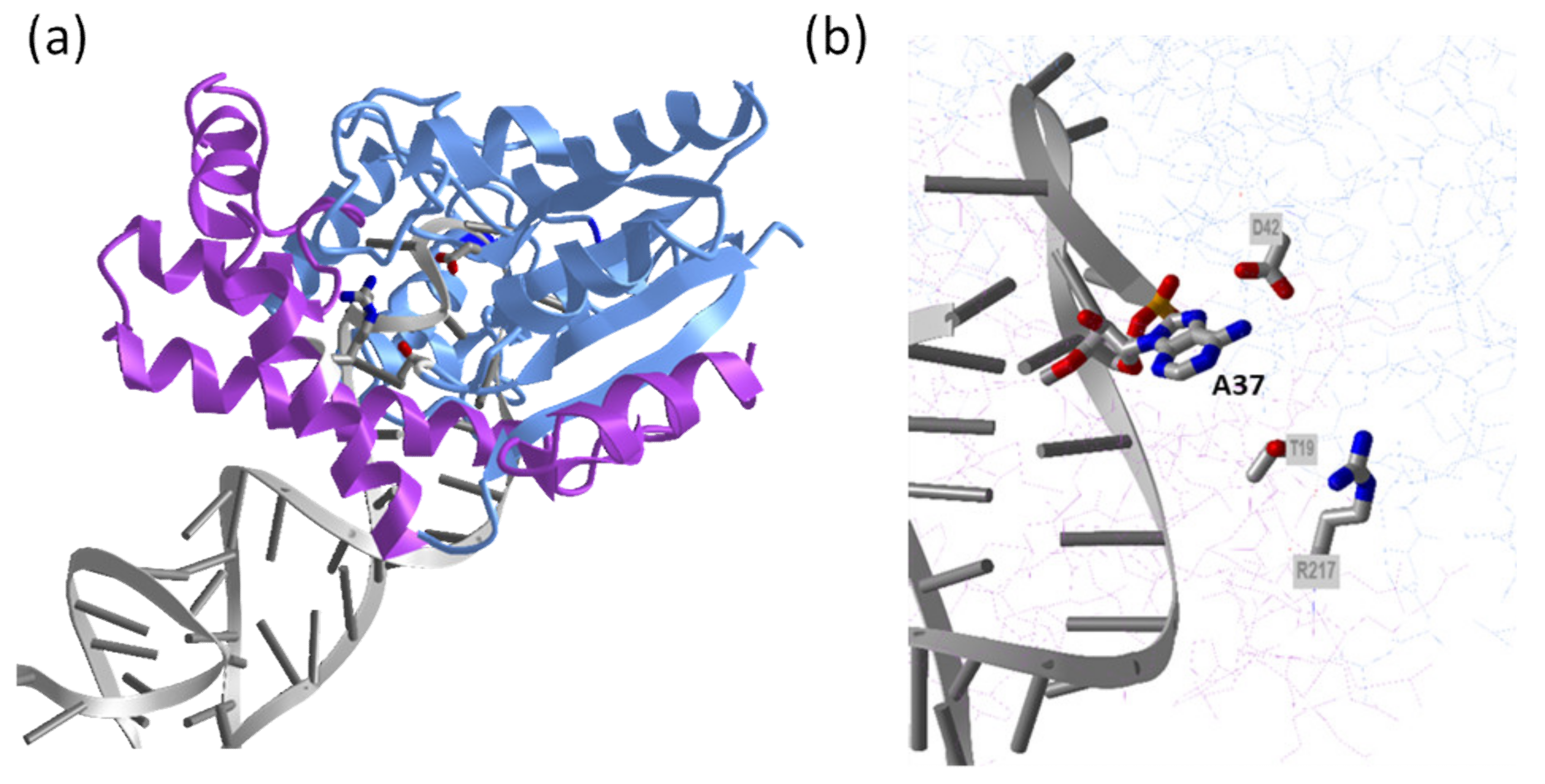
2.2. tRNA-Dependent CK Biosynthesis Pathway
3. Formation of Trans-Zeatin: Unclarified Pathways
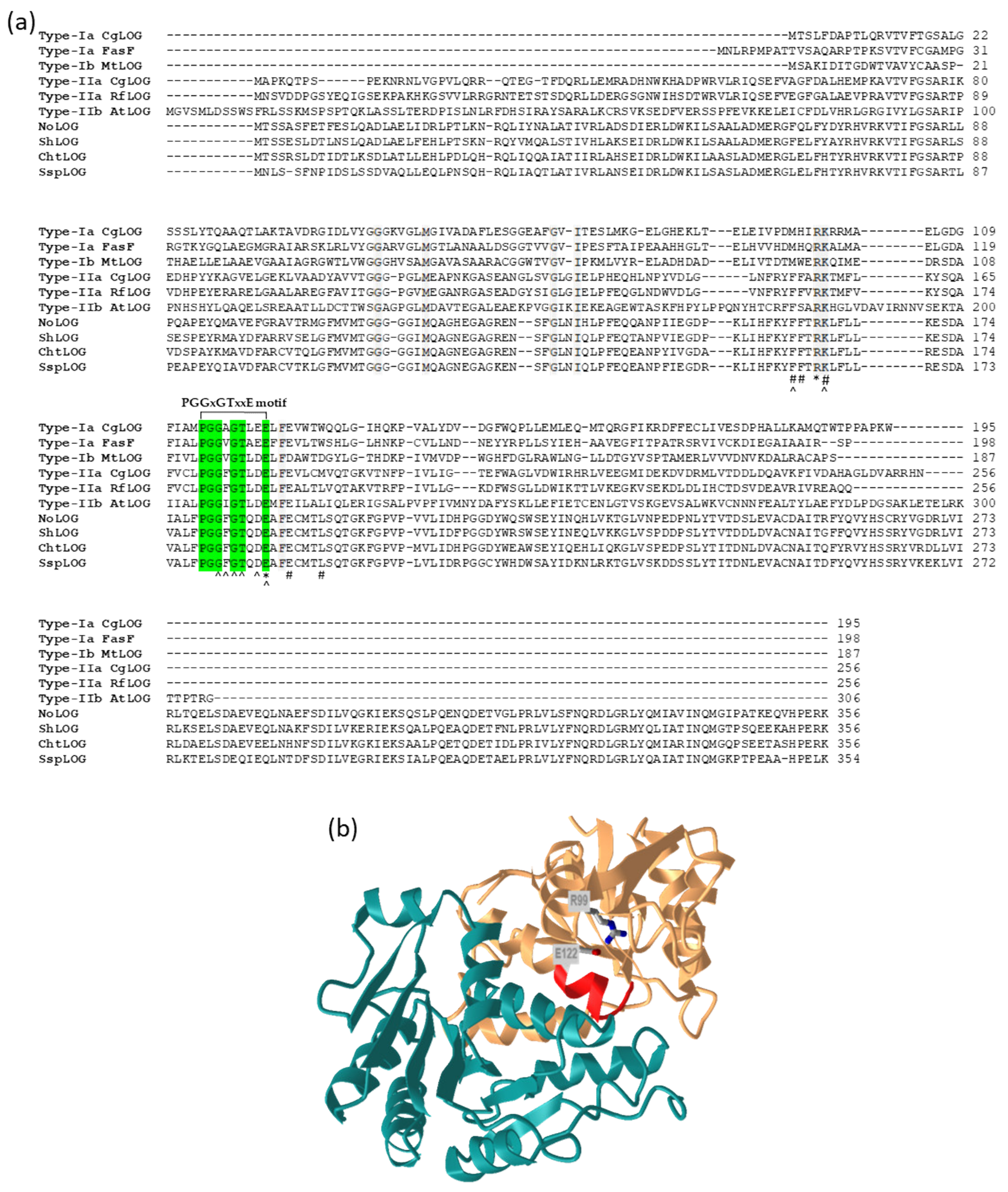
4. Cytokinin Activation: Removal of Phosphoribose by LOG
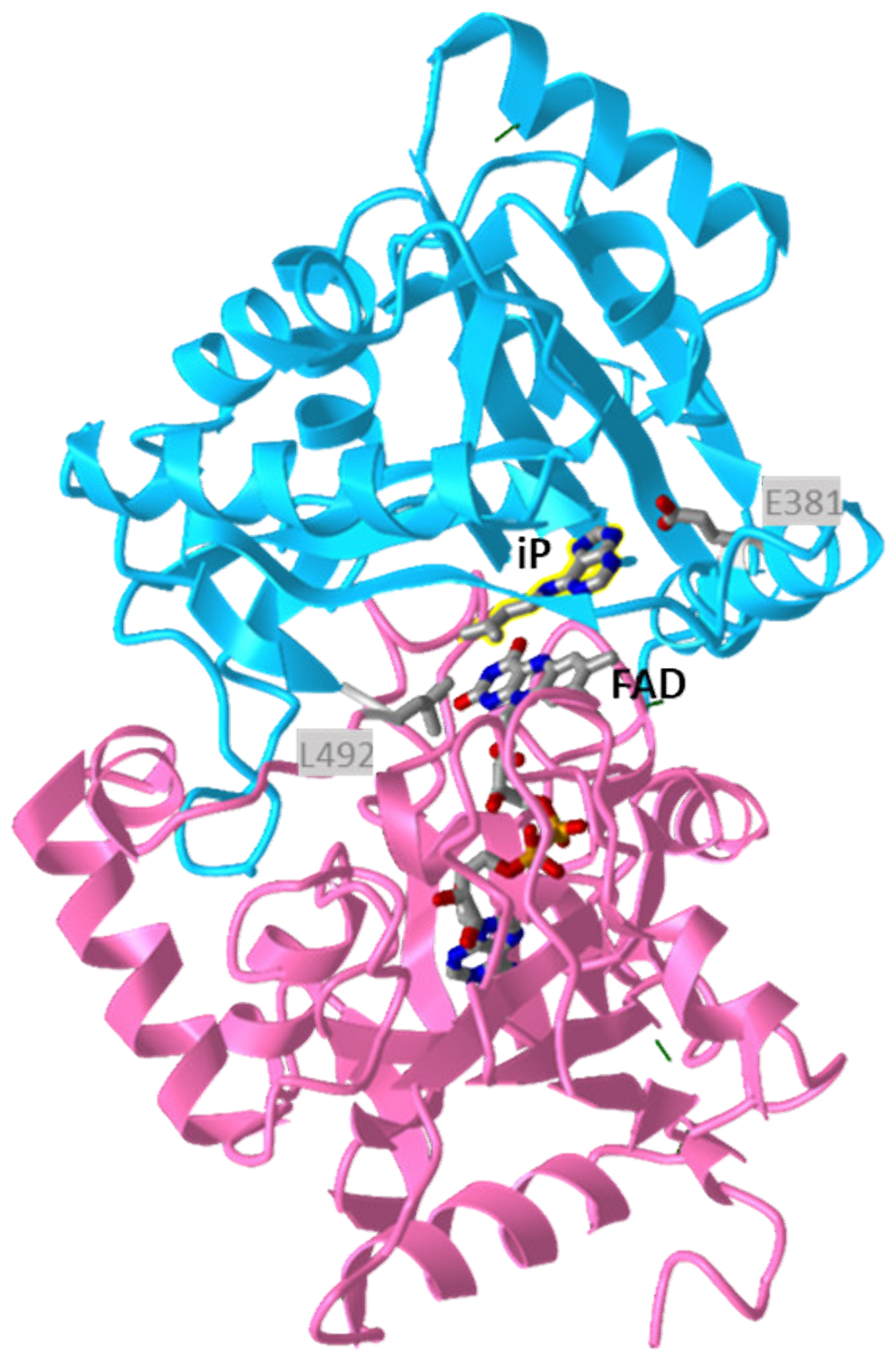
5. Degradation of Cytokinins by Cytokinin Dehydrogenase
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mok, M.C. Cytokinins and plant development—An overview. In Cytokinins—Chemistry, Activity, and Function, 1st ed.; Mok, D.W.S., Mok, M.C., Eds.; CRC Press: Boca Raton, FL, USA, 1994; pp. 155–166. [Google Scholar] [CrossRef]
- Spíchal, L. Cytokinins–recent news and views of evolutionally old molecules. Funct. Plant Biol. 2012, 39, 267–284. [Google Scholar] [CrossRef] [Green Version]
- Sakakibara, H. Cytokinins: Activity, biosynthesis, and translocation. Annu. Rev. Plant Biol. 2006, 57, 431–439. [Google Scholar] [CrossRef] [Green Version]
- Radhika, V.; Ueda, N.; Tsuboi, Y.; Kojima, M.; Kikuchi, J.; Kudo, T.; Sakakibara, H. Methylated cytokinins from the phytopathogen Rhodococcus fascians mimic plant hormone activity. Plant Physiol. 2015, 169, 1118–1126. [Google Scholar] [CrossRef] [Green Version]
- Frébort, I.; Kowalska, M.; Hluska, T.; Frébortová, J.; Galuszka, P. Evolution of cytokinin biosynthesis and degradation. J. Exp. Bot. 2011, 62, 2431–2452. [Google Scholar] [CrossRef] [PubMed]
- Takei, K.; Sakakibara, H.; Sugiyama, T. Identification of genes encoding adenylate isopentenyltransferase, a cytokinin biosynthesis enzyme, in Arabidopsis thaliana. J. Biol. Chem. 2001, 276, 26405–26410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kakimoto, T. Identification of plant cytokinin biosynthetic enzymes as dimethylallyl diphosphate:ATP/ADP isopentenyltransferases. Plant Cell Physiol. 2001, 42, 677–685. [Google Scholar] [CrossRef]
- Persson, B.C.; Esberg, B.; Olafsson, O.; Bjork, G.R. Synthesis and function of isopentenyl adenosine derivatives in transfer-RNA. Biochimie 1994, 76, 1152–1160. [Google Scholar] [CrossRef]
- Esberg, B.; Leung, H.-C.E.; Tsui, H.-C.T.; Björk, G.R.; Winkler, M.E. Identification of the miaB gene, involved in methylthiolation ofisopentenylated A37 derivatives in the tRNA of Salmonella typhimurium and Escherichia coli. J. Bacteriol. 1999, 181, 7256–7265. [Google Scholar] [CrossRef] [Green Version]
- Persson, B.C.; Björk, G.R. Isolation of the gene (miaE) encoding the hydroxylase involved in the synthesis of 2-methylthio-cis-ribozeatin in tRNA of Salmonella typhimurium and characterization of mutants. J. Bacteriol. 1993, 175, 7776–7785. [Google Scholar] [CrossRef] [Green Version]
- Kurakawa, T.; Ueda, N.; Maekawa, M.; Kobayashi, K.; Kojima, M.; Nagato, Y.; Sakakibara, H.; Kyozuka, J. Direct control of shoot meristem activity by a cytokinin-activating enzyme. Nature 2007, 445, 652–655. [Google Scholar] [CrossRef]
- Martin, R.C.; Mok, M.C.; Shaw, G.; Mok, D.W.S. An enzyme mediating the conversion of zeatin to dihydrozeatin in Phaseolus embryos. Plant Physiol. 1989, 90, 1630–1635. [Google Scholar] [CrossRef] [Green Version]
- Entsch, B.; Parker, C.W.; Letham, D.S.; Summons, R.E. Preparation and characterization, using high-performance liquid chromatography, of an enzyme forming glucosides of cytokinins. Biochim. Biophys. Acta 1979, 570, 124–139. [Google Scholar] [CrossRef]
- Martin, R.C.; Mok, M.C.; Habben, J.E.; Mok, D.W.S. A maize cytokinin gene encoding an O-glucosyltransferase specific to cis-zeatin. Proc. Natl. Acad. Sci. USA 2001, 98, 5922–5926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dixon, S.C.; Martin, R.C.; Mok, M.C.; Shaw, G.; Mok, D.W.S. Zeatin glycosylation enzymes in Phaseolus: Isolation of O-glucosyltransferase from P. lunatus and comparison to O-xylosyltransferase from P. vulgaris. Plant Physiol. 1989, 90, 1316–1321. [Google Scholar] [CrossRef] [Green Version]
- Brzobohatý, B.; Moore, I.; Kristoffersen, P.; Bako, L.; Campos, N.; Schell, J.; Palme, K. Release of active cytokinin by a beta-glucosidase localized to the maize root meristem. Science 1993, 262, 1051–1054. [Google Scholar] [CrossRef]
- Hošek, P.; Hoyerová, K.; Kiran, N.S.; Dobrev, P.I.; Zahajská, L.; Filepová, R.; Motyka, V.; Müller, K.; Kamínek, M. Distinct metabolism of N-glucosides of isopentenyladenine and trans-zeatin determines cytokinin metabolic spectrum in Arabidopsis. New Phytol. 2020, 225, 2423–2438. [Google Scholar] [CrossRef]
- Galuszka, P.; Frébort, I.; Šebela, M.; Sauer, P.; Jacobsen, S.; Peč, P. Cytokinin oxidase or dehydrogenase? Mechanism of cytokinin degradation in cereals. Eur. J. Biochem. 2001, 268, 450–461. [Google Scholar] [CrossRef] [PubMed]
- Žižková, E.; Kubeš, M.; Dobrev, P.I.; Přibyl, P.; Šimura, J.; Zahajská, L.; Záveská Drábková, L.; Novák, O.; Motyka, V. Control of cytokinin and auxin homeostasis in cyanobacteria and algae. Ann. Bot. 2017, 119, 151–166. [Google Scholar] [CrossRef] [Green Version]
- Frébortová, J.; Plíhal, O.; Florová, V.; Kokáš, F.; Kubiasová, K.; Greplová, M.; Šimura, J.; Novák, O.; Frébort, I. Light influences cytokinin biosynthesis and sensing in Nostoc (Cyanobacteria). J. Phycol. 2017, 53, 703–714. [Google Scholar] [CrossRef]
- Stirk, W.A.; van Staden, J. Potential of phytohormones as a strategy to improve microalgae productivity for biotechnological applications. Biotech. Adv. 2021, 44, 107612. [Google Scholar] [CrossRef]
- Pertry, I.; Václavíková, K.; Gemrotová, M.; Spíchal, L.; Galuszka, P.; Depuydt, S.; Temmerman, W.; Stes, E.; De Keyser, A.; Riefler, M.; et al. Rhodococcus fascians impacts plant development through the dynamic Fas-mediated production of a cytokinin mix. Mol. Plant Microbe Interact. 2010, 23, 1164–1174. [Google Scholar] [CrossRef] [Green Version]
- Samanovic, M.I.; Tu, S.; Novák, O.; Iyer, L.M.; McAllister, F.E.; Aravind, L.; Gygi, S.P.; Hubbard, S.R.; Strnad, M.; Darwin, K.H. Proteasomal control of cytokinin synthesis protects Mycobacterium tuberculosis against nitric oxide. Mol. Cell 2015, 19, 984–994. [Google Scholar] [CrossRef] [Green Version]
- Upadhyaya, N.M.; Letham, D.S.; Parker, C.W.; Hocart, C.H.; Dart, P.J. Do rhizobia produce cytokinins? Biochem. Int. 1991, 24, 123–130. [Google Scholar]
- Koenig, R.L.; Morris, R.O.; Polacco, J.C. tRNA is the source of low-level trans-zeatin production in Methylobacterium spp. J. Bacteriol. 2002, 184, 1832–1842. [Google Scholar] [CrossRef] [Green Version]
- Podlešáková, K.; Fardoux, J.; Patrel, D.; Bonaldi, K.; Novák, O.; Strnad, M.; Giraud, E.; Spíchal, L.; Nouwen, N. Rhizobial synthesized cytokinins contribute to but are not essential for the symbiotic interaction between photosynthetic Bradyrhizobia and Aeschynomene legumes. Mol. Plant. Microbe Interact. 2013, 26, 1232–1238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jorge, G.L.; Kisiala, A.; Morrison, E.; Aokia, M.; Nogueirab, A.P.O.; Emery, R.J.N. Endosymbiotic Methylobacterium oryzae mitigates the impact of limited water availability in lentil (Lens culinaris Medik.) by increasing plant cytokinin levels. Environ. Exp. Bot. 2019, 162, 525–540. [Google Scholar] [CrossRef]
- Frébortová, J.; Greplová, M.; Seidl, M.F.; Heyl, A.; Frébort, I. Biochemical characterization of putative adenylate dimethylallyltransferase and cytokinin dehydrogenase from Nostoc sp. PCC 7120. PLoS ONE 2015, 10, e0138468. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Ding, J.; Lin, S.; Liu, D.; Gu, T.; Wu, H.; Trigiano, R.N.; McAvoy, R.; Huang, J.; Li, Y. Evolution and roles of cytokinin genes in angiosperms 2: Do ancient CKXs play housekeeping roles while non-ancient CKXs play regulatory roles? Hortic. Res. 2020, 7, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lisitskaya, T.B.; Trosheva, T.D. Microorganisms stimulating plant growth for sustainable agriculture. Russ. J. Gen. Chem. 2013, 83, 2765–2774. [Google Scholar] [CrossRef]
- Akiyoshi, D.E.; Klee, H.; Amasino, R.M.; Nester, E.W.; Gordon, M.P. T-DNA of Agrobacterium tumefaciens encodes an enzyme of cytokinin biosynthesis. Proc. Natl. Acad. Sci. USA 1984, 81, 5994–5998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barry, G.F.; Rogers, S.G.; Fraley, R.T.; Brand, L. Identification of a cloned cytokinin biosynthetic gene. Proc. Natl. Acad. Sci. USA 1984, 81, 4776–4780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Powell, G.K.; Morris, R.O. Nucleotide sequence and expression of a Pseudomonas savastanoi cytokinin biosynthetic gene: Homology with Agrobacterium tumefaciens tmr and tzs loci. Nucleic Acids Res. 1986, 14, 2555–2565. [Google Scholar] [CrossRef]
- Crespi, M.; Messens, E.; Caplan, A.B.; van Montagu, M.; Desomer, J. Fasciation induction by the phytopathogen Rhodococcus fascians depends upon a linear plasmid encoding a cytokinin synthase gene. EMBO J. 1992, 11, 795–804. [Google Scholar] [CrossRef]
- Akiyoshi, D.E.; Regier, D.A.; Gordon, M.P. Nucleotide sequence of the tzs gene from Pseudomonas solanacearum strain K60. Nucleic Acids Res. 1989, 17, 8886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lichter, A.; Barash, I.; Valinsky, L.; Manulis, S. The genes involved in cytokinin biosynthesis in Erwinia herbicola pv. gypsophilae: Characterization and role in gall formation. J. Bacteriol. 1995, 177, 4457–4465. [Google Scholar] [CrossRef] [Green Version]
- Joshi, M.; Loria, R. Streptomyces turgidiscabies possesses a functional cytokinin biosynthetic pathway and produces leafy galls. Mol. Plant. Microbe Interact. 2007, 20, 751–758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serdyuk, O.P.; Shirshikova, G.N.; Smolygina, L.D.; Butanaev, A.M.; Kreslavsky, V.D.; Yartseva, N.V.; Chekunova, E.M. Agrobacterial ipt gene for cytokinin biosynthesis is found in phototrophic non-sulfur purple bacteria Rhodobacter sphaeroides and Rhodopseudomonas palustris. Russ. J. Genet. 2017, 53, 1113–1118. [Google Scholar] [CrossRef]
- Nishii, K.; Wright, F.; Chen, Y.-Y.; Möller, M. Tangled history of a multigene family: The evolution of isopentenyltransferase genes. PLoS ONE 2018, 13, e0201198. [Google Scholar] [CrossRef]
- Wang, X.; Lin, S.; Liu, D.; Gan, L.; McAvoy, R.; Ding, J.; Li, Y. Evolution and roles of cytokinin genes in angiosperms 1: Do ancient IPTs play housekeeping while non-ancient IPTs play regulatory roles? Hortic. Res. 2020, 7, 28. [Google Scholar] [CrossRef] [Green Version]
- Sakakibara, H.; Kasahara, H.; Ueda, N.; Kojima, M.; Takei, K.; Hishiyama, S.; Asami, T.; Okada, K.; Kamiya, Y.; Yamaya, T.; et al. Agrobacterium tumefaciens increases cytokinin production in plastids by modifying the biosynthetic pathway in the host plant. Proc. Natl. Acad. Sci. USA 2005, 102, 9972–9977. [Google Scholar] [CrossRef] [Green Version]
- Akiyoshi, D.E.; Regier, D.A.; Jenl, G.; Gordon, M.P. Cloning and nucleotide sequence of the tzs gene from Agrobacterium tumefaciens strain T37. Nucleic Acids Res. 1985, 13, 2773–2788. [Google Scholar] [CrossRef] [Green Version]
- Blackwell, J.R.; Horgan, R. Cloned Agrobacterium tumefaciens ipt1 gene product, DMAPP:AMP isopentenyl transferase. Phytochemistry 1993, 34, 1477–1481. [Google Scholar] [CrossRef]
- Krall, L.; Raschke, M.; Zenk, M.H.; Baron, C. The Tzs protein from Agrobacterium tumefaciens C58 produces zeatin riboside 5′-phosphate from 4-hydroxy-3-methyl-2-(E)-butenyl diphosphate and AMP. FEBS Lett. 2002, 527, 315–318. [Google Scholar] [CrossRef] [Green Version]
- Sugawara, H.; Ueda, N.; Kojima, M.; Makita, N.; Yamaya, T.; Sakakibara, H. Structural insight into reaction mechanism and evolution of cytokinin biosynthesis. Proc. Natl. Acad. Sci. USA 2008, 105, 2734–2739. [Google Scholar] [CrossRef] [Green Version]
- Crespi, M.; Vereecke, D.; Temmerman, W.; van Montagu, M.; Desomer, J. The fas operon of Rhodococcus fascians encodes new genes required for efficient fasciation of host plants. J. Bacteriol. 1994, 176, 2492–2501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jameson, P.E.; Dhandapani, P.; Song, J.; Zatloukal, M.; Strnad, M.; Remus-Emsermann, M.N.P.; Schlechter, R.O.; Novák, O. The cytokinin complex associated with Rhodococcus fascians: Which compounds are critical for virulence? Front. Plant. Sci. 2019, 10, 674. [Google Scholar] [CrossRef] [Green Version]
- Kakimoto, T. Biosynthesis of cytokinins. J. Plant. Res. 2003, 116, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Chu, H.-M.; Ko, T.-P.; Wang, A.H.-J. Crystal structure and substrate specificity of plant adenylate isopentenyltransferase from Humulus lupulus: Distinctive binding affinity for purine and pyrimidine nucleotides. Nucleic Acid Res. 2010, 38, 1738–1748. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Youkharibache, P.; Zhang, D.; Lanczycki, C.J.; Geer, R.C.; Madej, T.; Phan, L.; Ward, M.; Lu, S.; Marchler, G.H.; et al. iCn3D, a web-based 3D viewer for sharing 1D/2D/3D representations of biomolecular structures. Bioinformatics 2020, 36, 131–135. [Google Scholar] [CrossRef] [Green Version]
- Gibb, M.; Kisiala, A.; Morrison, E.; Emery, R.J.N. The origins and roles of methylthiolated cytokinins: Evidence from among life kingdoms. Front. Cell Dev. Biol. 2020, 8, 605672. [Google Scholar] [CrossRef]
- Soederberg, T.; Poulter, C.D. Escherichia coli dimethylallyl diphosphate: tRNA dimethylallyltransferase: Essential elements for recognition of tRNA substrates within the anticodon stem-loop. Biochemistry 2000, 39, 6546–6553. [Google Scholar] [CrossRef] [PubMed]
- Soderberg, T.; Poulter, C.D. Escherichia coli dimethylallyl diphosphate: tRNA dimethylallyltransferase: Site directed mutagenesis of highly conserved residues. Biochemistry 2001, 40, 1734–1740. [Google Scholar] [CrossRef]
- Xie, W.; Zhou, C.; Huang, R.H. Structure of tRNA dimethylallyltransferase: RNA modification through a channel. J. Mol. Biol. 2007, 367, 872–881. [Google Scholar] [CrossRef]
- Zhou, C.; Huang, R.H. Crystallographic snapshots of eukaryotic dimethylallyltransferase acting on tRNA: Insight into tRNA recognition and reaction mechanism. Proc. Natl. Acad. Sci. USA 2008, 105, 16142–16147. [Google Scholar] [CrossRef] [Green Version]
- Seif, E.; Hallberg, B.M. RNA-protein mutually induced fit: Structure of Escherichia coli isopentenyl-tRNA transferase in complex with tRNA(Phe). J. Biol. Chem. 2009, 284, 6600–6604. [Google Scholar] [CrossRef] [Green Version]
- Moore, J.A.; Poulter, C.D. Escherichia coli dimethylallyl diphosphate: tRNA dimethylallyltransferase: A binding mechanism for recombinant enzyme. Biochemistry 1997, 36, 604–614. [Google Scholar] [CrossRef]
- Yong, J.W.H.; Letham, D.S.; Wong, S.C.; Farquhar, G.D. Rhizobium-induced elevation in xylem cytokinin delivery in pigeonpea induces changes in shoot development and leaf physiology. Funct. Plant. Biol. 2014, 41, 1323–1335. [Google Scholar] [CrossRef] [PubMed]
- Takei, K.; Yamaya, T.; Sakakibara, H. Arabidopsis CYP735A1 and CYP735A2 encode cytokinin hydroxylases that catalyze the biosynthesis of trans-zeatin. J. Biol. Chem. 2004, 279, 41866–41872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ajitkumar, P.; Cherayil, J.D. Presence of 2-methylthioribosyl-trans-zeatin in Azotobacter vinelandii tRNA. J. Bacteriol. 1985, 162, 752–755. [Google Scholar] [CrossRef] [Green Version]
- Buck, M.; McCloskey, J.A.; Basile, B.; Ames, B.N. cis 2-Methylthio-ribosylzeatin (ms2io6A) is present in the transfer RNA of Salmonela typhimurium, but not Escherichia coli. Nucleic Acids Res. 1982, 10, 5649–5662. [Google Scholar] [CrossRef] [PubMed]
- Corder, A.L.; Subedi, B.P.; Zhang, S.; Dark, A.M.; Foss, F.W., Jr.; Pierce, B.S. Peroxide-shunt substrate-specificity for the Salmonella typhimurium O2-dependent tRNA modifying monooxygenase (MiaE). Biochemistry 2013, 52, 6182–6196. [Google Scholar] [CrossRef]
- Subedi, B.P.; Corder, A.L.; Siai Zhang, S.; Foss, F.W., Jr.; Pierce, B.S. Steady-state kinetics and spectroscopic characterization of enzyme-tRNA interactions for the non-heme diiron tRNA-monooxygenase, MiaE. Biochemistry 2015, 54, 363–376. [Google Scholar] [CrossRef]
- Kaminska, K.H.; Baraniak, U.; Boniecki, M.; Nowaczyk, K.; Czerwoniec, A.; Bujnicki, J.M. Structural bioinformatics analysis of enzymes involved in the biosynthesis pathway of the hypermodified nucleoside ms(2)io(6)A37 in tRNA. Proteins 2008, 70, 1–18. [Google Scholar] [CrossRef]
- Carpentier, P.; Lepretre, C.; Basset, C.; Douki, T.; Torelli, S.; Duarte, V.; Hamdane, D.; Fontecave, M.; Atta, M. Structural, biochemical and functional analyses of tRNA-monooxygenase enzyme MiaE from Pseudomonas putida provide insights into tRNA/MiaE interaction. Nucleic Acids Res. 2020, 48, 9918–9930. [Google Scholar] [CrossRef] [PubMed]
- Kuroha, T.; Tokunaga, H.; Kojima, M.; Ueda, N.; Ishida, T.; Nagawa, S.; Fukuda, H.; Sugimoto, K.; Sakakibara, H. Functional analyses of LONELY GUY cytokinin-activating enzymes reveal the importance of the direct activation pathway in Arabidopsis. Plant. Cell 2009, 21, 3152–3169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hinsch, J.; Vrabka, J.; Oeser, B.; Novák, O.; Galuszka, P.; Tudzynski, P. De novo biosynthesis of cytokinins in the biotrophic fungus Claviceps purpurea. Environ. Microbiol. 2015, 17, 2935–2951. [Google Scholar] [CrossRef]
- Seo, H.; Kim, K.-J. Structural basis for a novel type of cytokinin-activating protein. Sci. Rep. 2017, 7, 45985. [Google Scholar] [CrossRef]
- Dzurová, L.; Forneris, F.; Savino, S.; Galuszka, P.; Vrabka, J.; Frébort, I. The tree-dimensional structure of “Lonely Guy” from Claviceps purpurea provides insights into the phosphoribohydrolase function of Rossmann fold-containing lysine decarboxylase-like protein. Proteins 2015, 83, 1539–1546. [Google Scholar] [CrossRef]
- Francis, I.M.; Stes, E.; Zhang, Y.; Rangel, D.; Audenaert, K.; Vereecke, D. Mining the genome of Rhodococcus fascians, a plant growth-promoting bacterium gone astray. New Biotechnol. 2016, 33, 706–717. [Google Scholar] [CrossRef] [PubMed]
- Creason, A.L.; Vandeputte, O.M.; Savory, E.A.; Davis, E.W.; Putnam, M.L.; Hu, E.; Swader-Hines, D.; Mol, A.; Baucher, M.; Prinsen, E.; et al. Analysis of genome sequences from plant pathogenic Rhodococcus reveals genetic novelties in virulence loci. PLoS ONE 2014, 9, e101996. [Google Scholar] [CrossRef]
- Seo, H.; Kim, S.; Sagong, H.-Y.; Son, H.F.; Jin, K.S.; Kim, I.-K.; Kim, K.-J. Structural basis for cytokinin production by LOG from Corynebacterium glutamicum. Sci. Rep. 2016, 6, 31390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, H.; Kim, K.-J. Structural insight into molecular mechanism of cytokinin activating protein from Pseudomonas aeruginosa PAO1. Environ. Microbiol. 2018, 20, 3214–3223. [Google Scholar] [CrossRef] [PubMed]
- Seo, H.; Kim, K.-J. Structural and biochemical characterization of the type-II LOG protein from Streptomyces coelicolor A3. Biochem. Biophys. Res. Commun. 2018, 499, 577–583. [Google Scholar] [CrossRef] [PubMed]
- Moramarco, F.; Pezzicoli, A.; Salvini, L.; Leuzzi, R.; Pansegrau, W.; Balducci, E. A Lonely Guy protein of Bordetella pertussis with unique features is related to oxidative stress. Sci. Rep. 2019, 9, 17016. [Google Scholar] [CrossRef] [PubMed]
- Mayaka, J.B.; Huang, Q.; Xiao, Y.; Zhong, Q.; Ni, J.; Shen, Y. The lonely guy (LOG) homologue SiRe_0427 from the thermophilic archaeon Sulfolobus islandicus REY15A is a phosphoribohydrolase representing a novel group. Appl. Environ. Microbiol. 2019, 85, e02739-19. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Whitty, C.D.; Hall, R.H. A cytokinin oxidase in Zea mays. Can. J. Biochem. 1974, 52, 787–799. [Google Scholar] [CrossRef]
- Bilyeu, K.D.; Cole, J.L.; Laskey, J.G.; Riekhof, W.R.; Esparza, T.J.; Kramer, M.D.; Morris, R.O. Molecular and biochemical characterization of a cytokinin oxidase from maize. Plant. Phys. 2001, 125, 378–386. [Google Scholar] [CrossRef] [Green Version]
- Malito, E.; Coda, A.; Bilyeu, K.D.; Fraaije, M.W.; Mattevi, A. Structures of Michaelis and product complexes of plant cytokinin dehydrogenase: Implications for flavoenzyme catalysis. J. Mol. Biol. 2004, 341, 1237–1249. [Google Scholar] [CrossRef] [Green Version]
- Hluska, T.; Hlusková, L.; Emery, R.J.N. The Hulks and the Deadpools of the cytokinin universe: A dual strategy for cytokinin production, translocation, and signal transduction. Biomolecules 2021, 11, 209. [Google Scholar] [CrossRef]
- Morris, R.O.; Bilyeu, K.D.; Laskey, J.G.; Cheikh, N.N. Isolation of a gene encoding a glycosylated cytokinin oxidase from maize. Biochem. Biophys. Res. Commun. 1999, 255, 328–333. [Google Scholar] [CrossRef]
- Houba-Herin, N.; Pethe, C.; d’Alayer, J.; Laloue, M. Cytokinin oxidase from Zea mays: Purification, cDNA cloning and expression in moss protoplasts. Plant J. 1999, 17, 615–626. [Google Scholar] [CrossRef]
- Schmülling, T.; Werner, T.; Riefler, M.; Krupková, E.; Bartrina y Manns, I. Structure and function of cytokinin oxidase/dehydrogenase genes of maize, rice, Arabidopsis and other species. J. Plant Res. 2003, 116, 241–252. [Google Scholar] [CrossRef]
- Kopečný, D.; Končitíková, R.; Popelka, H.; Briozzo, P.; Vigouroux, A.; Kopečná, M.; Zalabák, D.; Šebela, M.; Skopalová, J.; Frébort, I.; et al. Kinetic and structural investigation of the cytokinin oxidase/dehydrogenase active site. FEBS J. 2016, 283, 361–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heuts, D.P.H.M.; Scrutton, N.S.; McIntire, W.S.; Fraaije, M.W. What’s in a covalent bond? On the role and formation of covalently bound flavin cofactors. FEBS J. 2009, 276, 3405–3427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popelková, H.; Galuzska, P.; Frébortová, J.; Bilyeu, K.D.; Frébort, I. Cytokinin dehydrogenase: Characterization and structure homology modeling of the flavoprotein catabolizing plant hormones cytokinins. In Recent Research Developments in Proteins; Pandalai, S.G., Ed.; Transworld Research Network: Kerala, India, 2004; Volume 2, pp. 63–81. [Google Scholar]
- Bae, E.; Bingman, C.A.; Bitto, E.; Aceti, D.J.; Phillips, G.N., Jr. Crystal structure of Arabidopsis thaliana cytokinin dehydrogenase. Proteins 2008, 70, 303–306. [Google Scholar] [CrossRef]
- Frébortová, J.; Fraaije, M.W.; Galuszka, P.; Šebela, M.; Peč, P.; Hrbáč, J.; Novák, O.; Bilyeu, K.D.; English, J.T.; Frébort, I. Catalytic reaction of cytokinin dehydrogenase: Preference for quinones as electron acceptors. Biochem. J. 2004, 380, 121–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galuszka, P.; Popelková, H.; Werner, T.; Frébortová, J.; Pospíšilová, H.; Mik, V.; Köllmer, I.; Schmülling, T.; Frébort, I. Biochemical characterization and histochemical localization of cytokinin oxidases/dehydrogenases from Arabidopsis thaliana expressed in Nicotiana tabaccum L. J. Plant Growth Regul. 2007, 26, 255–267. [Google Scholar] [CrossRef]
- Stirk, W.A.; van Staden, J. Flow of cytokinins through the environment. Plant Growth Regul. 2010, 62, 101–116. [Google Scholar] [CrossRef]
- Akhtar, S.S.; Mekureyaw, M.F.; Pandey, C.; Roitsch, T. Role of cytokinins for interactions of plants with microbial pathogens and pest insects. Front. Plant Sci. 2020, 10, 1777. [Google Scholar] [CrossRef] [Green Version]
- Jameson, P.E. Cytokinins and auxins in plant–pathogen interactions—An overview. Plant. Growth Regul. 2000, 32, 369–380. [Google Scholar] [CrossRef]
- Ma, K.-W.; Ma, W. Phytohormone pathways as targets of pathogens to facilitate infection. Plant Mol. Biol. 2016, 91, 713–725. [Google Scholar] [CrossRef] [Green Version]
- Pertry, I.; Václavíková, K.; Depuydt, S.; Galuszka, P.; Spíchal, L.; Temmerman, W.; Stes, E.; Schmülling, T.; Kakimoto, T.; van Montagu, M.C.E.; et al. Identification of Rhodococcus fascians cytokinins and their modus operandi to reshape the plant. Proc. Natl. Acad. Sci. USA 2009, 106, 929–934. [Google Scholar] [CrossRef] [Green Version]
- Jameson, P.E. Virulent Rhodococcus fascians produce unique methylated cytokinins. Plants 2019, 8, 582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akiyoshi, D.E.; Morris, R.O.; Hinz, R.; Mischke, B.S.; Kosuge, T.; Garfinkel, D.J.; Gordon, M.P.; Nester, E.W. Cytokinin/auxin balance in crown gall tumors is regulated by specific loci in the T-DNA. Proc. Natl. Acad. Sci. USA 1983, 80, 407–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, H.-H.; Yang, F.-J.; Cheng, T.-F.; Chen, Y.-C.; Lee, Y.-L.; Tsai, Y.-L.; Lai, E.-M. The Tzs protein and exogenous cytokinin affect virulence gene expression and bacterial growth of Agrobacterium tumefaciens. Phytopathology 2013, 103, 888–899. [Google Scholar] [CrossRef] [Green Version]
- Tsavkelova, E.A.; Klimova, S.Y.; Cherdyntseva, T.A.; Netrusov, A.I. Microbial producers of plant growth stimulators and their practical use: A review. Appl. Biochem. Microbiol. 2006, 42, 117–126. [Google Scholar] [CrossRef]
- Großkinsky, D.K.; Tafner, R.; Moreno, M.V.; Sebastian, A.; Stenglein, S.A.; García de Salamone, I.E.; Nelson, L.M.; Novák, O.; Strnad, M.; van der Graaff, E.; et al. Cytokinin production by Pseudomonas fluorescens G20-18 determines biocontrol activity against Pseudomonas syringae in Arabidopsis. Sci. Rep. 2016, 6, 23310. [Google Scholar] [CrossRef] [PubMed]
- Numan, M.; Bashira, S.; Khan, Y.; Mumtaz, R.; Shinwaric, Z.K.; Khan, A.L.; Khan, A.; AL-Harrasib, A. Plant growth promoting bacteria as an alternative strategy for salt tolerance in plants: A review. Microbiol. Res. 2018, 209, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Hussain, A.; Krischke, M.; Roitsch, T.; Hasnain, S. Rapid determination of cytokinins and auxin in cyanobacteria. Curr. Microbiol. 2010, 61, 361–369. [Google Scholar] [CrossRef] [PubMed]
- Samanovic, M.I.; Hsu, H.-C.; Jones, M.B.; Jones, V.; McNeil, M.R.; Becker, S.H.; Jordan, A.T.; Strnad, M.; Xu, C.; Jackson, M.; et al. Cytokinin signaling in Mycobacterium tuberculosis. mBio 2018, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guzzo, M.B.; Li, Q.; Nguyen, H.V.; Boom, W.H.; Nguyen, L. The Pup-proteasome system protects mycobacteria from antimicrobial antifolates. Antimicrob. Agents Chemother. 2021, 65, e01967-20. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme | Tested Substrate | Second Substrate | Km (µM) | kcat (s−1) | Reference |
|---|---|---|---|---|---|
| Tmr | AMP | DMAPP | 0.086 ± 0.008 | 4.1 × 10−1 | [43] |
| DMAPP | AMP | 8.28 ± 0.82 | n.a. | [43] | |
| DMAPP | AMP | 10.1 ± 0.5 | n.a. | [41] | |
| HMBPP | AMP | 13.6 ± 2.5 | n.a. | [41] | |
| Tzs | AMP | DMAPP | 0.035 ± 0.005 | n.a. | [45] |
| DMAPP | AMP | 7.9 ± 0.6 | 5.3 × 10−2 | [45] | |
| HMBPP | AMP | 8.2 ± 0.4 | 2.5 × 10−2 | [45] | |
| FasD | DMAPP | AMP | 0.030 ± 0.016 | 7.9 × 10−3 | [22] |
| HMBPP | AMP | 0.026 ± 0.003 | 6.8 × 10−4 | [22] | |
| HMBPP | ADP | 0.550 ± 0.020 | 9.4 × 10−4 | [22] | |
| NoIPT1 | AMP | DMAPP | 0.63 ± 0.180 | n.a. | [28] |
| DMAPP | AMP | 27.1 ± 2.47 | 5.3 × 10−3 * | [28] | |
| HMBPP | AMP | n.d. | n.d. | [28] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Frébortová, J.; Frébort, I. Biochemical and Structural Aspects of Cytokinin Biosynthesis and Degradation in Bacteria. Microorganisms 2021, 9, 1314. https://doi.org/10.3390/microorganisms9061314
Frébortová J, Frébort I. Biochemical and Structural Aspects of Cytokinin Biosynthesis and Degradation in Bacteria. Microorganisms. 2021; 9(6):1314. https://doi.org/10.3390/microorganisms9061314
Chicago/Turabian StyleFrébortová, Jitka, and Ivo Frébort. 2021. "Biochemical and Structural Aspects of Cytokinin Biosynthesis and Degradation in Bacteria" Microorganisms 9, no. 6: 1314. https://doi.org/10.3390/microorganisms9061314