
CosR Regulation of perR Transcription for the Control of Oxidative Stress Defense in Campylobacter jejuni



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
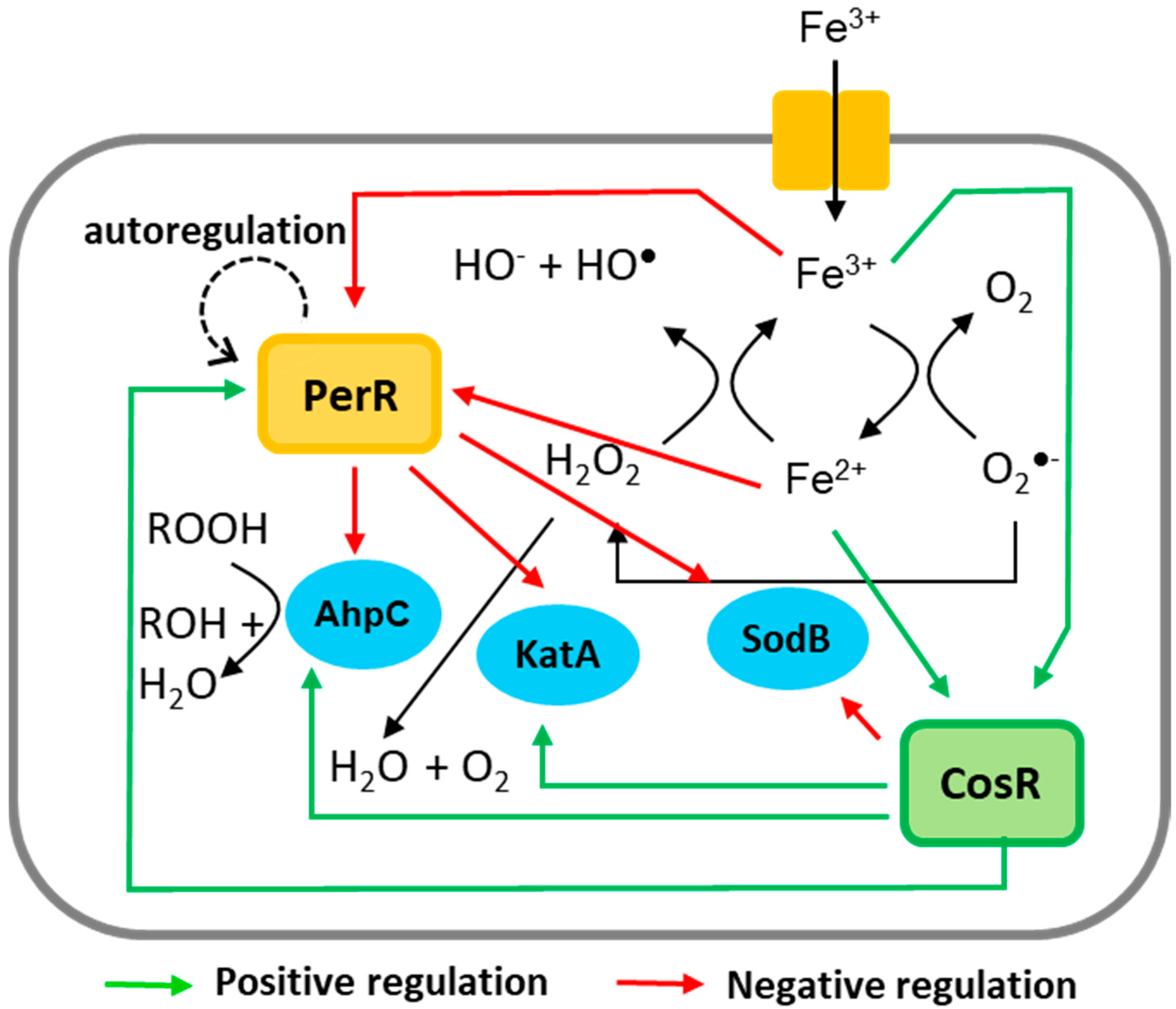
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
2.2. CosR Knockdown and Overexpression
2.3. Construction of a PcosR::lacZ Promoter Fusion and β-Galactosidase Assay
2.4. Electrophoretic Mobility Shift Assay
2.5. DNase I Footprinting Assay
3. Results
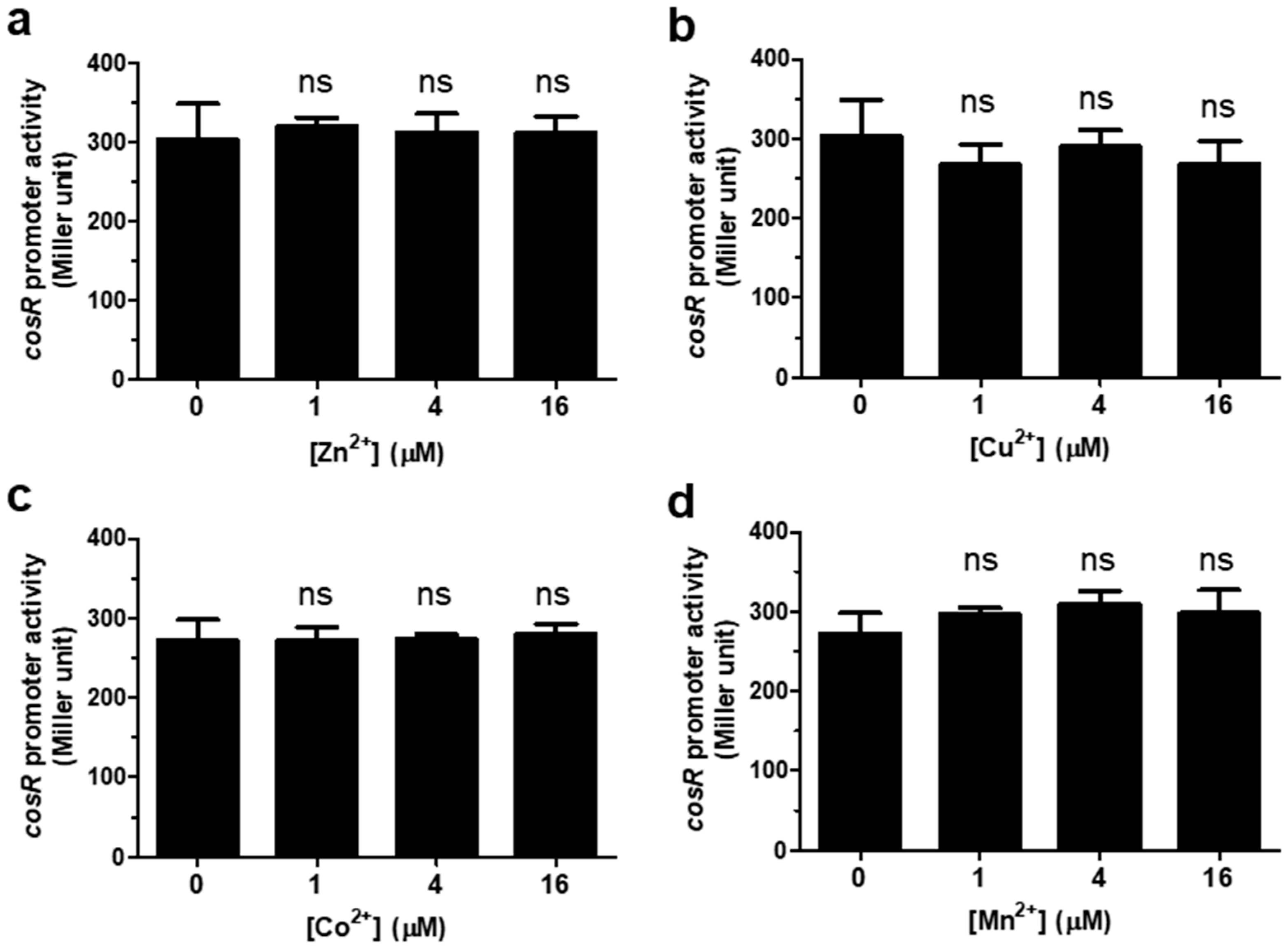
3.1. Regulation of cosR Transcription by Iron
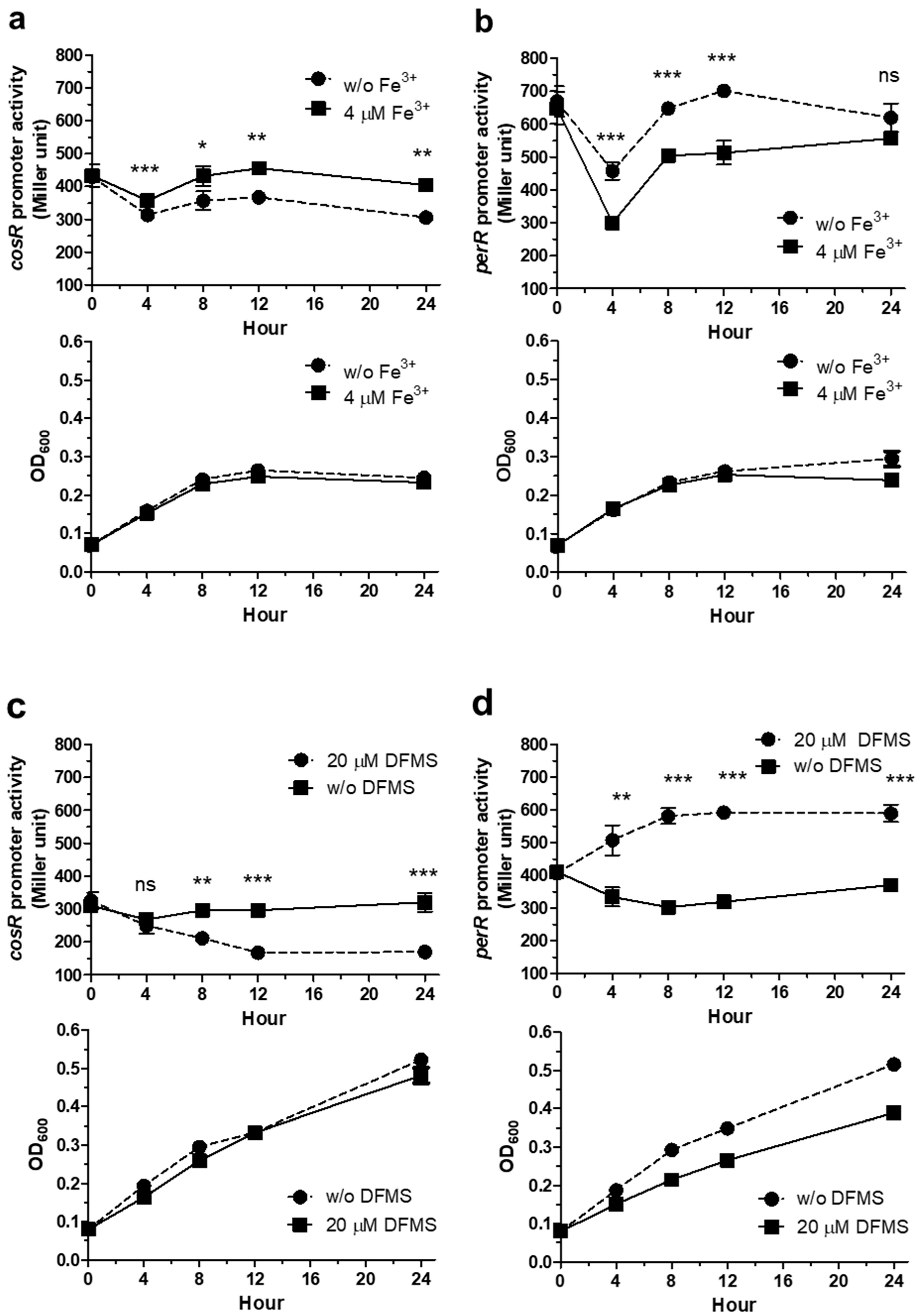
3.2. Iron Regulation of cosR and perR Transcription over the Growth of C. jejuni
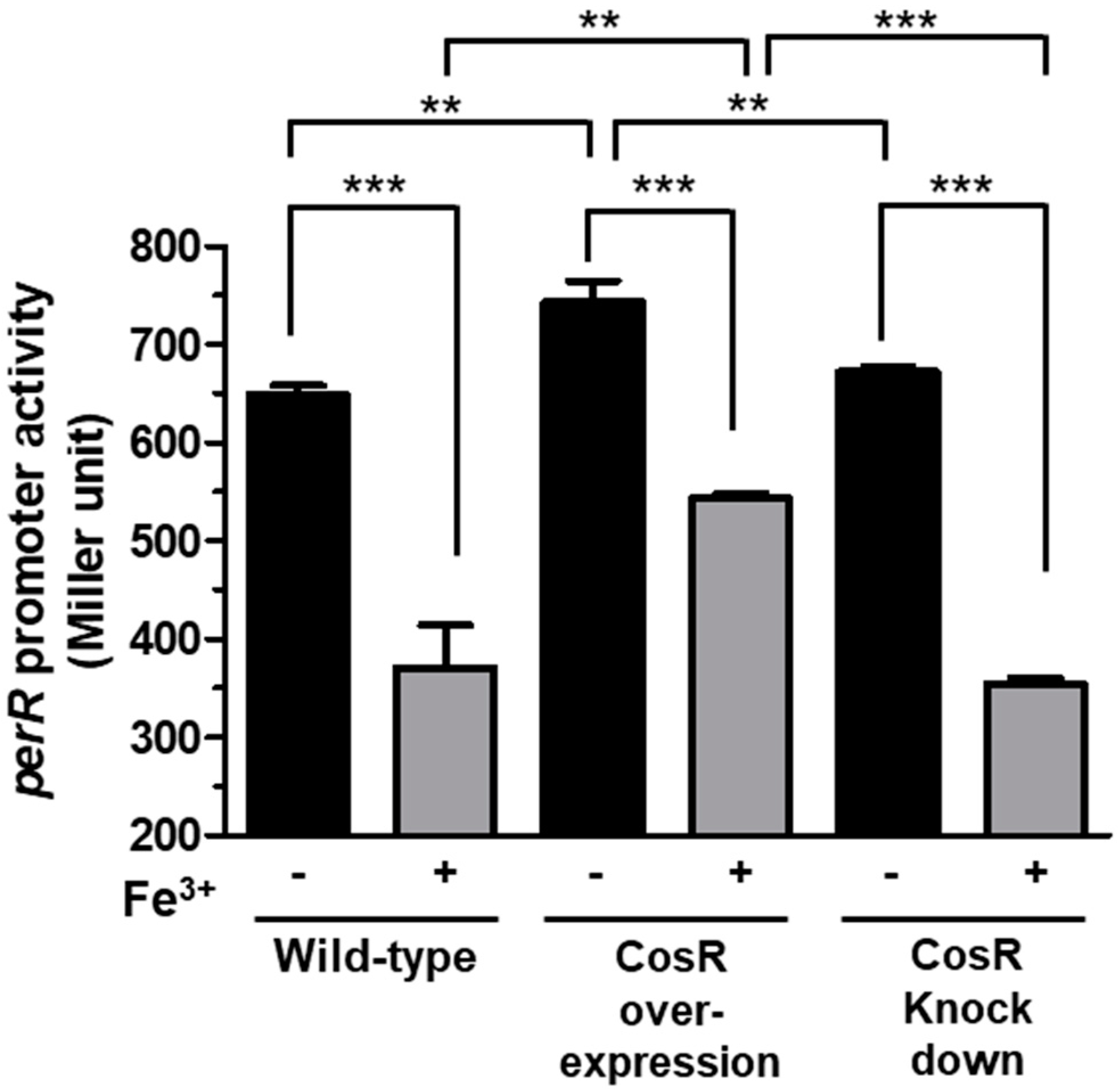
3.3. CosR Regulation of perR Transcription
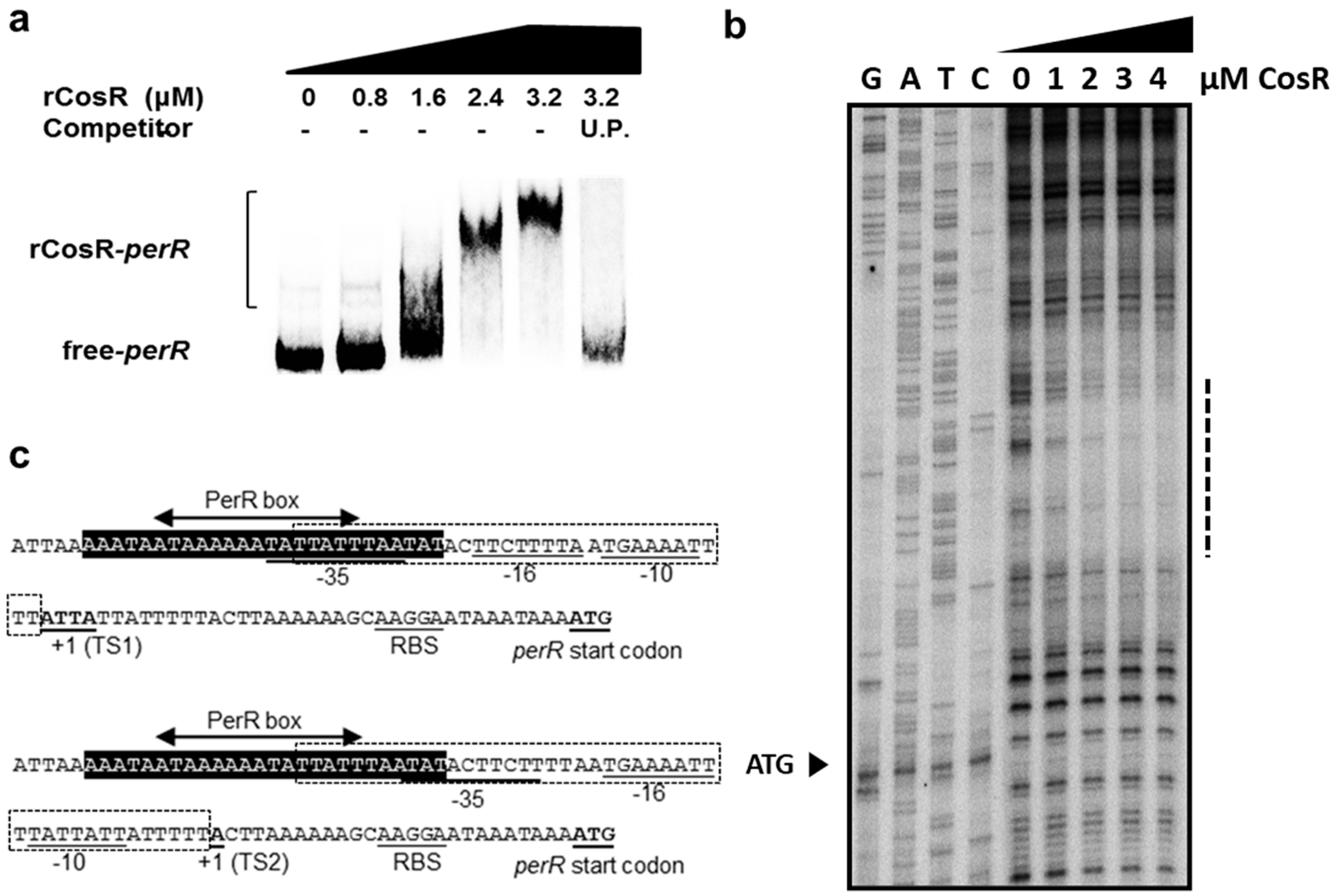
3.4. CosR Binding to the perR Promoter
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Kirk, M.D.; Pires, S.M.; Black, R.E.; Caipo, M.; Crump, J.A.; Devleesschauwer, B.; Dopfer, D.; Fazil, A.; Fischer-Walker, C.L.; Hald, T.; et al. World health organization estimates of the global and regional disease burden of 22 foodborne bacterial, protozoal, and viral diseases, 2010: A data synthesis. PLoS Med. 2015, 12, e1001921. [Google Scholar]
- Heimesaat, M.M.; Backert, S.; Alter, T.; Bereswill, S. Human campylobacteriosis-a serious infectious threat in a one health perspective. Curr. Top. Microbiol. Immunol. 2021, 431, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Wachira, V.K.; Peixoto, H.M.; de Oliveira, M.R.F. Systematic review of factors associated with the development of Guillain-Barre syndrome 2007–2017: What has changed? Trop. Med. Int. Health TM IH 2019, 24, 132–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skarp, C.P.A.; Hänninen, M.L.; Rautelin, H.I.K. Campylobacteriosis: The role of poultry meat. Clin. Microbiol. Infect. 2016, 22, 103–109. [Google Scholar] [CrossRef] [Green Version]
- Oh, E.; Chui, L.; Bae, J.; Li, V.; Ma, A.; Mutschall, S.K.; Taboada, E.N.; McMullen, L.M.; Jeon, B. Frequent implication of multistress-tolerant Campylobacter jejuni in human infections. Emerg. Infect. Dis. 2018, 24, 1037–1044. [Google Scholar] [CrossRef] [PubMed]
- Oh, E.; McMullen, L.; Jeon, B. Impact of oxidative stress defense on bacterial survival and morphological change in Campylobacter jejuni under aerobic conditions. Front. Microbiol. 2015, 6, 295. [Google Scholar] [CrossRef] [PubMed]
- Oh, E.; McMullen, L.M.; Chui, L.; Jeon, B. Differential survival of hyper-aerotolerant Campylobacter jejuni under different gas conditions. Front. Microbiol. 2017, 8, 954. [Google Scholar] [CrossRef] [Green Version]
- Handley, R.A.; Mulholland, F.; Reuter, M.; Ramachandran, V.K.; Musk, H.; Clissold, L.; Le Brun, N.E.; van Vliet, A.H.M. PerR controls oxidative stress defence and aerotolerance but not motility-associated phenotypes of Campylobacter jejuni. Microbiology 2015, 161, 1524–1536. [Google Scholar] [CrossRef] [Green Version]
- Chiang, S.M.; Schellhorn, H.E. Regulators of oxidative stress response genes in Escherichia coli and their functional conservation in bacteria. Arch. Biochem. Biophys. 2012, 525, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Imlay, J.A. Cellular defenses against superoxide and hydrogen peroxide. Annu. Rev. Biochem. 2008, 77, 755–776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Vliet, A.H.; Baillon, M.L.; Penn, C.W.; Ketley, J.M. Campylobacter jejuni contains two fur homologs: Characterization of iron-responsive regulation of peroxide stress defense genes by the PerR repressor. J. Bacteriol. 1999, 181, 6371–6376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.C.; Oh, E.; Hwang, S.; Ryu, S.; Jeon, B. Non-selective regulation of peroxide and superoxide resistance genes by PerR in Campylobacter jejuni. Front. Microbiol. 2015, 6, 126. [Google Scholar] [CrossRef] [Green Version]
- Hwang, S.; Kim, M.; Ryu, S.; Jeon, B. Regulation of oxidative stress response by CosR, an essential response regulator in Campylobacter jejuni. PLoS ONE 2011, 6, e22300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, S.; Miller, W.G.; Ryu, S.; Jeon, B. Divergent distribution of the sensor kinase CosS in non-thermotolerant campylobacter species and its functional incompatibility with the response regulator CosR of Campylobacter jejuni. PLoS ONE 2014, 9, e89774. [Google Scholar] [CrossRef] [Green Version]
- Garénaux, A.; Guillou, S.; Ermel, G.; Wren, B.; Federighi, M.; Ritz, M. Role of the Cj1371 periplasmic protein and the Cj0355c two-component regulator in the Campylobacter jejuni NCTC 11168 response to oxidative stress caused by paraquat. Res. Microbiol. 2008, 159, 718–726. [Google Scholar] [CrossRef] [PubMed]
- Raphael, B.H.; Pereira, S.; Flom, G.A.; Zhang, Q.; Ketley, J.M.; Konkel, M.E. The Campylobacter jejuni response regulator, CbrR, modulates sodium deoxycholate resistance and chicken colonization. J. Bacteriol. 2005, 187, 3662–3670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, S.; Zhang, Q.; Ryu, S.; Jeon, B. Transcriptional regulation of the CmeABC multidrug efflux pump and the KatA catalase by CosR in Campylobacter jejuni. J. Bacteriol. 2012, 194, 6883–6891. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.; Hwang, S.; Ryu, S.; Jeon, B. Regulation of perR expression by iron and PerR in Campylobacter jejuni. J. Bacteriol. 2011, 193, 6171–6178. [Google Scholar] [CrossRef] [Green Version]
- Butcher, J.; Handley, R.A.; van Vliet, A.H.; Stintzi, A. Refined analysis of the Campylobacter jejuni iron-dependent/independent Fur- and PerR-transcriptomes. BMC Genom. 2015, 16, 498. [Google Scholar] [CrossRef] [Green Version]
- Oh, E.; Andrews, K.J.; Jeon, B. Enhanced biofilm formation by ferrous and ferric iron through oxidative stress in Campylobacter jejuni. Front. Microbiol. 2018, 9, 1204. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Zeng, X.; Lin, J. Enterobactin-specific antibodies inhibit in vitro growth of different gram-negative bacterial pathogens. Vaccine 2020, 38, 7764–7773. [Google Scholar] [CrossRef]
- Palyada, K.; Threadgill, D.; Stintzi, A. Iron acquisition and regulation in Campylobacter jejuni. J. Bacteriol. 2004, 186, 4714–4729. [Google Scholar] [CrossRef] [Green Version]
- van Vliet, A.H.; Wooldridge, K.G.; Ketley, J.M. Iron-responsive gene regulation in a Campylobacter jejuni fur mutant. J. Bacteriol. 1998, 180, 5291–5298. [Google Scholar] [CrossRef] [Green Version]
- Holmes, K.; Mulholland, F.; Pearson, B.M.; Pin, C.; McNicholl-Kennedy, J.; Ketley, J.M.; Wells, J.M. Campylobacter jejuni gene expression in response to iron limitation and the role of Fur. Microbiology 2005, 151, 243–257. [Google Scholar] [CrossRef]
- Wösten, M.M.; Boeve, M.; Koot, M.G.; van Nuenen, A.C.; van der Zeijst, B.A. Identification of Campylobacter jejuni promoter sequences. J. Bacteriol. 1998, 180, 594–599. [Google Scholar] [CrossRef] [Green Version]
- Schaefer, J.; Jovanovic, G.; Kotta-Loizou, I.; Buck, M. Single-step method for β-galactosidase assays in Escherichia coli using a 96-well microplate reader. Anal. Biochem. 2016, 503, 56–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barnawi, H.; Masri, N.; Hussain, N.; Al-Lawati, B.; Mayasari, E.; Gulbicka, A.; Jervis, A.J.; Huang, M.H.; Cavet, J.S.; Linton, D. RNA-based thermoregulation of a Campylobacter jejuni zinc resistance determinant. PLoS Pathog. 2020, 16, e1009008. [Google Scholar] [CrossRef]
- Bradley, J.M.; Svistunenko, D.A.; Wilson, M.T.; Hemmings, A.M.; Moore, G.R.; Le Brun, N.E. Bacterial iron detoxification at the molecular level. J. Biol. Chem. 2020, 295, 17602–17623. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.W.; Helmann, J.D. Functional specialization within the Fur family of metalloregulators. Biometals 2007, 20, 485–499. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.W.; Helmann, J.D. The PerR transcription factor senses H2O2 by metal-catalysed histidine oxidation. Nature 2006, 440, 363–367. [Google Scholar] [CrossRef] [PubMed]
- Jacquamet, L.; Traore, D.A.; Ferrer, J.L.; Proux, O.; Testemale, D.; Hazemann, J.L.; Nazarenko, E.; El Ghazouani, A.; Caux-Thang, C.; Duarte, V.; et al. Structural characterization of the active form of PerR: Insights into the metal-induced activation of PerR and Fur proteins for DNA binding. Mol. Microbiol. 2009, 73, 20–31. [Google Scholar] [CrossRef]
- Lund, E.K.; Wharf, S.G.; Fairweather-Tait, S.J.; Johnson, I.T. Increases in the concentrations of available iron in response to dietary iron supplementation are associated with changes in crypt cell proliferation in rat large intestine. J. Nutr. 1998, 128, 175–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naikare, H.; Palyada, K.; Panciera, R.; Marlow, D.; Stintzi, A. Major role for FeoB in Campylobacter jejuni ferrous iron acquisition, gut colonization, and intracellular survival. Infect. Immun. 2006, 74, 5433–5444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, C.E.; Rock, J.D.; Ridley, K.A.; Williams, P.H.; Ketley, J.M. Utilization of lactoferrin-bound and transferrin-bound iron by Campylobacter jejuni. J. Bacteriol. 2008, 190, 1900–1911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, F.; Zeng, X.; Haigh, R.D.; Ketley, J.M.; Lin, J. Identification and characterization of a new ferric enterobactin receptor, CfrB, in Campylobacter. J. Bacteriol. 2010, 192, 4425–4435. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, M.; Hwang, S.; Ryu, S.; Jeon, B. CosR Regulation of perR Transcription for the Control of Oxidative Stress Defense in Campylobacter jejuni. Microorganisms 2021, 9, 1281. https://doi.org/10.3390/microorganisms9061281
Park M, Hwang S, Ryu S, Jeon B. CosR Regulation of perR Transcription for the Control of Oxidative Stress Defense in Campylobacter jejuni. Microorganisms. 2021; 9(6):1281. https://doi.org/10.3390/microorganisms9061281
Chicago/Turabian StylePark, Myungseo, Sunyoung Hwang, Sangryeol Ryu, and Byeonghwa Jeon. 2021. "CosR Regulation of perR Transcription for the Control of Oxidative Stress Defense in Campylobacter jejuni" Microorganisms 9, no. 6: 1281. https://doi.org/10.3390/microorganisms9061281